










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkVaccimonitor
versión impresa ISSN 1025-028X
Vaccimonitor vol.22 no.1 Ciudad de la Habana ene.-abr. 2013
ARTÍCULO ORIGINAL
Adjuvant activity of peanut, cottonseed and rice oils on cellular and humoral response
Actividad adyuvante de los aceites de maní, de semilla de algodón y de arroz en la respuesta celular y humoral
Erika Freitas,1* Ana Claudia Marinho,2 Deijanira Albuquerque,3 Leiliane Teles,2 Mércia Sindeaux,4 María Teresa Salles,5 Diana Célia Sousa,6 Margarida María Lima,4 María da Guia Silva,2 Dirce Fernandes2
1 Departamento de Biología, Universidade Federal do Ceará. CEP: 60455-900. Fortaleza, CE, Brasil.
2 Departamento de Bioquímica e Biología Molecular, Universidade Federal do Ceará. CEP: 60455-900. Fortaleza, CE, Brasil.
3 Departamento de Ciências Básicas em Saúde, Universidade Federal do Mato Grosso. CEP: 78000-900. Cuiabá, MT, Brasil.
4 Departamento de Patología e Medicina Legal, Universidade Federal do Ceará. CEP: 60430-970. Fortaleza, CE, Brasil.
5 Departamento de Química Orgânica, Universidade Federal do Ceará. CEP: 60455-900. Fortaleza, CE, Brasil.
6 Faculdade de Veterinária, Universidade Estadual do Ceará. CEP: 60740-002. Fortaleza, CE, Brasil.
email: erika.mota@ufc.br
* DSc, Professor. UFC.
ABSTRACT
The potentiality of the usage of vegetable oils such as soybean, corn, olive, sesame, murici seed, rapeseed, linseed, rice and cashew nuts as adjuvant of the humoral and cellular immune response has been recently shown. In the present work, besides of evaluating the adjuvant action of peanut, cottonseed and rice oils on humoral and cellular immune responses against ovalbumin (OVA) we also evaluated the protective immune response induced by Leishmania antigens. The peanut oil significantly increased the synthesis of anti-ovalbumin antibodies in the primary response, but it did not favor cellular response. Concerning mice immunized with L. amazonensis antigens emulsified with peanut oil exacerbated skin lesions and lymph node parasite load what suggests stimulation of the Th2 immune response and down regulation of Th1 response. The cottonseed oil was shown to have adjuvant effect to the humoral response, stimulating a secondary response and also favored the delayed-type hypersensitivity (DTH) response to OVA. The rice oil stimulated a strong DTH reaction to OVA and enhanced the synthesis of antibodies after the third dose. Mice immunized with L. amazonensis antigens emulsified with rice oil or cotton seed oil were protected from developing skin lesions and lymph node parasite load. These results emphasize the interest and importance of the vegetable oils as tools in different procedures of immunization and their differential role in relation to the other adjuvant under usage.
Key words: plant oils, adjuvants, cellular immune response, antibody response.
RESUMEN
Recientemente se ha demostrado el uso potencial de los aceites vegetales, tales como los de soya, maíz, oliva, ajonjolí, colza, nabina, linaza, arroz y anacardo, como adyuvantes de las respuestas inmune celular y humoral. En este trabajo, además de evaluar la acción adyuvante de los aceites de maní, de la semilla de algodón y del arroz en la respuesta inmune celular y humoral contra la ovalbúmina (OVA), también se evaluó la respuesta inmune protectora inducida por los antígenos de Leishmania. El aceite de maní incrementó significativamente la síntesis de los anticuerpos anti-ovalbúmina en la respuesta primaria, pero no favoreció la respuesta celular. Con respecto a los ratones inmunizados con los antígenos de L. amazonensis emulsionados con aceite de maní, este exacerbó las lesiones de la piel y la carga parasitaria en los nódulos linfáticos, lo que sugiere la estimulación de la respuesta inmune Th2 y la baja regulación de la respuesta Th1. El aceite de semilla de algodón mostró tener efecto adyuvante para la respuesta humoral, al estimular la respuesta secundaria y favorecer también la respuesta de hipersensibilidad demorada (DTH) a OVA. El aceite de arroz estimuló una fuerte reacción DTH a OVA y amplió la síntesis de los anticuerpos luego de la tercera dosis. Los ratones inmunizados con los antígenos de L. amazonensis emulsionados con aceite de arroz o de semilla de algodón recibieron protección contra el desarrollo de lesiones de la piel y la carga parasitaria de los nódulos linfáticos. Estos resultados destacan el interés y la importancia de los aceites vegetales como herramientas en los diferentes procedimientos de inmunización y su papel diferencial en comparación con otros adyuvantes en uso.
