










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkVaccimonitor
versión impresa ISSN 1025-028X
Vaccimonitor vol.20 no.2 Ciudad de la Habana Mayo.-ago. 2011
ARTICULOS ORIGINALES
Adjuvant activity of rice oil on the immune response to ovalbumin
Actividad Adyuvante del aceite de arroz en la respuesta inmune contra ovoalbúmina
Isabela Thommen Maciel Sartor,1 Edson Moleta Colodel,1 Deijanira Albuquerque2*
1 Department of Medical Clinical Veterinary, Faculty of Agronomy, Veterinary Medicine and Zootechnics, Brazil.
2 Department of Basic Health Science, Faculty of Medicine, Federal University of Mato Grosso, Avenida Fernando Corrêa da Costa, 2.367 Boa Esperança, 78060-900 Cuiabá, Mato Grosso, Brazil. email:albuquerqued@yahoo.com
* Prof. Dr. Deijanira Alves de Albuquerque Ph.D. Department of Basic Health Science, Faculty of Medicine, Federal University of Mato Grosso, Avenida Fernando Corrêa da Costa, 2.367 Boa Esperança, 78.060-900 Cuiabá, Mato Grosso, Brazil. Phone: +55 65 36158847.
ABSTRACT
We have evaluated the adjuvant activity of rice oil (RO) on the immune response to ovalbumin (OVA), its depots and possible side effects at the injection sites. Immunization of mice with OVA emulsified in rice oil (OVA+RO) resulted in an antibody response significantly higher than that determined in mice immunized with soluble OVA. In addition, these high antibody levels were observed for a period as long as 6 weeks after immunization. However, the adjuvant action of RO was significantly lower to incomplete Freund's adjuvant (IFA). In secondary immune response, the pattern of OVA-specific antibodies stimulated by RO was predominantly IgG1. Despite of the significant humoral response enhanced by RO at no time was noted any lesions at the site of injection. Besides, histological analysis showed that inflammatory reaction caused by RO was mild and transient, suggesting that RO appears to be a safe, effective and chemically define alternative to IFA in many situations.
Keywords: Rice oil, adjuvants, antibody response, vaccine, cytokines.
RESUMEN
Evaluamos al aceite de arroz (AA) como adyuvante para la respuesta inmune de ovoalbúmina (OVA), su depósito y los efectos colaterales posibles en el lugar de la inyección. La inmunización de ratones con OVA emulsionada en aceite de arroz (OVA+AA) tuvo como resultado una respuesta de anticuerpos significativamente mayor que la determinada en ratones inmunizados con OVA soluble. Además, estos altos niveles de anticuerpos se observaron durante seis semanas después de la inmunización. Sin embargo, la acción adyuvante del AA fue significativamente menor a la del adyuvante incompleto de Freund (AIF). En la respuesta inmune secundaria, el patrón de anticuerpos específicos contra OVA, estimulados por AA, fue predominantemente IgG1. A pesar del significativo aumento de la respuesta humoral inducido por AA, no se observaron lesiones en el lugar de la inyección. Además, el análisis histológico mostró que la reacción inmunológica causada por AA fue ligera y temporal, sugiriendo que el AA parece ser una alternativa segura, eficaz y químicamente definida para el AIF en muchas situaciones.
Palabras clave : Aceite de arroz, adyuvantes, respuesta de anticuerpos, vacunas, citoquinas.
INTRODUCTION
Adjuvants such as Freund's preparations are efficient in extraordinarily increasing the intensity of the immune response to a variety of soluble and particulate antigens. However, the complete Freund adjuvant (CFA) can cause fever and abscess formation at the site of injection (1), a situation which renders it unsatisfactory for use in human and veterinary vaccines (2,3).
In addition, it has been clearly documented that CFA can also cause sever chronic pain and suffering in small animals as well (1,4). With advances in many areas of biological sciences and increasing concern for the welfare of experimental animals, nowadays the use of CFA has been restricted and in some countries it is already forbidden for animals use also. Incomplete Freund adjuvant (IFA) is still wildly used for veterinary vaccines (5,6). However, IFA also causes severe and large inflammatory reactions at the injection sites and consequently economic lose (7,8). Therefore, whenever possible alternatives to Freund's preparations should be used.
Mineral oil is used in CFA and IFA because previous investigators have been unable to produce stable emulsions with vegetable oils. One exception is the adjuvant 65 which uses peanut oil with aluminium monostearate as stabilizer (9). The efficacy of adjuvant 65 is comparable to that of IFA. Few years ago, the Silva Lima's group demonstrated that stable water-in-oil emulsions can be prepared with soybean, corn, cotton seed and rice oils (10,11). In this work, we describe the adjuvant activity of rice oil from Oryza sativa L. on the antibody response to OVA in mice. We also evaluated IL-4 and IFN-g production in mice immunized with OVA+RO or ovalbumin emulsified in IFA (OVA+IFA), oil deposition and inflammatory reaction in mice injected with RO emulsified in saline. The rationale for the use of RO as adjuvant was fundamentally based on the fact that it is metabolizable and produces a stable water-in-oil emulsion (11). Besides, RO has long been used in human dietary without reported side effects.
