










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkVaccimonitor
versión On-line ISSN 1025-0298
Vaccimonitor vol.26 no.1 Ciudad de la Habana ene.-abr. 2017
ARTÍCULO ORIGINAL
Activación del polisacárido capsular de Streptococcus pneumoniae serotipo 23F para la obtención de vacunas conjugadas
Activation of the capsular polysaccharide serotype 23F of Streptococcus pneumoniae to obtain conjugate vaccine
Janoi Chang-Calderón*, Yohanna Serrano-Rodríguez,** Raine Garrido-Arteaga, Jessy Pedroso-Fernández, Félix Cardoso-San Jorge, Laura Rodríguez-Noda, Darielys Santana-Mederos, Dagmar García-Rivero, Yury Valdés-Balbín, Vicente Vérez-Bencomo
Instituto Finlay de Vacunas. Ave. 27 No. 19805, La Lisa, La Habana, Cuba.
email: yserrano@finlay.edu.cu
* Doctor en Ciencias, Investigador Agregado.
** Maestra en Ciencias, Investigadora Agregada.
RESUMEN
En la actualidad, las vacunas conjugadas constituyen un gran hito en el desarrollo de fármacos que protegen contra las enfermedades infecciosas. Estas vacunas no solo disminuyen drásticamente la mortalidad y morbilidad de diferentes enfermedades causadas por bacterias en la población infantil; sino que también repercuten sobre la población no vacunada. Las vacunas conjugadas se basan en establecer una unión covalente entre un polisacárido y una proteína portadora para lo cual existen diferentes procedimientos químicos. Todos los procedimientos de conjugación requieren la presencia de grupos reactivos complementarios que muchas veces son generados en ambas macromoléculas. Este trabajo se enfoca en el estudio de la reacción de fragmentación y de la oxidación peryódica sobre el polisacárido capsular serotipo 23F de Streptococcus pneumoniae para su uso como antígeno vacunal. Se estableció la fragmentación del polisacárido mediante hidrólisis con ácido acético y trifluoroácetico. En el caso de la reacción de oxidación se encontró que la cantidad de moles de peryodato de sodio y la temperatura influyen de manera directamente proporcional sobre la generación de grupos carbonilos. Adicionalmente se demostró que el sustituyente glicerol-fosfatos presente en la estructura del serotipo 23F es relevante para conservar la antigenicidad. El procedimiento descrito permite obtener conjugados inmunogénicos a partir del polisacárido capsular de Streptococcus pneumoniae serotipo 23F en el modelo de conejos.
Palabras clave: Streptococcus pneumoniae, vacunas conjugadas, hidrolisis ácida, oxidación peryódica.
ABSTRACT
Nowadays conjugate vaccines are a major milestone in the development of drugs against infectious diseases. These vaccines drastically reduce mortality and morbidity from different diseases caused by bacteria in children; but also impact on non-vaccinated population. Conjugate vaccines are based on a covalent bond between a polysaccharide and a carrier protein for which there are different chemical procedures. All conjugate procedures require the presence of additional reactive groups that often are generated in both macromolecules. This work focus on the study of the fragmentation reaction and peryodic oxidation on the capsular polysaccharide serotype 23F Streptococcus pneumoniae for use as a vaccine antigen. It was possible to establish the fragmentation reaction of the polysaccharide by hydrolysis with acetic and trifluoroacetic acid. Directly proportional ratio was found between numbers of moles of sodium periodate and temperature on the oxidation reactions. In addition the glycerol-phosphate substituent was found as important motif to preserve the antigenicity. The procedure allows immunogenic conjugate from capsular polysaccharide serotype 23F of Streptococcus pneumoniae in rabbit models.
Keywords: Streptococcus pneumoniae, conjugate vaccines, peryodic oxidation, acid hydrolysis.
INTRODUCCIÓN
Actualmente las infecciones causadas por Streptococcus pneumoniae tienen un gran impacto sobre la morbilidad y mortalidad infantil, problemática que es enfrentada con gran efectividad mediante la introducción de las vacunas conjugadas (1,2). El éxito de estas vacunas se sustenta en lograr que el sistema inmune inmaduro de los niños menores de 5 años responda ante los polisacáridos (Ps), antígenos T-independientes, como si fueran antígenos T-dependiente. Esto se logra al establecer un enlace covalente entre el polisacárido bacteriano y una proteína portadora (antígenos T-dependientes).