Palabras clave: aceites vegetales, adyuvantes, respuesta inmune celular, respuesta de anticuerpos.
INTRODUCCIÓN
Adjuvants are essential tools used to stimulate the immune response to purified antigens that have low immunogenicity. Traditionally complete Freund´s adjuvant (CFA) has been widely used in experimental models, but the severity of its side effects have impeded its use in vaccines for human beings and other animals (1). Therefore, the development of new adjuvants is stimulated by a number of factors, not only on the low immunogenicity of purified antigens as by its promising influence on DNA vaccines (2).
The judicious choice of adjuvants can influence certain aspects of antibody response like epitope specificity, affinity, class and/or isotype (3). Therefore, it becomes very important to study and develop new immunologic adjuvants with low toxicity, high efficiency and extensive resource (4).
Vegetable oils are such potential adjuvant sources. The most important advantage of vegetable oils is that they have minimal adverse side-effects since they are biodegradable compounds (5). Additionally, they are pure, safe, cheap and easily available. Initially, it was shown that vegetable oil based formulations are potent immunological adjuvants (5, 6). More recently, our group has shown that vegetable oils such as soybean, corn, olive, sesame, murici seed, rapeseed, linseed, rice and cashew nuts potently stimulated the humoral and cellular immune response (7-9).
In this work, we showed that emulsions made with soluble OVA plus peanut, cottonseed and rice oils were able to stimulate a strong antibody and DTH response to OVA. Additionally, peanut, cottonseed and rice oils regulated the immune response induced by Leishmania amazonensis in mice.
MATERIALS AND METHODS
Animals: The 8-week-old female Swiss mice and adult rats used in this work were provided by Central Animal House of the Federal University of Ceará, Brazil. The animals were kept in micro-isolators and all experiments have been conducted in accordance with the guideline for care and use of experimental animal of the National Counsel of Animal Experimentation Control (CONCEA-Brazil).
Antigens, plant oils and adjuvants: OVA (grade V) was obtained from Sigma Chemical Co., St. Louis, USA. Leishmania amazonensis (MHOM/Br/76/Leila) was used for both, infection and antigen preparation. Peanut (Arachis hypogeae L.) and cottonseed (Gossypium hirsutum L.) (Sigma) and commercial edible rice (Oryza sativa L.) oils were used after microbiological analysis and determination of the fatty acid content. Complete (CFA) and incomplete (IFA) Freund's adjuvants were purchased from Sigma Chemical Co., St. Louis, USA.
Determination of plant oils adjuvants activity on humoral immune response
Groups of 10 Swiss mice were immunized by subcutaneous (s.c.) injection with 10 mg OVA alone or emulsified to CFA, peanut (PN), cottonseed (CT) or rice (RC) oils following an immunization scheme with each dose administrated on days 0, 14 and 28. Non-immunized mice received equal volume of sterile physiological saline (SAL). The mice were bled from the orbital plexus on days 14, 28, 42 after the first injection. The sera were stored at -20 °C until usage. The total OVA-specific antibody levels were assayed by ELISA using sera from each animal of different groups, as previously described by Sartor et al (9). For each serum, the absorbance of six serial dilutions (1/400 and 1/39,062) were added and multiplied by 1000. These coded values were called ELISA scores (ELISA*), as described by Verdolin et al (10).
