










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkVaccimonitor
versión On-line ISSN 1025-0298
Vaccimonitor v.18 n.2 Ciudad de la Habana Mayo-ago. 2009
ARTICULOS ORIGINALES
Dinámicas de las interacciones de Neisseria meningitidis con las barreras celulares y los efectores inmunes.
Dynamics of Neisseria meningitidis interactions with human cellular barriers and immune effectors.
Natalie J. Griffiths, Claudia Sa E Cunha, Isabel Murillo, Abdel-Rahman Youssef, Elena Borodina, Darryl J. Hill and Mumtaz Virji
Department of Cellular and Molecular Medicine, School of Medical Sciences,
University of Bristol, Bristol BS8 1TD, UK
Abstract
Neisseria meningitidis outer membrane (OM) adhesins, Opa and Opc are known to exert significant influence on bacterial adhesion and invasion properties. They are also likely to affect the dynamics of cellular barrier penetration as they target human receptors that are subject to upregulation under inflammatory conditions. As some of the targeted receptors are also expressed on immune cells, it is possible that the OM proteins, when presented on bacteria or in OM vesicle vaccines, have the additional capacity to modulate host immune responses. In our recent studies, in vitro model systems were used to further explore these possibilities. The studies illustrated that the major human receptors targeted by Opa and Opc, i.e. CEACAMs and integrins, when upregulated by inflammatory cytokines, encourage enhanced cellular adhesion, invasion and barrier traversal. Tissue infiltration by fully capsulate bacteria via Opa proteins was also observed for piliated Opa+ meningococci. Other studies indicate that Opc increases meningococcal resistance to serum-mediated killing by binding to the complement regulatory molecule vitronectin. In addition, although adverse immunomodulatory effects have been reported for Opa-expressing gonococci and meningococcal OMVs, our studies indicate that interactions with CD4+ T cell expressed CEACAM1 does not offer immunomodulatory properties to meningococci.
Keywords: Neisseria meningitidis, Opa, Opc
Introduction
It is now well established that the major receptors targeted by neisserial Opa proteins belong to the CEACAM (carcinoembryonic antigen-related cell adhesion molecule) family of receptors, of which CEACAM1 is the most widely targeted by distinct Opa types. CEACAM1 is one of several related molecules expressed on human epithelial cells, endothelial cells and leukocytes but it is the only member of the family expressed on T cells.
One of our interests has been to assess how neisserial Opa targeting of CEACAM1, expressed on human barrier cells as well as immune cells, might alter host susceptibility to infections by the pathogens. In recent studies, we have examined the potential role of CEACAM1 in increasing target cell susceptibility to meningococcal infiltration. In vitro transfected cellular model systems and human respiratory epithelial cells exposed to IFNg demonstrated the importance of the receptor upregulation in increased cellular invasion by meningococci. The data imply that cell-surface remodelling by virally-induced cytokines could be one factor that increases host susceptibility to bacterial infection (1).
Further, it is noteworthy that polymorphisms in CEACAM structure have been associated with changes in meningococcal interactions and virulence as demonstrated by in vitro mutagenesis studies of CEACAM1 (2) and in surveys of meningococcal case versus control samples (3). In addition to the association of certain CEACAM haplotypes with increased host susceptibility, certain meningococcal Opa repertoires were correlated with hyper-invasiveness and disease (3, 4). Association of limited numbers of Opa repertoires with disease also suggests that Opa proteins could be candidate vaccine antigens in their own right. Therefore, understanding their full potential as virulence factors is important for future approaches to control meningococcal infection.
In addition, Opc, another opacity protein, unique to N. meningitidis is an effective invasin for human endothelial cells (5). Meningococcal entry into the meninges may also involve entry via brain endothelial cells (6). Our recent studies have assessed how integrin targeting enables meningococci to interact with endothelial cells at diverse vascular sites at which integrin profiles may vary considerably (7). Opc has also been shown to interact with intracellular alpha-actinin, a modulator of several receptor and cytoskeletal functions, whether this enables meningococci to influence cellular functions to facilitate their passage across human barrier cells remains to be shown (8). In addition, we have analysed how Opc targeting of serum factors such as vitronectin may enhance its serum resistance properties. Below we provide some data from studies on potential immunomodulatory properties of meningococcal opacity proteins.
