










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkVaccimonitor
versión On-line ISSN 1025-0298
Vaccimonitor v.19 n.2 Ciudad de la Habana Mayo-ago. 2010
ARTICULOS ORIGINALES
Evaluación de la tolerancia a la crioconservación de dos cepas de Escherichia coli K12 de uso frecuente en biotecnología.
Assessment of cryopreservation tolerance of two Escherichia coli K12 strains of biotecnology frequent use.
Diliana Celeste Pérez-Reytor*, Angela Estela Sosa Espinosa**
Laboratorio Colección de Microorganismos de Interés Biotecnológico. Departamento de Seguridad y Ambiente. Centro de Ingeniería Genética y Biotecnología. P.O.Box 6162, Cubanacán, Playa, Ciudad de La Habana, Cuba. email:angela.sosa@cigb.edu.cu
* Licenciada en Microbiología; Máster en Ciencias Bioquímicas en la mención de Biología Molecular; Aspirante a investigador.
** Licenciada en Ciencias Biológicas; Master en Microbiología; Investigador Auxiliar.
RESUMEN
Uno de los métodos más recomendados para la conservación de E. coli es la congelación entre -20ºC y -70ºC, donde comúnmente se utiliza glicerol o dimetilsulfóxido como criopreservante. Los bancos de E. coli que se conservan por esta vía pueden mantener altas viabilidades por más de 10 años. A pesar de que es una práctica común están poco documentadas las diferencias que existen entre las cepas de interés biotecnológico en cuanto a la resistencia a la congelación. En este artículo se comparan dos cepas mutantes de E. coli K12: la cepa RRI y la cepa HB101, con el objetivo de evaluar el comportamiento de ambas a dos temperaturas de almacenamiento y diferentes concentraciones de glicerol, después de ocho ciclos sucesivos de congelación-descongelación. Las cepas estudiadas sólo difieren en su capacidad de reparación al daño sobre el ADN al presentar la HB101 una variante mutada del gen recA. Cuando se utilizó una temperatura de almacenamiento de -70 ºC y diferentes concentraciones de criopreservante las cepas tuvieron un comportamiento similar de supervivencia, después de los ocho ciclos. Sin embargo, el comportamiento de la supervivencia difiere cuando la temperatura de congelación en los ciclos es de -20 ºC. Además de la supervivencia y la estabilidad de los marcadores genéticos estudiados, observamos que el efecto de los ciclos de congelación y descongelación retardó el tiempo de aparición de las colonias en medio sólido.
Palabras clave: Crioconservación E. coli, congelación-descongelación E. coli.
ABSTRACT
One of the most recommended methods for E. coli preservation is freezing from -20 ºC to -70 ºC. Glycerol or dymethylsulfoxide as cryoprotectant are commonly used for this purpose. E. coli banks conserved in this way maintain high viability for more than ten years. Though E. coli cryopreservation is a common practice, freezing resistance differences between strains of biotechnological interest are poorly documented. This article compares two mutant strains of E. coli K12, RRI and HB101, in order to evaluate their behavior at both different storage temperatures and glycerol concentrations after eight successive cycles of freezing-thawing. These isolates only differ in their ability to repair DNA damage due to a recA gene mutation in the HB101 variant. When a storage temperature of -70 ºC and different concentrations of cryoprotectant was used the strains showed similar survival after eight cycles. However, the survival behavior differs when the freezing temperatures in the cycles was -20 ºC. In addition to the survival and stability of the genetic markers studied, we observed that the effect of the freezing-thawing cycles delayed their growth on solid medium.
Keywords: Cryoconservation E. coli, freezing-thawing E. coli.
INTRODUCCION
Existen diferentes métodos para la preservación de cultivos microbianos. De estos, la congelación sigue siendo una de las principales formas utilizadas hasta la fecha para el almacenamiento de cepas. La garantía de la conservación de los recursos microbianos depende en gran medida de la estabilidad de los microorganismos a la congelación cuando se almacenan cultivos puros y homogéneos en condiciones que aseguren su viabilidad y estabilidad genética (1).