Existen diversos métodos químicos que describen la formación de esta unión covalente entre ambas macromoléculas, pero en todos los casos es necesario la presencia de grupos reactivos complementarios (3-5). Las proteínas al estar formadas por aminoácidos presentan una gran diversidad de grupos reactivos como: grupos aminos, ácidos carboxilos, tioles, guanidinio, entre otros. Los polisacáridos, por otro lado, presentan menor diversidad de grupos reactivos que la presentada en las proteínas. Debido a esto, en muchos casos se introduce una etapa para la activación de los polisacáridos en los procedimientos de conjugación. Esta etapa de transformación química tiene por objetivo generar grupos más reactivos en el Ps.
Una de estas reacciones es la disminución de la talla molecular que permite el uso de los extremos reductores para luego introducir un brazo espaciador (6). Adicionalmente, la disminución de la talla también brinda ventajas tecnológicas en el trabajo con los Ps como son: un producto con distribución de talla mejor definida, evitar los problemas de solubilidad y viscosidad de los polisacáridos capsulares nativos, y facilitar los procesos de purificación del producto final (7,8). La oxidación con peryodato de sodio es un procedimiento muy utilizado para la activación de los polisacáridos. En esta reacción se forman grupos aldehídos, mediante la oxidación de dos grupos hidroxilos vecinales que son los más abundantes en los polisacáridos capsulares (PsC), con la consiguiente ruptura del enlace carbono-carbono. Los polisacáridos capsulares están formados por unidades repetitivas que pueden contener uno o varios monosacáridos y además pueden presentar ramificaciones (9).
La reactividad frente a la oxidación con peryodato en los polisacáridos es primeramente sobre los alcoholes primarios, luego los hidroxilos vecinales en configuración cis y como último los dioles de configuración trans. Esta susceptibilidad es independientemente a la ubicación de estos en la estructura del polisacárido, como parte de una ramificación o dentro de la cadena lineal (10). Por ello, el uso de este procedimiento para la obtención de antígenos vacunales requiere de un control de las condiciones de la reacción para evitar afectaciones significativas de la estructura de los polisacáridos y con ello la pérdida de información inmunológica del polisacárido. El presente trabajo tiene como objetivo establecer un procedimiento para la obtención de polisacáridos fragmentados y activados con grupos aldehídos a partir del polisacárido capsular de Streptococcus pneumoniae serotipo 23F. El procedimiento se enfoca en el estudio de la reacción de disminución de la talla molecular del polisacárido capsular mediante la hidrólisis ácida y la formación de grupos aldehídos mediante la oxidación con peryodato de sodio sin afectar la antigenicidad del Ps.
MATERIALES Y MÉTODOS
Materiales: La proteína toxoide tetánico (TT) lote 6017 suministrada por el Instituto Finlay de Vacunas se obtiene como ingrediente farmacéutico activo en el proceso productivo de la vacuna triple bacteriana DPT®. En el caso del PsC 23F lote 8001, se obtuvo en el Departamento de Desarrollo Farmacéutico del Instituto, cumpliendo con los requisitos recomendados por la Organización Mundial de la Salud (OMS) para la producción de vacunas (11).
Se utilizaron materiales de referencia como el PsC 23F purificado proveniente de la American Type Culture Collection (ATCC) y el suero estándar internacional generado en conejos, específico contra el serogrupo 23 (23A, 23B y 23F), suministrado por Statens Serum Institute, Copenhagen.
Técnicas analíticas: La cuantificación de Ps para determinar los rendimientos de las reacciones se realizó mediante la técnica descrita por Brückner (12) y los grupos carbonilos generados se determinaron a través del método informado por Porro y cols. (13). En el caso del contenido de proteína en los conjugados se empleó la técnica de cuantificación de proteínas en disolución, descrita por Lowry y cols. (14).