MATERIALS AND METHODS
Animals
The 9-week-old C57BL/6 mice used in this study were obtained from Oswaldo Cruz Foundation, Salvador, BA. The animals were kept in micro-isolators and all experiments were performed according to the institutional ethical guidelines on the use of animals in research.
Antigen and adjuvants
Ovalbumin, (OVA, grade V) and IFA were obtained from Sigma-Aldrich Brazil Ltda. Commercial edible RO was obtained from Bleu Ville R Santalucia S.A. Company.
Emulsion preparation
Water-in-oil emulsions of RO or IFA were prepared according to Moncada, et al. (12) with a slight modification for the use of glass syringes. In short: 0.5 mL of soluble OVA (0.1 mg/mL) was added to 0.5 mL of oil phase and emulsified to a water-in-oil emulsion. Stability of emulsion of RO or IFA was assessed by placing a drop of the emulsions onto the surface of distillated cold water as previous described (13).
Immunizations
Mice were immunized once with OVA (10 mg) emulsified with RO or IFA. Control animals received 10 mg of soluble OVA and 21 days later all mice received one booster with 10 mg of soluble OVA. All immunizations were done by subcutaneous (sc) route in a total volume of 0.2 mL per animal. All experiments were repeated twice.
ELISA assay
Anti-OVA antibodies were assayed by standard ELISA procedures, as previously described (14). In short, polystyrene plates (Falcon) were coated overnight at 4 °C with 0.5 mg of OVA per well, washed with PBS 0.05%-Tween-20, blocked with 1%-BSA/PBS for 1h at room temperature and coated with serial dilutions of each mouse antiserum starting at 1/100. After overnight incubation at 4 °C, the plates were washed, incubated for 1 h at 37 °C with goat antisera to mouse immunoglobulins (all isotypes, IgG1 or IGg2a subclasses) conjugated to peroxidase (Zymed), washed and developed by addition of H 2 O 2 and orthophenylenediamine (OPD). The reaction was stopped by addition of H 2 SO 4 and read in a Microplate Reader, Bio-Rad R Laboratories. The optical density (DO) was measured in an ELISA reader at 490nm, the results are expressed as the means (DO) ± SD.
Ex vivo supernatants
The ex vivo supernatants were obtained from spleens of mice immunized with OVA+RO or OVA+IFA. Briefly, six weeks after the primary immunization all mice received a second boost with soluble OVA (10 mg). Three days latter the animals were sacrificed, their spleens collected, minced in complete culture medium (Nutricel R ) and centrifuged at 4 °C to obtain ex vivo supernatants.
Measurement of cytokines
Cytokine levels in culture supernatants were determined by ELISA, using monoclonal antibody pairs and recombinant cytokines purchased from PharMingen R , as previously described (15).
Footpad histology
C57BL/6 mice were injected into a footpad with RO emulsified in saline. The contralateral footpad received only saline and served as control. Animals received a total volume of 40 mL into each footpad. Three days latter, the mice were sacrificed, their feet removed and immersed in 10% phosphate-buffered formalin for 24 h. Tissues were decalcified, sliced, imbedded in paraffin and 4 µm sections were stained with hematoxylin/eosin for light microscopic examination. Evaluations of oily deposits and possible adverse effects of RO at the injection sites were done with coded samples in a blind study.
Statistical analysis
ELISA results were analyzed using one-way analysis of variance (ANOVA) followed by the Tukey test to determine significant differences among groups. Values of P <0.05 were considered statistically significant.
RESULTS
Stimulation of OVA-specific antibody response by RO
To determine the capacity of RO as an adjuvant, C57BL/6 mice (four per group) were immunized sc with OVA+RO (10 mg/animal). For comparative purposes, mice were injected with OVA+IFA or received soluble OVA. The mice were bled weekly and their sera were evaluated for OVA-specific antibodies by ELISA. Data in Figure 1 show that mice immunized with OVA+RO developed a secondary antibody response significantly higher than that of the mice injected with soluble OVA. However, the OVA-specific antibody levels stimulated by RO were significantly lower than those stimulated by IFA.