El método de preservación utilizado debe permitir altas viabilidades, prevenir cambios en las características morfológicas y bioquímicas, en la secuencia nucleotídica y en la estabilidad plasmídica de las cepas. La selección de este método depende en gran medida de las disposiciones económicas, posibilidad de obtener cultivos puros y el propósito del laboratorio.
En las colecciones de cultivo se recomienda utilizar dos métodos diferentes para el almacenamiento de los bancos. Otros criterios se tienen en cuenta y dependen de cada microorganismo en particular, así existen opiniones diversas sobre: la viabilidad que debe alcanzar el cultivo, la fisiología del cultivo en el momento de la conservación, la estabilidad genética, los costos del proceso y la frecuencia de uso del recurso (2).
Para preservar mejor las cepas contra el daño por congelación se emplean diversos agentes crioprotectores. Estas moléculas mejoran la estabilidad de las cepas al reducir al mínimo el contenido de agua intracelular, evitar la vitrificación y proteger las macromoléculas en el ambiente interno de la célula (3).
Estos compuestos se han clasificado teniendo en cuenta su permeabilidad a la célula. Los llamados compuestos penetrantes son un grupo de moléculas de alto peso molecular como: dimetil sulfóxido (DMSO), glicerol, metanol y etilenglicol; se aplican generalmente en altas concentraciones (0,5-1,5 M). Otro grupo de compuestos son llamados no penetrantes, donde están: el almidón, el hidroxietílico (HES), la polivinil pirrolidina (PVP), azúcares, proteínas y diferentes moléculas de polietilenglicol que se utilizan a bajas concentraciones (0,01 M) (3).
Como práctica común, en muchos laboratorios biotecnológicos se utiliza el almacenamiento de microorganismos desde -20 a -70 ºC . Los altos niveles de supervivencia alcanzados por este método son de interés, tanto desde el punto de vista biofísico como práctico. Sin embargo, estos niveles de supervivencia varían entre las diferentes especies y géneros microbianos, por lo que el estudio de la estabilidad a la congelación es un paso obligado de la conservación de cepas microbianas particulares (4).
En el caso de cepas de interés de E. coli los efectos del estado fisiológico del cultivo y su susceptibilidad a la muerte por congelación y descongelación repetida se estudiaron por primera vez por Harrison y colaboradores (5), quienes informaron que cultivos crecidos aeróbicamente eran más resistentes que los cultivos crecidos en condiciones anaerobias. Ciclos sucesivos de congelación y descongelación de cultivos de cepas de E. coli cultivadas en medio mínimo y congeladas en el mismo medio, al cual se le ha añadido una fuente de carbono, trae como resultado un decrecimiento lineal en el logaritmo del número de células viables en función del número de ciclos, siendo la pendiente de la curva un índice de susceptibilidad del cultivo.
Derivados de estas observaciones existen diferentes criterios interlaboratorios, en cuanto al tipo y la concentración de criopreservantes, el intervalo de tiempo que se puede establecer para las verificaciones de los bancos, el momento fisiológico para detener el crecimiento del cultivo y el medio de conservación a utilizar.
En 1970 Lapage y colaboradores (6) proponen concentraciones de glicerol del 40% y temperaturas entre -10 a -20 ºC para la conservación de células bacterianas, criterio que es retomado por Miller en 1972 (7). En la metodología se propone añadir el glicerol directamente a un cultivo crecido hasta la etapa estacionaria.
La Colección de Cultivos Americana (ATCC) (8) utiliza preferentemente el DMSO como criopreservante, mientras que la compañía New England Biolabs (NEB) (9) propone 50% de glicerol cuando se conserva a -70 ºC. Estos son solo algunos ejemplos de la diversidad de métodos de conservación.