Evaluation of adjuvant activity of plant oils on the delayed-type hypersensitivity reaction to ovalbumin
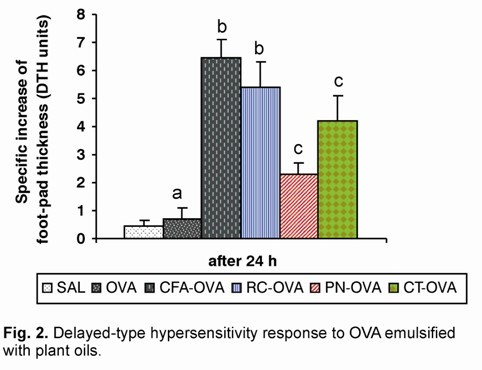
According Zimecki & Kruzel (11), mice were immunized by s.c. injection into tail base with 50 µg OVA (control) and with 50 µg OVA emulsified to CFA, PN, CT or RC oils. Non-immunized mice received equal volume of saline (SAL). After 7 days, the DTH reaction was elicited by s.c. injection of 50 µg OVA emulsified in IFA into one hind feet. The other foot-pad (control) was injected with IFA only. Foot-pad thickness was measured 24 h later with a dial-gauge caliper. The specific DTH reaction was calculated by subtracting the foot-pad thickness inject with IFA alone from the foot-pad thickness inject with OVA emulsified in IFA. All data are expressed in DTH units (1 unit = 0.1 mm) as mean values from twelve determinations ± S.E.
Evaluation of adjuvant activity of plant oils on the vaccine against leishmaniasis in a murine experimental model
The Leishmania antigens were prepared according to De Luca et al. (12) with modifications. In details, promastigotes were grown in Schneider's drosophila medium (Sigma Chemical Co., St. Louis, USA) supplemented with antibiotics [100 IU/mL penicillin G-potassium, 100 µg/mL streptomycin sulphate (Sigma)], 2 mM L-glutamine (Gibco BRL, Grand Island, USA), 5% sterile fetal bovine serum and 2% of sterile normal human urine for 5 days, as described by Teixeira et al. (13). The culture was then centrifuged at 4 °C for 20 min at 1350 x g. Promastigotes were washed three times, then suspended in SAL and incubated at 56 °C for 30 min. After the thermal treatment, the suspension of promastigotes was halved and the organisms in one half were disrupted by sonication during three times for 30 seconds (70 Hz). Then the two halves were mixed and the protein content was determined by the method of Lowry et al (14) with bovine seroalbumine (BSA) as the standard.
Swiss mice were subcutaneously (5/ group) immunized with the Leishmania antigens (360 µg/animal) emulsified or not with studied oils at 0 and 15th day. Non-immunized mice received equal volume of SAL. Two weeks after the second dose, the mice were subcutaneously challenged with 106 viable stationary-phase promastigotes in the left hind foot-pad.
Lesion sizes were measured with a dial gauge caliper (Mitutoyo, 0.01 mm sensitivity) and expressed as the difference in thickness (mm) between the infected foot-pad and counter lateral uninfected foot-pad. Measurements were initiated one week after the infection and continued for a period of 5 months. The animals were killed 5 months post-infection to assess parasitological loads in the draining lymph nodes, using the limiting dilution assay of Titus et al (15). The number of parasites, present in lymph nodes, was determined by minimum c2 analysis applied a Poisson distribution (16).
Determination of antioxidant potential of plant oils
The antioxidant activity of the peanut, cottonseed and rice oils was determined according to Owen et al. (17). Briefly, the relevant concentration range in methanol was added to 2 mL plastic centrifuge tubes in duplicate and the solvent was removed under a stream of nitrogen. The dried residue, resuspended in 1.0 mL of phosphate buffer saline (PBS), and 10.0 µL of a 1:5 dilution of xanthine oxidase in NH4SO4 (3.20 mol/L) was added to initiate the reaction. The tubes were incubated for 3 h until reaction completion at 37 °C. After incubation, 20 µL of the reaction mixture was analyzed by HPLC. The quantification of phenolic compounds and hydroxylation of hypoxanthine was carried out at 278 nm and the hydroxylation of salicylic acid at 325 nm.
