










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkVaccimonitor
versión On-line ISSN 1025-0298
Vaccimonitor v.18 n.2 Ciudad de la Habana Mayo-ago. 2009
ARTICULOS ORIGINALES
Inmunización intranasal con AFCo1 induce respuesta inmune de memoria, sistemica y mucosal en ratones neonatal.
Intranasal immunization with AFCo1 induce systemic, mucosal and memory immune response in neonatal mice
Julio A. Balboa, Belkis Romeu, Maribel Cuello, Caridad Zayas, Judith del Campo, Elizabeth González, Reynaldo Acevedo, Miriam Lastre, Osmir Cabrera, and Oliver Pérez
Immunology Department, Research Vice-presidency, Finlay Institute, P.O. Box 16017, Havana, Cuba
email: jbalboa@finlay.edu.cu
Abstract
Neonates have a poorly developed immune system. Respiratory pathogens cause disease during early periods of live. Consequently, it is important to develop protective vaccines that induce immunity and immunological memory against respiratory pathogens early in life. Intranasal (i.n.) route could be an effective via for immunization. Therefore, we explored the effectiveness of AF (Adjuvant Finlay) PL1 (Proteoliposome) from Neisseria meningitidis serogroup B and its derivate Cochleate (AFCo1) by nasal route in neonatal mice. They were immunized i.n. 3 times 7 days apart and anti PL systemic and mucosal antibody response were measured by ELISA. In addition, a prime-boost strategy was used to evaluate the humoral immune response in neonate mice. The 3 doses of AFPL1 or AFCo1 induced significant levels of anti PL IgG antibodies in comparison whit control, but AFCo1 (2017 U/mL) was significantly higher than AFPL1 (1107 U/mL). AFCo1 and AFPL1 induced a predominant Th1 pattern with IgG2a/IgG1 >1 by i.n. immunization and AFCo1 induced a high anti PL IgA saliva response in saliva. Interestingly, one nasally prime at 7 days of born and a memory one boost i.n. dose 9 weeks later with AFCo1 or AFPL1 showed similar specific IgG levels and IgG2a/IgG1 relation than 3 i.n. doses in adult mice. In conclusion, these results represent the first report of neonatal intranasal vaccination using AFCo1 capable to induce systemic and mucosal immunity and priming for memory.
Keywords: Neisseria meningitidis,Outer membrane vesicle, Proteoliposome, AFCo1, AFPL1, Neonatal immunization.
Introduction
It is becoming increasing clear that both human and mice neonates mount various responses, ranging from deficient or deviant to fully mature, depending on the conditions of antigen exposure and it was demonstrated that the strong Th1-cell promoting adjuvant, the complete Freund's adjuvant elicited similar response than adult mice (1).
The impairment of CD4 and CD8 T-cell function in early life results can be overcome by specific adjuvants/delivery systems. It is also important to circumvent maternal antibodies inhibition of vaccine response. An alternative strategy includes the development of mucosal vaccine.
Neisseria meningitidis outer membrane vesicles (OMV, Proteoliposome, PL) has been widely used as a core antigen for meningococcal vaccination in the Cuban using Meningococcal BC vaccine VA-MENGOC-BC® (2). Intraperitoneal (i.p.) immunization of neonatal mice with N. meningitidis PL obtained from the epidemic meningococcal strain Cu385 (B:4:P1.19,15) induced a significant antibody response, conferred protection against bacteremia to mice challenged with meningococci and elicited a significant increase in antibody levels to a subsequent antigen dose, compared to unprimed mice. However, intranasal (i.n.) immunization was less effective and did not generate a protective immune response (3).
The structural transformation of the PL into cochleate structures (AFCo1) provides additional properties to PL as higher stability and better induction of mucosal immune response (4).
The aim of this work was to demonstrate the capacity of AFCo1 to induce systemic and mucosal response in neonatal mice. In parallel, the priming for memory was also evaluated.
Materials and methods
PL of the epidemic meningococcal strain Cu385 (B:4:P1.19,15) was supplied as ethanol precipitate by the manufacturing plant of Finlay Institute, Cuba. It was dissolved in Tris Cl buffer 30 mM and sodium deoxicolate 0.5 mM at 1 mg/mL. AFCo1 was prepared from this PL by rotational dialysis (4). Pregnant female BALB/c (H-2d) mice and newborn mice (1 to 3 days) were obtained from The National Center for the Production of Laboratory Animals, Havana, Cuba and housed in the Animal Care Unit during the experiments. Newborn mice were caged with their mothers during lactation, which lasted for 3 weeks.