Con vistas a homogeneizar los criterios de elección de un método válido de conservación para las cepas de E. coli que se conservan en la Colección de Microorganismos de Interés Biotecnológico, en este trabajo se estudiaron dos cepas de E. coli que solo difieren en su genotipo en la presencia o no de una mutación en el gen recA, cuyo producto génico es una enzima reparadora involucrada en la respuesta SOS de la célula. Para la selección de los criterios de elección se tuvieron en cuenta diferentes métodos de conservación que se emplean comúnmente en los laboratorios del Centro de Ingeniería Genética y Biotecnología.
La tolerancia de las cepas se evaluó a dos temperaturas de almacenamiento y diferentes concentraciones de glicerol como preservante; tomando como criterios de estabilidad a la congelación: la supervivencia del cultivo después de ocho congelaciones y descongelaciones sucesivas, la observación de la estabilidad de los marcadores genéticos listados en sus genotipos y la capacidad de recuperación del cultivo, medida como la habilidad de aparición de las colonias en un medio sólido de propósitos generales.
MATERIALES Y METODOS
Material biológico
Las cepas de E. coli K12 que se utilizaron en este estudio fueron: la cepa RRI que presenta mutaciones en los genes hsdS20, supE44, ara-14, leuB6, proA2, lacY1, rpsL20, xyl-5, mtl-1, galK2 y se comporta como receptora en la conjugación bacteriana, y la cepa HB101, de igual genotipo, pero con una mutación adicional en el gen recA (9), procedentes de la Colección de Microorganismos de Interés Biotecnológico del Centro de Ingeniería Genética y Biotecnología.
Confección de los bancos de trabajo
A partir de colonias aisladas en placas de medio Luria-Bertani (LB) se inocularon Erlenmeyers que contenían 50 mL del mismo medio (10). Los cultivos se incubaron con una agitación de 250 r.p.m. a 37 ºC durante 8 h. Después de este tiempo los Erlenmeyers fueron incubados en hielo durante 10 min y luego se colectó la biomasa por centrifugación a 3 000 r.p.m. y 4 ºC durante 15 min. Concluida la centrifugación los sobrenadantes fueron decantados en condiciones asépticas y los precipitados se resuspendieron en 25 mL de medio LB, al cual se añadieron concentraciones diferentes de glicerol (0; 10,96; 15 y 30%). Cada concentración de glicerol se distribuyó en 10 viales a razón de 1 mL por vial y se almacenaron a -20 y -70 ºC.
Ciclos de congelación-descongelación
Después de 24 h de congelación se realizaron ocho ciclos sucesivos de descongelación por 20 min en hielo y congelación durante 1 h a las temperaturas de -20 ºC ó -70 ºC, según la temperatura de almacenamiento del banco. Después de descongelar en cada ciclo fue extraído un vial del banco y se hicieron diluciones seriadas desde 10-1 hasta 10-6 que se sembraron por el método de traza de dilución (11) sobre medio LB, las placas fueron incubadas a 37 ºC hasta que las colonias fueron visibles. Cada ensayo fue replicado tres veces y se calculó la media del número de viables y la desviación estándar en cada experimento. Los datos se analizaron por regresión lineal y el valor de la pendiente fue tomado como la cantidad de viables que sobrevive por ciclo de descongelación (12).
Chequeo fenotípico de las cepas
Después de la confección del banco y en el octavo ciclo de descongelaciones sucesivas fue verificado el genotipo de ambas cepas por siembra en placas que contenían diferentes nutrientes añadidos al medio M9 (10). Las variantes empleadas para la verificación de auxotrofias fueron: medio M9 con adición de glucosa (0,2%) y leucina (25 mg/mL), medio M9 con adición de glucosa (0,2%) y prolina (25 mg/mL) y medio M9 con adición de glucosa (0,2%), leucina y prolina (25 mg/mL). Para la verificación de afectación
catabólica las variantes empleadas fueron: medio M9 con adición de leucina y prolina a una concentración final de 25 gm/mL, con adición de 0,2% de galactosa; medio M9 con adición de leucina y prolina a una concentración final de 25 mg/mL, con adición de 0,2% de melibiosa, y medio M9 con adición de leucina y prolina a una concentración final de 25 mg/mL, con adición de 0,2% de xilosa. En todos los casos se evaluó la habilidad de las cepas de crecer en estos medios. Las placas fueron crecidas a 37 ºC hasta la visualización de las colonias. Cada ensayo se replicó tres veces y se calculó la media y la desviación estándar en cada experimento.