A. Opc interactions with activated vitronectin lead to increased serum resistance of meningococci
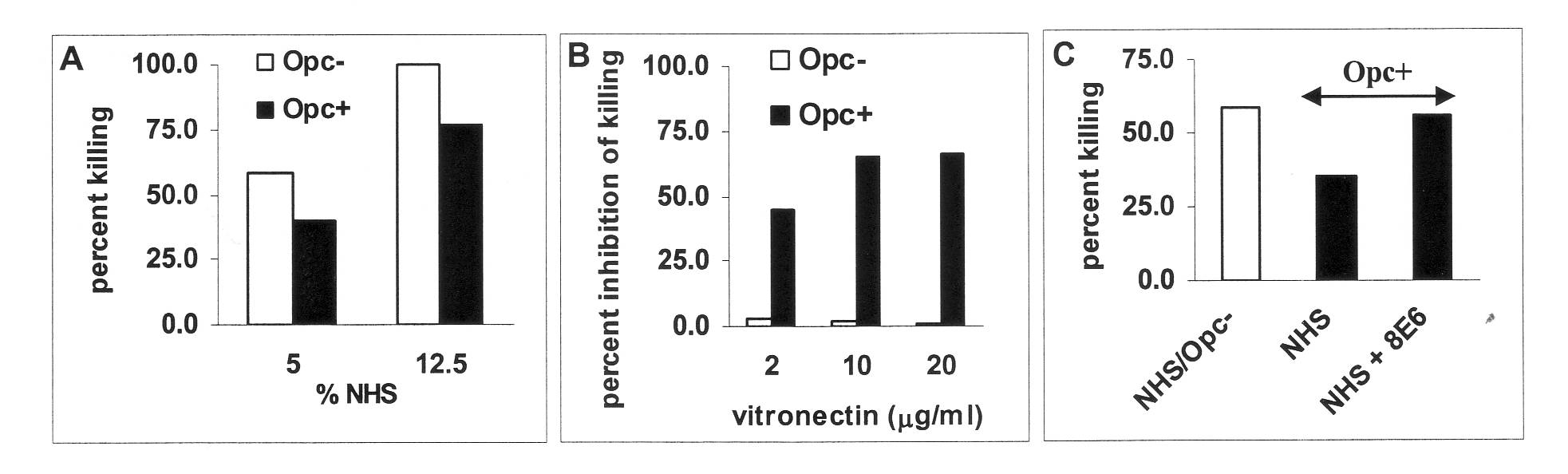
The major serum protein targeted by Opc is vitronectin, a well known modulator of complement action. We have analysed the molecular structural requirement for Opc targeting of human vitronectin and the resultant effect on serum resistance. Although Opc may bind to several serum proteins, vitronectin was found to be the major target for Opc; and it is the activated/unfolded form of vitronectin that is particularly targeted. We have identified the binding region for Opc on vitronectin, which becomes exposed only in the activated form of the protein. In addition, binding to vitronectin which is observed for Opc+ but not Opc- bacteria, leads to increased serum resistance (Fig.1). Depletion of activated serum vitronectin by anti-vitronectin antibodies decreased such serum resistance (Figure. 1C).
Figure 1. Serum killing of acapsulate Opc+ and Opc- N. meningitidis isolates (NHS: normal human serum; endogenous complement). Opc-expressing meningococci are inherently more serum resistant (A). The addition of activated vitronectin to NHS inhibits killing and saturable inhibition of killing in the presence of 10% NHS with additional activated vitronectin is observed only with Opc+ bacteria (B). The monoclonal anti-vitronectin antibody (8E6) depletes activated vitronectin available for Opc interactions, and restores serum killing of Opc+ N. meningitidis (C). Similar observations were made using capsulate bacteria (not shown).
Conclusions
Opc is a multifunctional protein. Its binding to vitronectin has been shown to enhance cellular interactions. Current studies have shown that vitronectin targeting also leads to increased serum resistance: the addition of activated vitronectin increases Opc-expressing meningococcal survival in normal adult serum whereas its depletion results in restoration of serum bactericidal activity.
B. Direct adhesion to human CD4+ T cells by meningococcal opacity proteins does not affect T cell responses to other neisserial antigens.
Several neisserial antigens have been shown to induce proliferative response in T cells (e.g. reference (9)). In contrast, interactions of Opa+ N. gonorrhoeae and N. meningitidis OMV with CEACAM1 inhibited CD4+ T cell responses (10, 11). Whether Opa-expressing live meningococci exert similar effects is not known. In view of this and of the notion that N. meningitidis carriage itself may be an immunising event that may induce lasting T cell memory (12), further studies were conducted to define the consequences of meningocococcal interactions with human CD4+ T cells.