Neonatal (7 days old) mice were immunized with AFCo1 or PL (50 µg per dose per mouse). Mice received three inoculations for mucosal routes, separated by a 7 day interval. Age-matched mice inoculated with sterile Tris-Cl buffer were used as controls. Antibodies specific titers (IgG, IgA and IgG subclasses) in sera and saliva of immunized mice were determined by capture enzyme-linked immunosorbent assay (ELISA) (5). The differences between the groups were evaluated by a Tukey multiple comparison test and by a Student's t test using Graph Pad Prism 4 software (CA, USA).
Results and discussion
Previous studies from our group suggest that AFPL1 has a strong immunomodulatory activity over the immune cells involved in the initiation of immune responses and has the potential to deliver antigens to the immune system. Also, AFCo1 are immunogenic when administered via intramuscular or nasal routes and can activate mucosal immunity only through the latter (6) Bearing in mind these previous results, we used the immunomodulatory activity and the delivery system capacity of AFPL1, a nanoparticle, and its derivative microparticle cochleate structure AFCo1, which is more stable and could improve antigen presentation to the immune system.
It was previously demonstrated that i.n. immunization of adult mice with AFCo1 and AFPL1 induced strong specific IgG response in sera against PL antigens, significantly higher in AFCo1 immunized groups. AFCo1 enhanced the IgG response and the IgG2a subclass (7).
Intranasal immunization of neonatal mice with AFCo1 induced significantly (p<0.05) higher anti-PL specific serum IgG response than those observed in the group received i.n AFPL1 (Figure 1). In addition, both groups induced saliva anti-PL IgA response, but AFCo1 induced nine times more anti-PL IgA response than the AFPL1 group. J. Campo et al.(8) reported that AFCo1 was considerably more potent than AFPL1 for inducing IgA in saliva and vaginal washes, which it is in correspondence with the results that we obtained in neonatal mice.
The relation IgG2a/IgG1 was >1 for AFCo1 and AFPL1 i.n. immunized groups of neonatal mice similar than those observed in adults mice, both formulations induced a predominant Th1 pattern.
To assess the safety of AFCo1 and AFPL1 in neonatal mice, weight gain and the number of deaths were monitored weekly and compared to age-matched mice given sterile Tris Cl buffer. No deaths occurred, and no effect was observed on weight gain in infant and neonatal mice immunized i.n. with AFCo1 or AFPL1 compared to unimmunized mice (data not shown), indicating that AFCo1 and PL are safe and have a low reactogenicity profile already in early life.
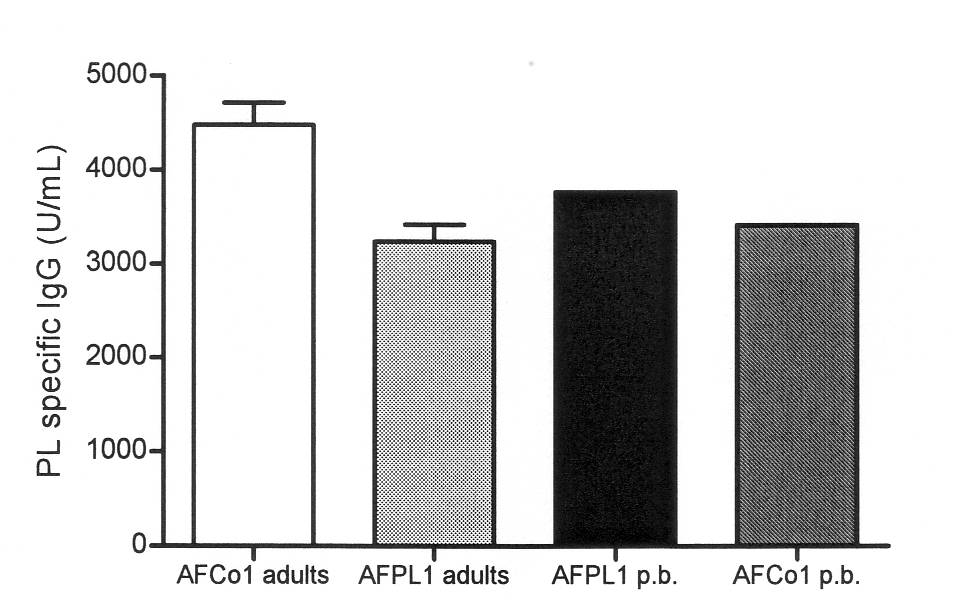
To explore the potential for priming mucosal immunization in early murine life, neonatal mice (7 days old) were immunized once with AFCo1 or AFPL1. Nine weeks after priming, mice were boosted with the similar corresponding dose and route than prime dose. The serum IgG response was comparable to that induced by AFCo1 and AFPL1 in adults groups. The IgG response in neonatal mice was compared with the media specific IgG (Figure 2) and the specific IgG2a/IgG1 ratio obtained in adult mice. Neonatal immunization does not