RESULTADOS Y DISCUSION
La gran variedad de métodos propuestos para la conservación de E. coli dificulta tener un criterio único de elección a la hora de conservar cepas de interés. Una incertidumbre adicional es la duda de si un método de conservación recomendado para la especie será efectivo para cepas mutantes de esta misma especie, ya que existe la posibilidad de que tengan una capacidad diferente de mantener alta supervivencia en las condiciones propuestas.
Los estudios realizados a cepas de E. coli para evaluar la resistencia a la congelación se han basado en la aplicación a un cultivo de ciclos sucesivos de congelación y descongelación, lo que resulta en un decrecimiento en el número de viables proporcional al número de ciclos del ensayo (12). Si este método permite evaluar la capacidad del microorganismo de tolerar la congelación puede ser utilizado para estudiar una condición de almacenamiento y determinar si es adecuada o no, o si existen cepas de la misma especie más sensibles a una condición determinada, ya que en este caso la disminución de las células viables por ciclo debe ser mayor.
Para conocer si dos cepas de E. coli tenían el mismo comportamiento a la conservación se evaluó la afectación del cultivo durante ocho ciclos de congelación-descongelación. La selección de los criterios se hizo teniendo en cuenta los valores propuestos por diferentes autores y las metodologías utilizadas en los laboratorios del Centro de Ingeniería Genética y Biotecnología para la conservación de cepas de E. coli.
Después de ocho ciclos de congelación a -70 ºC y descongelación lenta en hielo, el porcentaje de supervivencia de ambas cepas fue mayor cuando se utilizó una concentración de glicerol del 10,96% (Tabla 1), lo que coincide con la concentración que recomiendan Hill y colaboradores (13) cuando se utilizan criopreservantes penetrantes. A concentraciones de 30% de glicerol la supervivencia se afectó de forma drástica en ambas cepas; se observa un efecto mayor de la concentración sobre la cepa HB101.

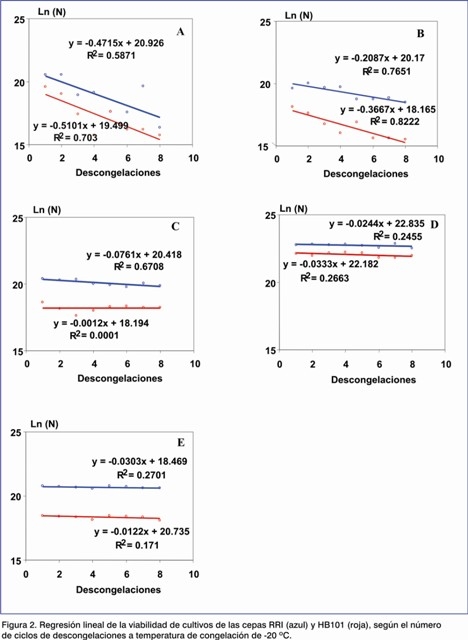
Las gráficas de regresión lineal de los valores de viabilidad obtenidos muestran que la afectación de la viabilidad en ambas cepas está en función del número de las descongelaciones a todas las concentraciones de criopreservantes que se ensayaron (Figura 1). Esto concuerda con lo planteado por otros autores (12).
Los diferentes métodos de conservación difieren sus propuestas con relación a dos factores que se conocen influyen en la viabilidad de los cultivos crioconservados: la concentración de criopreservante y la temperatura de almacenamiento. Debido a esto el mismo procedimiento de análisis fue utilizado variando la temperatura de almacenamiento a -20 ºC.