La evaluación de la identidad de los PsC se realizó mediante la resonancia magnética nuclear protónica (RMN 1H) en un equipo espectrómetro Bruker/Avance DPX 250. El procesamiento de la caída libre de inducción se realizó a través del software MestReNova. Las muestras se prepararon tomando entre 5-10 mg del producto seco y se disolvieron en 0,6 mL de agua deuterada (D2O, Merck >99.8 %) y se secaron en una liofilizadora (Christ Beta 1-8 LD). Este procedimiento se realizó dos veces antes de disolverlo en agua deuterada para adquirir el espectro, los cuales se adquirieron a una temperatura de 52 ºC. La determinación de la constante de distribución de la talla molecular (KD) se realizó por cromatografía líquida de alta resolución en columna de exclusión molecular TSK 5000 PW con NaCl 0,9 % como fase móvil y detección por índice de refracción para el PsC, polisacárido fragmentado (PsFg-23F) y el polisacárido oxidado (PsOX-23F) o detección ultravioleta (206 nm) para los conjugados (23F-TT). El análisis de los datos se realizó a través del software ClarityChrom.
Preparación y caracterización de los polisacáridos y conjugados de 23F
Disminución de la talla molecular del PsC 23F: A una disolución que contiene 10 mg de PsC 23F en 2 mL de agua destilada, se le adicionó igual volumen de la disolución del ácido en estudio, ácido acético (HAc) o ácido trifluoroacético (TFA), al doble de la concentración definida. La mezcla se mantuvo con agitación a 70 ºC entre 1–3 h. Transcurrido este tiempo la reacción se neutralizó utilizando una disolución de hidróxido de sodio 0,1 M y la mezcla se purificó por membranas de celulosa regenerada de 30, 10 y 1 kDa de porosidad. La eficiencia de la reacción se determinó de acuerdo al contenido de carbohidrato recobrado en las fracciones 10-30 kDa.
Oxidación con peryodato de sodio del PsFg-23F: Se preparó una disolución de Ps fragmentado en disolución tampón fosfato salina pH 7,0 de manera que se obtuvo una concentración de Ps de 4, 10 o 20 mg/mL, según lo definido en la aleatorización del diseño experimental. Luego, se adicionó igual volumen de disolución de NaIO4 que contiene los moles equivalentes establecidos en el diseño. La mezcla de reacción se mantuvo con agitación moderada, protegida de la luz a la temperatura correspondiente. Transcurridas las 3 horas de reacción se añadieron dos equivalentes de glicerol por mol de NaIO4 adicionado. Se conservaron las condiciones de reacción durante 30 min. La mezcla de reacción se purificó mediante ultrafiltración a través de membrana de celulosa regenerada de tamaño de poro 10 kDa.
Conjugación del PsOx-23F a TT mediante aminación–reductiva: La reacción de conjugación se realizó según el procedimiento descrito (15). Solamente se modificó la relación de masa de Ps y proteína que resultó de 1:1 y la concentración de la reacción fue de 10 mg/mL.
Evaluación de la inmunogenicidad: El protocolo en animales, estuvo sujeto al cumplimiento de las normas éticas en la experimentación animal y se aprobó por el Comité de Ética y el Departamento de Calidad de la institución. Se utilizaron conejos blancos de Nueva Zelanda, hembras, de 1,5-1,8 kg de peso (10-12 semanas de edad), procedentes del Centro Nacional para la Producción de Animales de Laboratorio (CENPALAB). Se crearon grupos de 5 animales que se inmunizaron con una dosis de 5 μg de Ps contenido en el conjugado o 25 μg de PsC 23F. Además, se incluyó un grupo control al que se le administró suero fisiológico. Las inmunizaciones se realizaron a los 0, 14 y 28 días por vía subcutánea y las extracciones de sangre se realizaron 7 días después de cada inmunización.
Evaluación de memoria inmunológica: A los 120 días, cuando la respuesta de IgG contra el PsC 23F disminuyó, se aplicó una dosis de refuerzo de 25 μg de PsC 23F. Se evaluó la inducción de IgG siete días posteriores a la inmunización. Todos los sueros obtenidos se conservaron a -20 °C hasta su evaluación.