Antibody subclasses stimulated by RO
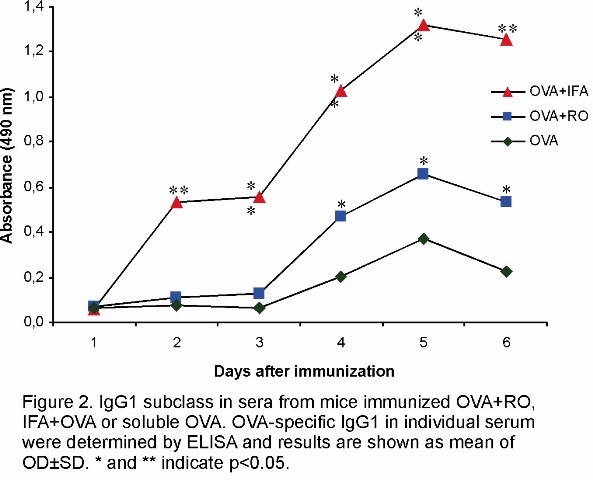
Sera from mice immunized with OVA+RO were also tested for the presence of the OVA-specific IgG1 and IgG2a subclasses. As shown in Figure 2, RO as well as IFA stimulated high levels of IgG1. Under the same experimental conditions IgG2a was not detected.
Evaluation of IL-4 and IFN-g in ex vivo supernatants
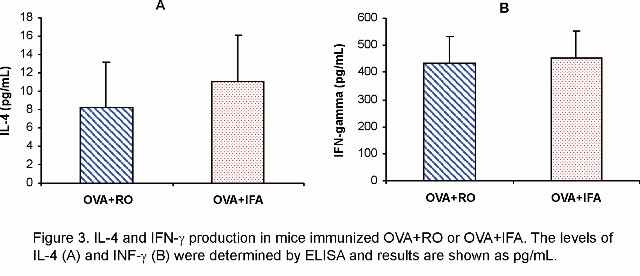
As RO stimulated the OVA-specific antibody response, including IgG1, we next verified whether this oil would enhance IL-4 production, since this cytokine plays an important role on IgG1 synthesis (16,17). The data show that RO as well as IFA increased IL-4 production (Figure 3A). Under the same conditions, both adjuvants RO and IFA also enhanced IFN-g synthesis (Figure 3B). IL-4 and IFN-g were undetectable in ex vivo supernatants from spleens of non immunized mice (data not shown).
Footpad histology
Since RO enhanced OVA-specific antibody production, IL-4 and IFN-g, we further investigated whether this oil would remains at the site of injections. Histological sections from representative sites of injections demonstrated oily deposits at the interstitial space (Figure 4A, arrows). We also analyzed the same sections for inflammatory reaction caused by this oil. Note that RO induced only a mild inflammatory response, mainly characterized by influx of leukocytes (Figure 4A). Figure 4B shows a tissue section from a footpad that was injected with saline alone and served as control.
DISCUSSION
Previous studies have reported that the adjuvant 65, which contains peanut oil, which is a metabolizable oil, stimulates antibody response comparable to that of IFA (9). Few years ago the Silva Lima´s group showed that soybean, corn, cotton seed and rice oils were able to produce stable water-in-oil emulsions without adding any stabilizer (10, 11). In this study, we showed that water-in-oil emulsions of RO subcutaneously injected in mice remains at the injection sites and cause only a mild inflammatory reaction. Besides, immunization of mice with OVA+RO resulted in a secondary antibody response to OVA that was significantly higher than that observed in mice injected with soluble OVA. To assess the magnitude of adjuvant action of RO, mice were immunized with OVA+RO or OVA+IFA. Analysis of sera from both animal groups showed that the levels of OVA-specific antibodies enhanced by RO were significantly lower than those obtained when IFA was the adjuvant (Figure 1). When comparing adjuvant actions of RO versus IFA, the lower adjuvant activity of RO may be accounted for its metabolizable property.
We have also evaluated the subclasses pattern of IgG to OVA stimulated by RO. The results also showed that, during secondary response RO preferentially stimulated IgG1 production. In this regard, the action of RO is similar to that IFA, which mainly stimulates the IgG1 subclass in mice. It has been previously demonstrated that in mice, IgG1 production is partially IL-4-independent (18, 19). Other authors have reported that regarding to the anaphylactic activity of IgG1, mice produce two subclasses of IgG1: one which is anaphylactic and IL-4-dependent and the other which is non-anaphylactic and is IL-4-independent (20). In our study, we have showed that, under the same conditions RO as well as IFA enhanced both IL-4 and IFN-g production by spleen cells from mice immunized with OVA+RO or OVA+IFA (Figure 3A and 3B, respectively). As we did not determined whether the IgG1 stimulated by RO was or not anaphylactic, we would speculate that RO may elicit both, Th1 and Th2 response.
Despite of significant adjuvant action of RO, the inflammatory reaction induced by this oil was mild and transient. This event also can be accounted for its metabolizable property.
Although our data show that RO enhances IL-4, IFN-g and OVA-specific antibodies production, they do not identify underlying mechanism by which this occurs. At least one would speculate that RO might be another type of depot-acting adjuvant. This hypothesis is supported by the fact that oily depots were observed at the site of injections (Figure 4A).