Las gráficas de regresión lineal muestran que la supervivencia de la cepa RRI en esta condición dependió del aumento de la concentración de glicerol, mientras que la HB101 mantuvo un comportamiento similar al observado cuando utilizamos una temperatura de -70 ºC (Figura 2).
Los niveles de la viabilidad después de la congelación pueden variar de acuerdo con numerosos factores, entre los que se incluyen la eficacia del agente crioprotector y las propiedades de la cepa en particular (13).
A pesar de todos los efectos potencialmente perjudiciales de la congelación las poblaciones bacterianas pueden protegerse a sí mismas de muchos de ellos o recuperarse bajo las circunstancias adecuadas (14).
La cepa HB101 tiene afectada su capacidad de reparación de daños del ADN por la mutación en el gen recA. Esta diferencia con la cepa RRI puede ser la causa del comportamiento desigual de las cepas en una misma condición de almacenamiento. Adicionalmente, la concentración de glicerol adecuada varió entre las dos cepas cuando la temperatura de congelación fue de -20 ºC. Para la cepa RRI la concentración de criopreservante, en la que se observó mayor supervivencia, fue del 30%, mientras que en la cepa HB101 se mantuvo una supervivencia del 100%, entre 10,96 y 15% de glicerol (Tabla 2). Este resultado sugiere que la capacidad de la cepa de recobrarse después del almacenamiento depende de otras características individuales que aparentemente son independientes del fenotipo RecA. Algunos autores plantean que la exposición a las tensiones en frío, osmótica y otros, pueden aumentar posteriormente la supervivencia a los ciclos de congelación-descongelación, presumiblemente debido a los cambios en los niveles de determinadas proteínas o a otros componentes celulares que brindan tolerancia (15).

Algunos autores plantean que la congelación y la descongelación causan daño oxidativo y por lo tanto puede ser mutagénico (16, 17). En ninguna de las dos cepas, cuando se comprobó el fenotipo verificable como criterio de estabilidad genética, encontramos pérdida de ninguno de los marcadores listados en su genotipo al cabo de las ocho descongelaciones. Sin embargo, la habilidad de crecimiento de ambas cepas en medio sólido LB después de cada descongelación fue diferente. Para el caso de la RRI se obtuvo crecimiento después de 16 h de incubación a 37 ºC, mientras que las colonias de la cepa HB101 eran visibles entre las 24 y las 36 h de incubación a esta misma temperatura y se observó que en cada ciclo se demoraba más el momento de aparición de las colonias. Este comportamiento sugiere que el daño que sufren las células durante la congelación puede ser muy diverso y que no solo se manifiesta en una disminución de la viabilidad del cultivo o el cambio de marcadores genéticos conocidos. Además, esta diferencia en la habilidad de crecimiento en medio sólido puede estar influenciada por la mutación recA presente en la cepa HB101, ya que no se observó en la cepa RRI.
No se descarta la posibilidad de que aumentando el número de ciclos de congelación y descongelación se puedan encontrar evidencias de posibles cambios en el genoma.
Existen estudios sobre la evolución de poblaciones de E. coli a largo plazo hechos por Lienski y colaboradores (18, 19, 20), pero opinamos que la simulación con ocho ciclos es suficiente para los propósitos de esta investigación. El estudio de viabilidad durante 10 años de almacenamiento a temperatura estable de -70 ºC no ha mostrado variación en la viabilidad de ninguna de las dos cepas.
Nuestros resultados sugieren que no existe una condición única en cuanto a la concentración de criopreservante aplicable a los mutantes de E. coli cuando la temperatura de almacenamiento es de -20 ºC. A una temperatura de -70 ºC es posible que la conservación sea independiente del genotipo de la célula y dependa más de la metodología de conservación.