Ensayos de ELISAsEvaluación del título de IgG contra el PsC 23F: Para evaluar la respuesta de anticuerpos contra el PsC 23F, se siguió un ELISA similar al recomendado por la OMS (16) aunque no fue posible utilizar el sistema de detección por fosfatasa alcalina, sustituyéndose por un conjugado anti conejo-IgG-HRP. Los pasos del ELISA se detallan a continuación: a las placas de 96 pocillos de poliestireno (Maxisorp) recubiertas con 10 µg/mL de PsC 23F (50 µL/pocillo) y bloqueadas con 150 µL/pocillo de disolución de albúmina de suero bovino al 1 %, se les adicionaron los sueros extraídos luego de cada inmunización en diluciones dobles seriadas comenzando por 1/100 en disolución tamponada fosfato salina pH 7,2 más BSA 1 %, EDTA 0,01 M, Tween 20 0,3 %, incubándose durante 90 min a temperatura ambiente. Se adicionó el conjugado anti conejo-IgG peroxidasa (dilución 1/10000) y se incubó durante 90 min a temperatura ambiente. El revelado se realizó adicionando 100 µL/pocillo de orto-fenilendiamina 0,5 mg/mL en disolución citrato (Na2HPO4 52 mM y ácido cítrico 25 mM, pH 5,6) y se mantuvo la placa en oscuridad durante 20 min. Se detuvo la reacción adicionando HCl 3M, se leyó la densidad óptica (DO) a una longitud de onda de 492 nm en un lector de ELISA. Al final de cada incubación se realizó el lavado de las placas con disolución de lavado (disolución tampón fosfato salina pH 7,2, Tween 20 0,05 %).
El cálculo del título de anticuerpos IgG se realizó mediante un análisis de regresión entre los valores de DO y el logaritmo en base 10 del recíproco de la dilución del suero. Para la selección de los animales respondedores, se consideró como valor de corte para la titulación, el doble del valor de absorbancia del suero preinmune (t=0) diluido 1/100, por tanto, el título de anticuerpos se expresó como el log10 de la dilución del suero que dio un valor de DO igual al doble del valor obtenido para el suero preinmune (t=0).
Evaluación de la conservación de la antigenicidad: Para la evaluación de la antigenicidad, se recubrieron las placas en las mismas condiciones que para el ELISA de titulación. Se adicionó el suero de referencia internacional contra el serogrupo 23 (1/6400), disuelto en solución tampón de dilución, previamente incubado durante toda la noche a 4 ºC, con diluciones seriadas 1/10 del PsC en un intervalo de concentraciones de 0,005 µg/mL hasta 500 µg/mL.
El resto del procedimiento se realizó de forma similar a lo descrito para la titulación contra el PsC. Los resultados se muestran como porcentaje de inhibición y se calculó según la siguiente fórmula: 1-[DO (sin inhibidor)-DO (con inhibidor) / DO (sin inhibidor)] X 100.
Análisis Estadístico
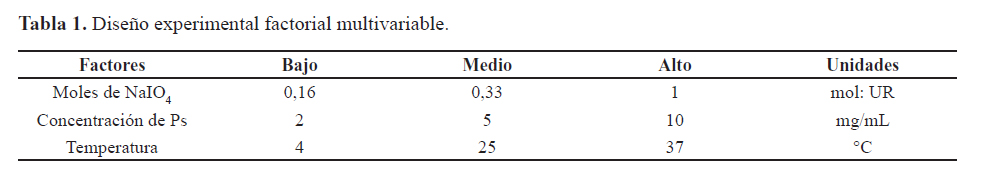
La influencia de la concentración de PsFg-23F, los equivalentes de NaIO4 y la temperatura en la reacción de oxidación con NaIO4 se realizó a través de un diseño experimental factorial multinivel 3^3 (tabla 1). Se escogieron como variables respuestas; la cantidad de grupos carbonilos introducidos en la estructura del PsFg-23F, expresados como mol de grupo carbonilo por unidad repetitiva (UR), y el rendimiento de la reacción a través del recobrado de PsFg-23F. El análisis estadístico se realizó a través del programa estadístico de computación Statgraphics Centurion XV (StatPoint Inc. 1982-2007), se consideró significación estadística para p = 0,05. Las diferencias en la respuesta de anticuerpos IgG anti-PsC 23F entre los distintos grupos experimentales se realizó mediante la aplicación de la prueba de Kruskal Wallis (no paramétrico) y la prueba de Dunn a posteriori. Las diferencias se consideraron significativas para p = 0,05. El procesamiento estadístico de los datos de todos los experimentos se realizó con el programa GraphPad Prism, versión 4.00.