Statistical analysis
The results were shown as the mean ± standard errors. Statistical analysis was performed using the Graph-Pad program PRISM 5.0. The data were compared by Mann-Whitney U-test. The significant differences between the means in the same group at different times the pared Student's t-test was used. Statistical significance was assigned for values of p < 0.05.
RESULT
Adjuvant activity of plant oils on the antibody response to ovalbumin
In order to evaluate the adjuvant effect of the peanut, cottonseed and rice oils on total anti-OVA antibodies, sera from mice immunized, as previously described were analyzed by ELISA. Data in figure 1 show that at the days 14th, 28th and 42nd after the first immunization, PN oil significantly increased the antibody levels against OVA compared to those obtained from mice immunized with OVA alone. Concerning to RC oil, it did not enhance total OVA-specific antibody levels on the days 14th and 28th (Fig. 1).
However, on the day 42nd RC oil significantly increased the total anti-OVA antibody response compared with those obtained from mice immunized with OVA alone. On the secondary response, the levels of total OVA-specific antibodies stimulated by CT oil were higher than those induced by OVA alone. According to the data, on the secondary response all plant oils studied stimulated the antibody levels to OVA. To assess the magnitude of adjuvant action of the peanut, cottonseed and rice oils, mice were immunized with OVA emulsified with CFA. On the 42nd day PN, RC and CT oils stimulated an OVA-specific humoral response similar to CFA (Fig. 1).
Stimulation of ovalbumin-induced delayed type hypersensitivity by plant oils
To investigate the adjuvant effect of the peanut, cottonseed and rice oils on cell-mediated response, DTH testing was performed. Results presented in Figure 2 show that RC-OVA stimulated an OVA-specific DTH response higher than OVA alone and similar to CFA-OVA. The PN and CT oils increased the OVA-specific DTH reaction higher than OVA alone and lower than that stimulated by CFA (Fig. 2).
Effect of plant oils on the immune response and protection against L. amazonensis
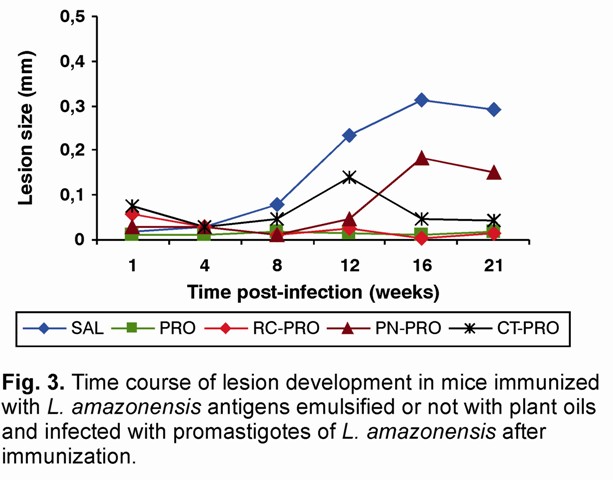
The adjuvant effect of the peanut, cottonseed and rice oils on the immune response was also evaluated using Leishmania antigens (PRO) emulsified or not with the studied oils. Non-immunized mice received equal volume of SAL. Figure 3 shows that, whereas non-immunized mice developed lesions from the 8th weeks after infection, mice subcutaneously immunized with Leishmania antigens emulsified with PN oil started developing lesions only from the 12th week post-infection. Furthermore, non-immunized mice as well as animals immunized with Leishmania antigens emulsified with PN oil developed similar and progressive lesions until the end of the experiment. Mice injected with Leishmania antigens emulsified with CT oil started developing smaller lesions from the 8th week post-infection that were controlled after the week 16th.
The vaccine of mice with Leishmania antigens alone as well as Leishmania antigens emulsified with RC oil protected them from infection with alive promastigotes (Fig. 3).
In order to obtain a better evaluation of the studied oils on the protection against Leishmania, the viable parasites were quantified in draining lymph nodes of immunized and non-immunized mice using a limiting dilution assay, as previously described (16).
Table 1 shows that lymph nodes from non-immunized mice presented a higher parasite load than lymph nodes from animals subcutaneously immunized with Leishmania antigens emulsified with PN oil. The vaccination of mice with Leishmania antigens alone as well as Leishmania antigens emulsified with RC oil presented negative parasite loads (Table 1).