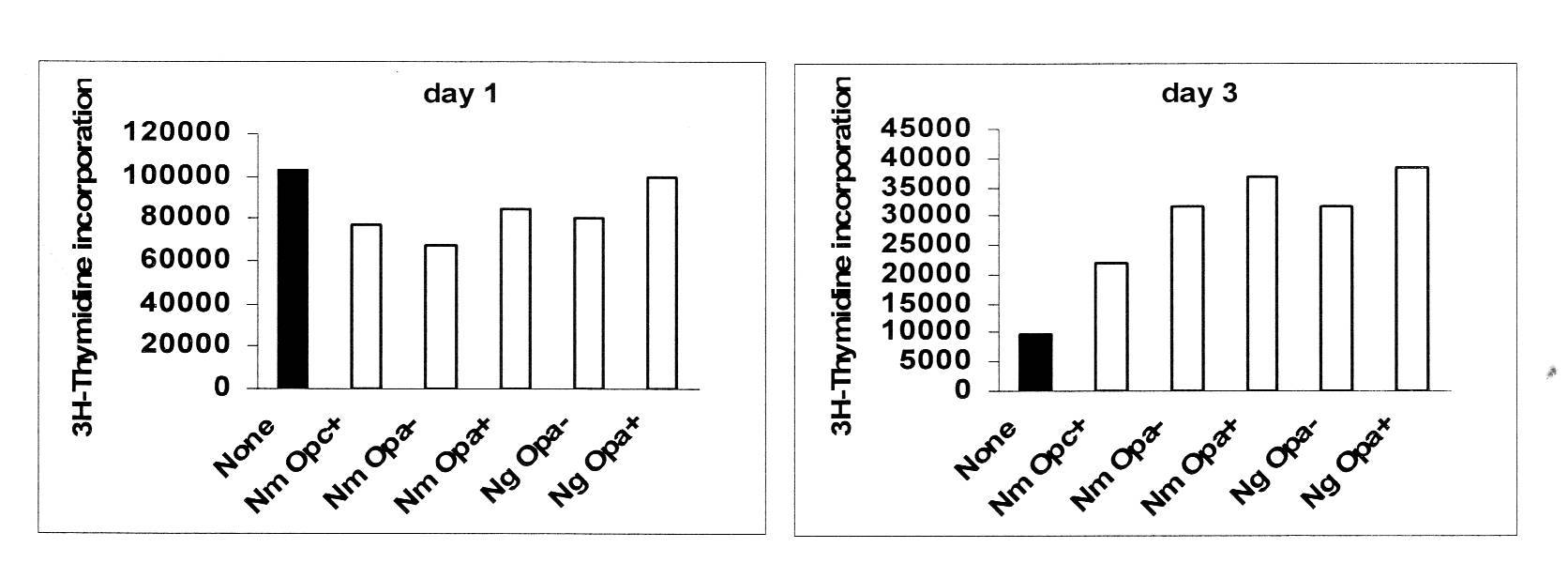
For this purpose, live and killed N. meningitidis of distinct serogroups either expressing or lacking Opa and Opc proteins, were added to isolated CD4+ T cells prestimulated to induce CEACAM1 expression. An early Opa-independent transient decrease in T cell proliferation was observed under some conditions which was followed by a strong stimulatory effect on T cell immune functions as assessed by 3H-thymidine incorporation assays (Fig. 2) and cytokine responses (not shown). Importantly, despite increased adhesion mediated by the opacity proteins, no Opa-dependent inhibitory effect on T cell proliferation was seen using a variety of assay conditions.
Figure 2. Effect of neisserial infection on proliferation of activated CD4+ T cells. CD4+ T cells were stimulated with anti-CD3 and anti-CD28 for 48 h, washed and incubated with IL-2 for further 48 h. After washing, the cells were left uninfected (filled bars) or infected with neisserial strains in the presence of soluble anti-CD3. Three hours after infection, gentamicin (50mg/ml) was added. At daily intervals after infection, cells were pulsed with 3H-thymidine. Average cpm incorporated in duplicate samples at day 1 and day 3 are shown (SE +/- 15%). Corroborating data were also obtained using the CFSE dilution method. Note: Opc+ and Opa+ meningococci (Nm) and Opa+ gonococci (Ng) adhere in greater numbers to stimulated CD4+ T cells than bacteria not expressing the proteins (not shown).