RESULTADOS Y DISCUSIÓN
Caracterización del PsC 23F mediante RMN 1H
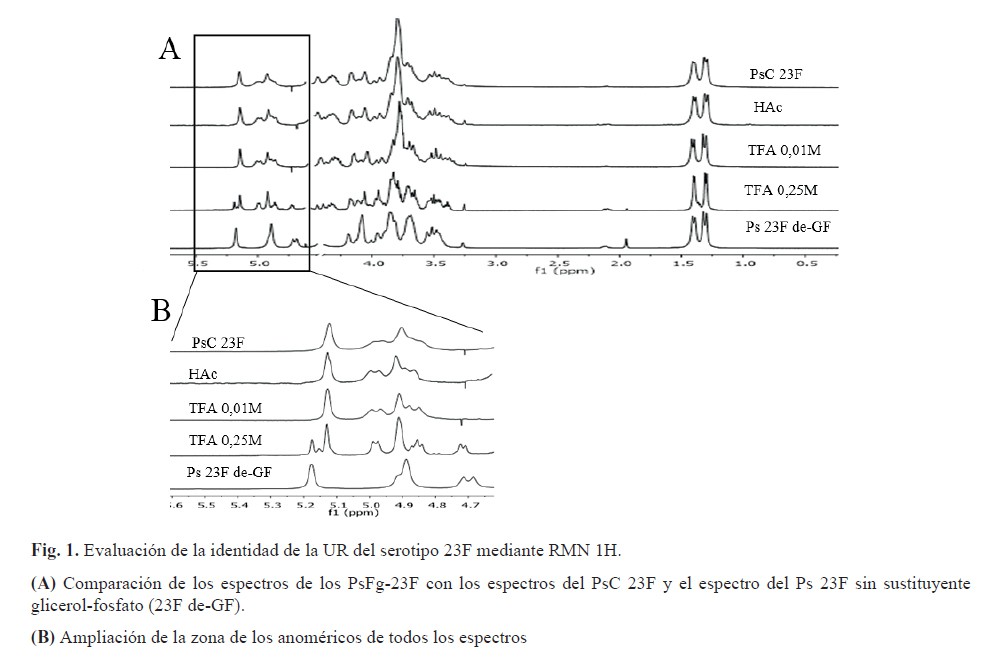
La identidad del PsC de Streptococcus pneumoniae serotipo 23F se realizó mediante RMN 1H. Se apreciaron las señales correspondientes a los grupos metilos de la a-L-ramnosa y ß-L-ramnosa a 1,26 y 1,36 ppm respectivamente. Además, se identificaron las señales correspondientes a los cuatro anoméricos a 4,83; 4,85; 4,93 y 5,1 ppm lo cual coincide con lo reportado en la caracterización de la unidad repetitiva del serotipo 23F por RMN 1H y con el espectro registrado para el PsC 23F ATCC de referencia. Adicionalmente se obtuvo un Ps de 23F sin sustituyente glicerol-fosfato según lo descrito por James Richards en 1988 (17) para utilizarlo como referencia de afectación del sustituyente glicerol-fosfato en los ensayos de identidad por RMN 1H y de antigenicidad por ELISA.
Obtención del PsFg-23F mediante hidrólisis ácida
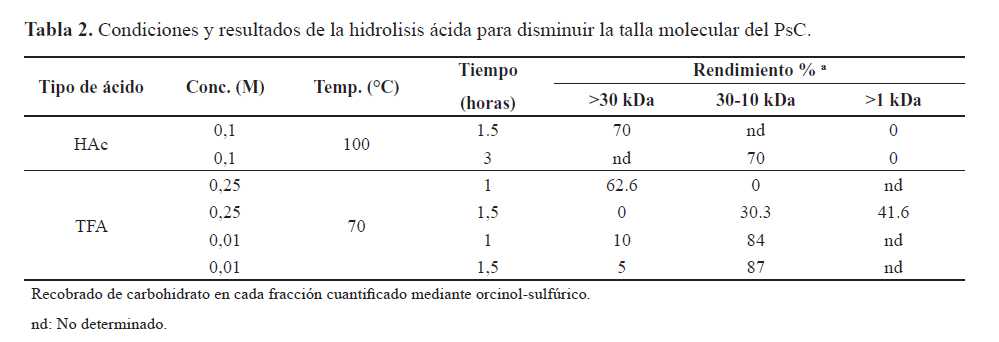
La fragmentación de los PsC se implementa mediante métodos físicos, mecánicos y químicos. La disminución de la talla del PsC 23F se estudió mediante hidrólisis ácida y se evaluaron dos tipos de ácidos en diferentes condiciones. Los resultados muestran que tanto con HAc como TFA se obtienen PsFg-23F con tallas entre 10-30 kDa, aunque los mejores rendimientos (84 %) se alcanzaron con TFA (tabla 2). Al ser el TFA un ácido más fuerte se requiere de un menor tiempo, así como de una menor temperatura de reacción. Estos resultados son comparables con los reportados para la reducción de la talla molecular de polisacáridos de neumococo por hidrólisis ácida en cuanto a la eficiencia del TFA (19).