Evaluation of antioxidant activity of plant oils
The antioxidant capacity of the peanut, cottonseed and rice oils is shown on the table 2. The compounds extracted from the PN, RC, and CT oils neutralized 40, 38, and 16% of reactive oxygen species (ROS) produced by the hypoxanthine/xanthine oxidase system, respectively.
DISCUSSION
In the current study, we investigated the adjuvant effect of cottonseed, peanut and rice oils on the humoral response and DTH reaction both induced by immunization of Swiss mice with OVA, and also to the capacity of protecting against murine experimental leishmaniasis after vaccination.
The peanut oil significantly elevated the synthesis of total anti-OVA antibodies already in the primary response, but it did not favor cellular response. Moreover, mice immunized with L. amazonensis antigens emulsified with peanut oil had exacerbated skin lesions and lymph node parasite load compared to the immunized control animals. These data suggest that peanut oil stimulated the Th2 immune response and down regulated Th1 response.
The cottonseed oil was a good adjuvant to the humoral response, stimulating a secondary response and being also capable to favor the DTH response to OVA.
Regarding to the rice oil it stimulated a strong DTH reaction to OVA and only enhanced the synthesis of antibodies after the third dose. Also, mice immunized with L. amazonensis antigens emulsified with rice oil or cottonseed oil were protected from developing skin lesions and lymph node parasite load as were the animals immunized with L. amazonensis antigens alone, suggesting that these oils did not alter the protective immune response induced by L. amazonensis antigens. In this work, the mechanism underlying the stimulation of the cellular immune response by rice oil was not described but we postulate that rice oil may act through its phenolic components, since it is the only one out of the studied oils that contains phenolic compounds, known by their ability to enhance the immune response (18).
Despite a significant humoral immune response stimulated by cottonseed, peanut and rice oils, at no time were any lesions noted at the site of injection of emulsions of these oils with antigens. This finding contrasts with the injection of IFA and CFA, which are known to induce severe lesions at their site of injections (1).
Although cottonseed, peanut and rice oils enhanced the antibody and cellular response to OVA, our results do not identify the underlying mechanism.
At least two different pathways are possible. First, cottonseed, peanut and rice oils may enhance the immune response by maintaining the antigen in the local of injection, since emulsions made of OVA with those oils were stable from 3 to 10 minutes. As it is well known, the sustained release of antigen over a period of time is necessary to stimulate the immune system and generate a high titer of antibody, perhaps though attraction and retention of professional antigen-presenting cells (6, 9). The second pathway may involve the regulation of the immune system by linoleic and oleic acids, as it has been previously shown (19, 20). Accordingly, peanut oil that contains 62.0% of oleic acid, it significantly stimulated the primary antibody response to OVA.
This result reinforce the data from literature showing that emulsions made of oleic acid plus diphiteric toxoid strongly increased the IgG levels to that toxoid (20). An important advantage of using vegetable oils as immunological adjuvants is that they have minimal adverse side-effects since they are easily metabolisable by the organism (5).
However, we hypothesize that the metabolizing property of vegetable oils may influence their ability to stimulate the primary immune response, as in our study cottonseed and rice oils did not stimulated the primary immune response to OVA. This potential disadvantage might be overcome by vegetable oils containing high levels of oleic acid such as peanut oil that stimulated the primary antibody response to OVA.
The variety of stimulated/regulated immune response by vegetable oil emulsions emphasizes the interest and importance of these oils as tools in different procedures of immunization and is a differential in relation to the other adjuvant under usage. We would conclude that these vegetable oils are potent adjuvants in immune response to OVA and apparently peanut oil down regulated the protective immune response induced by Leishmania antigens.
Acknowledgements
This work was supported by grants from CAPES, FUNCAP and CNPq.
REFERENCES
1. Gupta RK, Relyveld EH, Lindblad EB, Bizzini B, Ben-Efraim S, Gupta CK. Adjuvants- a balance between toxicity and adjuvanticity. Vaccine 1993;11:293-306.
2. Sesardic D, Dobbelaer R. European Union regulatory developments for new vaccine adjuvants and delivery systems. Vaccine 2004;22:2452-6.