generally lead to early and strong antibody response, this could be from suboptimal antigen presenting cell -T cell interaction, and can be overcome by use of specific adjuvants and delivery systems. Neonatal triggered Th2 responses are more difficult to fully redirect toward Th1 responses than responses induced in adults (9). However, the use of strong Th1-driving adjuvants or formulations could redirect fully the neonatal Th2 responses. The responses pattern induced in neonate mice after a prime-boost strategy with AFCo1 and AFPL1 corresponded with a full redirect Th2 pattern into Th1 response pattern. Thus, the development of optimal neonatal priming-early boosting strategies appears as promising, enhance subsequent responses and/or contribute to protection.
Figure 1. AFCo1 enhance PL-specific antibody response in neonatal mice. Anti-PL IgG antibodies in serum 21 days after the i.n. immunization with tree doses (50 µg per dose) of AFCo1 or PL at 7, 14, and 21 days after birth. Denotes significance statistical difference (p<0.05) between the antibody levels in the indicated groups to Student's t test using Graph Pad Prism 4 software.
Figure 2. Anti-PL IgG antibodies responses in serum after i.n. immunization with tree doses (50 µg per dose) of AFCo1 or AFPL1 at 0, 7, and 14 days in adult mice. Neonatal mice was prime (p) i.n. at 7 days of born and 9 weeks later they were i.n. boost (b) with the homologous compaund.
References
1. Adkins B, Leclerc C, Marshall-Clarke S. Neonatal adaptive immunity comes of age. Nature Reviews/Immunology 2004;4:553-64.
2. Sierra GVG, Campa HC, Varcacel NM, García IL, Izquierdo PL, Sotolongo PF, et. al. Vaccine against group B Neisseria meningitidis: Protection trial and mass vaccination results in Cuba. NIPH Annals 1991;14:195-210.
3. González S, Caballero E, Soria Y, Cobas K, Granadillo M, Pajón R. Immunization with Neisseria meningitidis outer membrane vesicles prevents bacteremia in neonatal mice. Vaccine 2006;24:1633-43.
4. Pérez O, Bracho G, Lastre M, Mora N, Del campo, J, Gil D, et al. Novel adjuvant base on a proteoliposome-derived cochleate structure containing native lipopolysaccharide as a pathogen-associated molecular pattern Immunol. Cell Biol 2004;82:603-10.
5. Pérez O, Lastre M, Cabrera O, del Campo J, Bracho G, Cuello M, et. al. New Vaccines Require Potent Adjuvants like AFPL1 and AFCo1. Scandinavian Journal of Immunology 2007;66:271-7.
6. Rodríguez T, Pérez O, Menager N, Ugrinovic S, Bracho G., Mastroeni P. Interactions of proteoliposomes from serogroup B Neisseria meningitidis with bone marrow-derived dendritic cells and macrophages: adjuvant effects and antigen delivery. Vaccine 2005;26:1312-21.
7. Bracho G, Lastre M, Del Campo J, Zayas C, González D, Gil D et. al. Proteoliposome derived cochleate as novel adjuvant. Vaccine. 2006;24(S2):30-1.
8. del Campo J, Zayas C; Romeo B; Acevedo R; González E; Bracho G; González L; Cuello M; Cabrera O; Balboa J; Lastre M. Mucosal immunization using proteoliposome and cochleate structures from Neisseria meningitis serogroup B induce mucosal and systemic responses. METHODS-D-09-00010R1. 2009 Accepted. 9. Siegrist CA. Neonatal and early life vaccinology. Vaccine 2001;19:3331-46.


{kind=link}
{kind=link}