Un requisito muy importante al modificar los PsC para utilizarlos como antígenos de vacunas conjugadas es lograr conservar los epítopes B. La unidad repetitiva del PsC 23F presenta dos ramificaciones, un residuo de L-ramnosa que debe ser conservado ya que es un determinante antigénico, lo cual se demostró con sueros de conejos, de caballos y humanos (20). La otra ramificación es un sustituyente glicerol-fosfato de la cual no encontramos información sobre su importancia inmunológica. Ambas ramificaciones pueden sufrir hidrólisis con TFA (21) y de ser eliminadas pueden causar afectación de la antigenicidad del PsFg-23F.
Los espectros de RMN 1H de los PsFg-23F obtenidos con ambos ácidos mostraron que todos conservaron la identidad de la unidad repetitiva al compararse con el espectro del PsC 23F, excepto cuando se utilizó TFA 0,25 M (figura 1A). En el espectro correspondiente se aprecia una señal en δ 5,12 ppm característica de la ausencia del sustituyente glicerol-fosfato (figura1B) y que se corresponde con la α-L-ramnosa según lo reportado por Richards en 1988 (17).
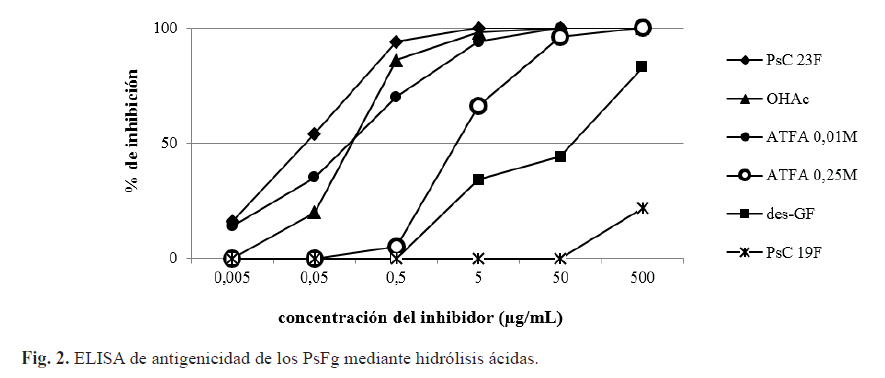
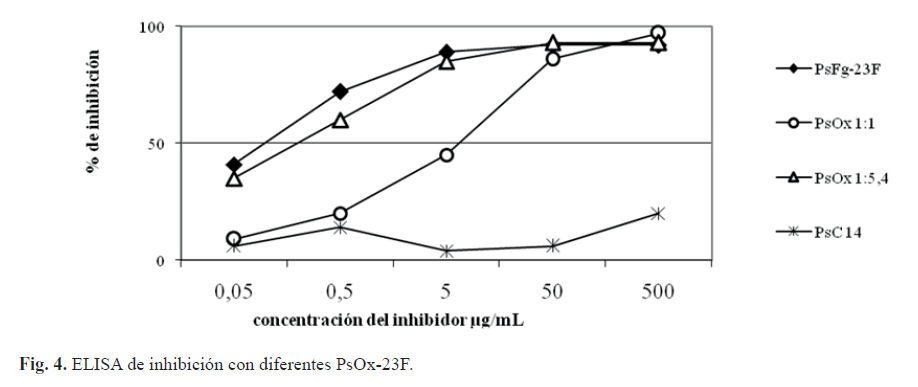
La antigenicidad se evaluó mediante un ELISA de inhibición, para lo cual se utilizó un suero de referencia específico contra el serogrupo 23. Las curvas de inhibición registradas con los diferentes fragmentos presentaron similitud a la obtenida para el PsC 23F, excepto para los fragmentos con afectaciones del sustituyente glicerol-fosfato (figura 2).