3. Allison AC, Byars NE. Immunological adjuvants: Desirable properties and side-effects. Mol Immunol 1991;28:279-84.
4. Sun JH. Research headway of immunopotentiator. Helongjiang Ani Hus Vet Med 1998;2(4):40-2.
5. Eghafona NO. Immune response following cocktails of inactived measles vaccine and Arachis hypogea L. (groundnut) or Cocos nucifera L. oils adjuvant. Vaccine 1996;14:1703-8.
6. Audibert F, Chedid L. Increase of immune response by administration of metabolizable vegetable oil emulsions. CR Acad Sci Hebd Seances Acad Sci D 1975;280:1629-32.
7. Silva ACM. Estudo comparativo entre os efeitos imunoadjuvantes e inflamatórios dos óleos de oliva, milho e soja. [Thesis Doctoral]. Fortaleza, CE, Brazil: Departamento de Bioquímica e Biologia Molecular, Universidade Federal do Ceará; 2004.
8. Feijó DF. Efeito imunoadjuvante dos óleos de murici (Byrsonima crassifolia), sésamo (Sesamum indicum), linhaça (Linum usistatissimum), canola (Brassica sp.) e da amêndoa de caju (Anacardium occidentale). [Thesis Master]. Fortaleza, CE, Brazil: Departamento de Bioquímica e Biologia Molecular, Universidade Federal do Ceará; 2005.
9. Sartor ITM, Colodel EM, Albuquerque D. Adjuvant activity of rice oil on the immune response to ovalbumin. VacciMonitor 2011;20(2):1-5.
10. Verdolin BA, Ficker SM, Faria AMC, Vaz NM, Carvalho CR. Stabilization of serum antibody responses triggered by initial mucosal contact with the antigen independently of oral tolerance induction. Braz J Med Biol Res 2001;34:211-19.
11. Zimecki M, Kruzel ML. Systemic or local co-administration of lactoferrin with sensitizing dose of antigen enhances delayed type hypersensitivity in mice. Immunol Lett 2000;74:183-8.
12. De Luca PM, Mayrink W, Pinto JÁ, Coutinho SG, Santiago MA, Toledo VP et al. Randomized double-blind placebo-controlled trial to evaluate the immunogenicity of a candidate vaccine against american tegumentary leishmaniasis. Acta Trop 2001;80:251-60.
13. Teixeira CR, Cavassani KA, Gomes RB, Teixeira MJ, Roque-Barreira MC, Cavada BS et al. Potential of KM+ lectin in immunization against Leishmania amazonensis infection. Vaccine 2006; 24:3001-8.
14. Lowry OH, Rosebrough NJ, Faar AL. Protein measurement with the Folin phenol reagent. J Biol Chem 1951;193:265-75.
15. Titus RG, Ceredig R, Cerotinni JC, Louis JA. Therapeutic effect of anti-L3T4 monoclonal antibody GK 1.5 on cutaneous leishmaniasis in genetically susceptible BALB/c mice. J Immunol 1985;135:2108-14.
16. Taswell C. Limiting dilution assays for the determination of immunocompetent cell frequencies III. Validity tests for the Single-hit Poisson model. J Immunol Methods 1984;72(1):29-40.
17. Owen RW, Giacosa A, Hull WE, Haubner R, Spiegelhader B, Bartsch H. The antioxidant/anticancer potential of phenolic compounds isolated from olive oil. Eur J Cancer 2000;36:1235-47.
18. Bendich A. Symposium: antioxidants, immune response, and animal function. Physiological role of antioxidants in the immune system. Journal of Dairy Science 1993;76:2789-94.
19. Calder PC. Immunoregularory and anti-inflammatory effects of n-3 polyunsaturated fatty acids. Braz J Med Biol Res 1998;31:467-90.
20. Schörder U, Svenson SB. Nasal and parenteral immunizations with diphteria toxoid using monoglyceride/ fatty acid lipid suspensions as adjuvants. Vaccine 1999;17:2096-2103.
Recibido: Agosto de 2012
Aceptado: Septiembre de 2012

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}