REFERENCIAS
1. Miyamoto-Shinohara Y, Sukenobe J, Imaizumi T, Nakahara T. Survival of freeze-dried bacteria. Journal of General and Applied Microbiology 2008; 54(1):9-24.
2. Kirsop BE, Doyle A. "100 Years of Culture Collections". Proceedings of the Kral Symposium to Celebrate the Centenary of the Establishment of the Firs Recorded Service Culture Collection. OSAKA, Japan: Institute of Fermentation, WFCC; 1990.
3. Hubalek Z. Protectants used in the cryopreservation of microorganisms. Cryobiology 2003,46(3):205_9.
4. Meza RA, Monroy AF, Mercado M, Poutou RA, Rodríguez P, Pedroza AM. Study of the stability in real time of cryopreserved strains banks. Universitas Scientiarum 2004; 9(2):35-42.
5. Harrison AP. Survival of bacteria upon repeated freezing and thawing. Journal of Bacteriology 1955;70:711-5.
6. Lapage SP, Shelton JE, Mitchell TG, Mackenzie AR. Culture collection and the preservation of bacteria. In: Methods in Microbiology. London: Academic Press; 1970. p.135-228.
7. Miller GH. Experiments in Molecular Genetics. 3th Edit. New York: Cold Spring Harbor Laboratory; 1977.
8. http://www.atcc.org/ATCCAdvancedCatalogSearch/tabid/112 /Default.aspx. US, 2009.
9. http://www.neb.com/nebecomm/tech_reference/competent_cells/shufflestrains.asp. UK; 2010.
10. Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual. 2nd Edit. New York: Cold Spring Harbor, Laboratory Press; 1989.
11. Pérez-Reytor DC, Domínguez I, Sosa AE. Evaluación del método de siembra en placa traza de la dilución en el control de calidad de bancos de mutantes de Escherichia coli K12. Biotecnología Aplicada 2002;19 (3):169-73.
12. Sleight SC, Wigginton NS, Lenski RE. Increased susceptibility to repeated freeze-thaw cycles in Escherichia coli following long-term evolution in a benign environment. BMC Evolutionary Biology 2006. Disponible en: http://www.biomedcentral.com/1471-2148/6/104.
13. Hill LR, Kokur M, Malik KA. Culture and Maintenance. In: Hill LR and Kirsop BE, eds. Living resource for Biotechnology. Bacteria. London: Cambridge University Press; 1991. p. 62-80.
14. Sleight CS, Lenski RE. Evolutionary adaptation to freeze-thaw-growth cycles in Escherichia coli. Physiological and Biochemical Zoology 2007; 80: 370_85.
15. Sleight SC, Orlic C, Schneider D, Lenski RE. Genetic basis of evolutionary adaptation by Escherichia coli to stressful cycles of freezing, thawing and growth. Genetics 2008,180:431-43
16. Calcott PH, Gargett AM. Mutagenicity of freezing and thawing. FEMS Microbiol Lett 1981;10:151-55.
17. Grecz N, Hammer TL, Robnett CJ, Long MD. Freeze-thaw injury: evidence for double strand breaks in Escherichia coli DNA. Biochem Biophys Res Commun 1980;93:1110-31.
18. Lenski RE, Winkworth CL, Riley MA. Rates of DNA sequence evolution in experimental populations of Escherichia coli during 20,000 generations. Journal of Molecular Evolution 2003; 56: 498_508.
19. Panoff JM, Thammavongs B, Gueguen M. Cryoprotectants Lead to Phenotypic Adaptation to Freeze-Thaw Stress in Lactobacillus delbrueckii ssp. Bulgaricus. CIP Cryobiology 2000; 40:264-9.
20. Grecz N, Hammer TL, Robnett CJ, Long MD. Freeze-thaw injury: evidence for double strand breaks in Escherichia coli DNA. Biochemical and Biophysical Research Communications 1980; 93:1110-3.
Recibido: Diciembre de 2009
Aceptado: Febrero de 2010


{kind=link}
{kind=link}