The observation that RO has adjuvant action on antibody response in vivo is an important contribution to studies on the use of metabolizable oils as adjuvants. This oil constitutes an important addition to the list of vegetable oils already employed to stimulate antibody response in absence of mineral oil.
ACKNOWLEDGEMENTS
The work was supported by grants from the National Council of Research and Technology (CNPq).
REFERENCES
1. Broderson JR. A retrospective review of lesions associated with the use of Freund's adjuvant. Lab Anim Sci 1989;39:400-5.
2. Claasen E, Leeuw W, Greve P, Hendriksen C, Boersma W. Freund's complete adjuvant: an effective but disagreable formula. Res Immunol 1992;143:478-83.
3. Mbow ML, Gregorio E, Valiante NM, Rappuoli R. New adjuvants for human vaccine, Current Opinion in Immunol 2010;22: 411-6.
4. Amyx HL. Control of animal pain and distress in antibody production and infectious disease studies. 1987; 191:1287-9.
5. Lambrecht BN, Kool M, Willart MAM, Hammad H. Mechanism of action of clinically approved adjuvants. Current Opinion in Immunology 2009; 21:23-9.
6. Reed SG, Bertholet S, Coler RN, Friede M. New horizons in adjuvants for vaccine development. Trends in Immunol 2009;30:23-32.
7. Moro E, Junqueira JOB. Levantamento da incidência de reações vacinais e/ou medicamentosas em carcaças de bovinos ao abate em frigoríficos no Brasil. A hora Vet 1999;122:74-7.
8. França AT, Alves GG, Mesquita AJ, Chiquetto CE, Bueno CP, Oliveira ASC. Perdas econômicas por abscessos vacinais e/ou medicamentosos em carcaças de bovinos abatidos no estado de Goiás. Ciência Animal Brasileira 2006;7(1):93-6.
9. Hilleman MR, Woodhour AF, Friedman A, Weibel RE, Stokes J. The clinical application of adjuvant-65. Ann Allergy 1972;30:152-8.
10. Silva ACM, Guerra JL, Melo DF, Lima MS. Adjuvant effect of corn and olive oils in mouse immune response. In: VI Pharmatech, III Annais of the SBTF Annual Meeting. Recife, Brazil: UFPE; 2001. p. 165-6.
11. Mota EF. Atividade adjuvante dos óleos de amendoim (Arachis hypogaea L), algodão (Gossipium hirsutum L) e arroz (Oryza sativa L) na resposta imune humoral e celular, Ph.D. Thesis, Departamento de Bioquímica e Biología Molecular, Universidade Federal do Ceará (UFC), Fortaleza, CE, Brazil; 2006.
12. Moncada C, Torres V, Israel Y. Simple method for the preparation of antigen emulsions for immunization. J Immunol Methods1993;162:133-40.
13. Becher P. Testing and emulsion properties, Emulsions Theory and Practice, second ed. New York: Reinhold; 1965
14. Aroeira LS, Cardillo F, De Albuquerque DA, Vaz NM, Mengel J. Anti-IL-10 treatment does not block either the induction or the maintenance of orally induced tolerance to OVA. Scand J Immunol 1995; 41:319-23.
15. Albuquerque DA, Saxena V, Adams DE, Boivin GP, Brunner HI, Witte DP. et al. An ACE inhibitor reduces Th2 cytokines and TGF-b1 and TGF-b2 isoforms in murine lupus nephritis. Kidney Int 2004;65:846-59.
16. Snapper CM, Finkelman FD, Paul WE. Regulation of IgG1 and IgE production by interleukin 4. Immunol Rev 2004;102:51-75.
17. McKenzie GJ, Fallon P, Emsom CL, Grencis R, McKenzie AN. Simultaneous disruption of interleukin (IL)-4 and IL-13 defines individual roles in T helper cell type 2-mediated responses. J Exp Med 1999;189:1565-72.
18. Brewer JM, Conacher M, Satoskar A, Bluethmann H, Alexander J. Interleukin-4-deficient mice, alum not only generates T helper 1 responses equivalent to freund's complete adjuvant, but continues to induce T helper 2 cytokine production. Eur J Immunol 1996;26:2062-6.
19. Grun JL, Maurer PH. Different T helper cell subsets elicited in mice utilizing two different adjuvant vehicles: the role of endogenous interleukin 1 in proliferative responses. Cell Immunol 1989;121:134-45.
20. Silva AS, Cavalcante LT, Faquim-Mauro EL, Macedo MS. Regulation of anaphylactic IgG1 antibody production by IL-4 and IL-10. Int Arch Allergy Immunol 2006;141:70-8.
Recibido: Diciembre de 2010
Aceptado: Marzo de 2011


{kind=link}
{kind=link}
{kind=link}
{kind=link}