Conclusions
Overall, these data suggest that certain surface components of the pathogenic Neisseria can exert early mild inhibitory effects but these are rapidly followed by longer lasting, stronger stimulatory effects on CD4+ T cells. Most importantly, the opacity proteins have no influence on T cell proliferation. Although unexpected, these observations suggest that human CD4+ T cells respond positively to one or more neisserial antigens to overcome any inhibition that could be induced via the engagement of CEACAM1.
As stated above, certain meningococcal Opa repertoires have been correlated with hyper-invasiveness and disease, however, they do not correlate with the severity of meningococcal disease (3). Although at present the molecular and cellular basis for such disease association is not entirely clear, in view of the current studies, it would seem premature to assign any disease association of meningococcal Opa proteins to their effects on T cell functions (11); perhaps the consequences of Opa/CEACAM interactions are engendered with a greater impact at other cellular interfaces (1, 13).
References
1. Griffiths NJ, Bradley, CJ, Heyderman RS, and Virji M. IFN-gamma amplifies NFkappaB-dependent Neisseria meningitidis invasion of epithelial cells via specific upregulation of CEA-related cell adhesion molecule 1. Cell Microbiol 2007; 9:2968.
2. Villullas S, Hill DJ, Sessions RB, Rea J, and Virji M. Mutational analysis of human CEACAM1: the potential of receptor polymorphism in increasing host susceptibility to bacterial infection. Cell Microbiol 2007; 9:329.
3. Callaghan, MJ, Rockett K, Banner C, Haralambous E, Betts H, Faust S, Maiden MC, Kroll JS, Levin M, Kwiatkowski DP, and Pollard AJ. Haplotypic diversity in human CEACAM genes: effects on susceptibility to meningococcal disease. Genes Immun 2008; 9:30.
4. Callaghan MJ, Jolley KA, and Maiden MC. Opacity-associated adhesin repertoire in hyperinvasive Neisseria meningitidis. Infect Immun 2006;74:5085.
5. Virji M, Makepeace K, Peak IR, Ferguson DJ, Jennings MP, and Moxon ER. Opc- and pilus-dependent interactions of meningococci with human endothelial cells: molecular mechanisms and modulation by surface polysaccharides. Mol Microbiol 1995;18:741.
6. Unkmeir A, Latsch K, Dietrich G, Wintermeyer E, Schinke B, Schwender S, Kim KS, Eigenthaler M, and Frosch M. Fibronectin mediates Opc-dependent internalization of Neisseria meningitidis in human brain microvascular endothelial cells. Mol Microbiol 2002;46: 933.
7. Virji M, Griffiths, NJ and Cunha CS. Opc and its multifarious receptors: identification of novel mechanisms that may operate for attachment and traversal of human endothelial cells. http://neisseria.org/ipnc/2008/Abstracts_oral_presentations_IPNC_2008.pdf (Abstract #055).
8. Cunha CS, Griffiths NJ, Murillo I, and Virji M. Neisseria meningitidis Opc invasin binds to the cytoskeletal protein alpha-actinin. Cell Microbiol 2009; 11: 389.
9. Robinson K, Wooldridge KG, Wells DB, Hasan A, Todd I, Robins A, James R, and Ala'Aldeen DA. T-cell-stimulating protein A elicits immune responses during meningococcal carriage and human disease. Infect Immun 2005;73: 4684.
10. Boulton IC, and Gray-Owen SD. Neisserial binding to CEACAM1 arrests the activation and proliferation of CD4+ T lymphocytes. Nat Immunol 2002;3: 229.
11. Lee HS, Boulton IC, Reddin K, Wong H, Halliwell D, Mandelboim O, Gorringe AR, and Gray-Owen SD. Neisserial outer membrane vesicles bind the coinhibitory receptor carcinoembryonic antigen-related cellular adhesion molecule 1 and suppress CD4+ T lymphocyte function. Infect Immun 2007;75: 4449.
12. Davenport V, Guthrie T, Findlow J, Borrow R, Williams NA, and Heyderman RS. Evidence for naturally acquired T cell-mediated mucosal immunity to Neisseria meningitidis. J Immunol 2003;171: 4263.
13. Muenzner P, Dehio C, Fujiwara T, Achtman M, Meyer TF, and Gray-Owen SD.. Carcinoembryonic antigen family receptor specificity of Neisseria meningitidis Opa variants influences adherence to and invasion of proinflammatory cytokine-activated endothelial cells. Infect Immun 20006;8: 3601.


{kind=link}
{kind=link}