El PsC 23F sin sustituyente glicerol-fosfato obtenido como referencia del estudio no es capaz de inhibir al suero específico en un 100 %, y el PsFg-23F (0,25M de ácido trifluoroacético) con afectación parcial del sustituyente glicerol-fosfato necesitó más de 10 veces la concentración para alcanzar el 50% de inhibición en comparación con el resto de los derivados evaluados (figura 2). En nuestro caso, todos los fragmentos obtenidos se encuentran en el mismo intervalo de tallas moleculares (KD), luego, la disminución de la antigenicidad está directamente relacionada con la afectación del sustituyente glicerol-fosfato. Este resultado indica que el glicerol-fosfato juega un rol fundamental en la antigenicidad del serotipo 23F, lo cual ha sido observado para el serotipo 18C de neumococo donde este sustituyente es un elemento importante en el reconocimiento por parte del sistema inmune (18).
Obtención del PsOx-23F mediante la oxidación con NaIO4
La unidad repetitiva del PsC 23F presenta varios sitios posibles de ser oxidados, algunos de los cuales se localizan en el residuo de L-ramnosa de la cadena lateral, descrita como determinante antigénico (19). Con el objetivo de tener mayor conocimiento de la reacción de oxidación con NaIO4 se realizó un diseño experimental multifactorial 3^3. El análisis estadístico arrojó que los parámetros equivalentes de NaIO4 (p=0,000 para ambas variables) y la temperatura (p=0,0036 para nivel de oxidación y p=0,0001 para el rendimiento) influyen de manera significativa sobre el nivel de oxidación y los rendimientos obtenidos, aunque los equivalentes de NaIO4 presentan mayor significación. De manera inesperada los resultados mostraron que la variación de la concentración de Ps en la reacción no influyó en ninguna de las variables evaluadas. Tanto los equivalentes de NaIO4 como la temperatura tienen una relación directa con la generación de grupos aldehído (p= 0,0018).
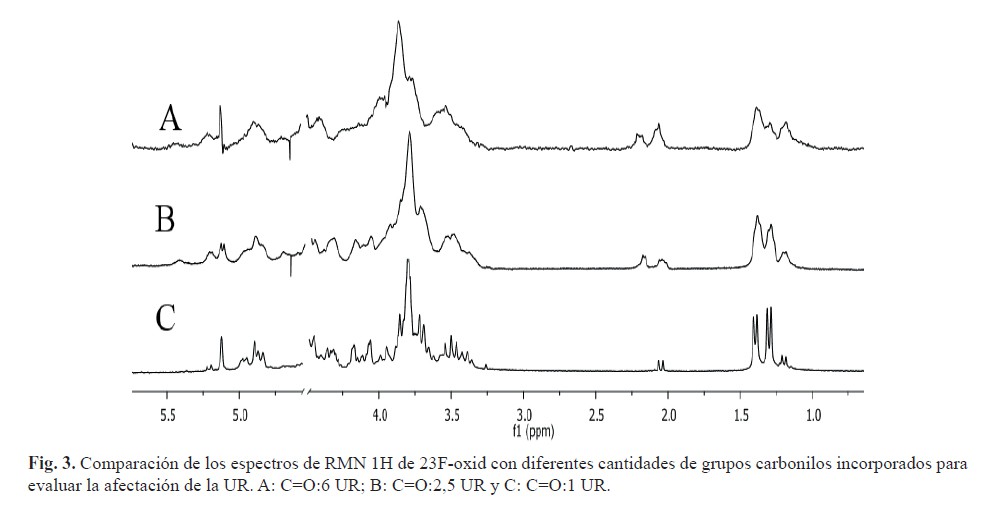
Además, se realizó una evaluación de la conservación de la identidad y la antigenicidad sobre los PsOx-23F. Al comparar los espectros de RMN 1H de PsOx-23F con diferentes contenidos de grupos carbonilos se observó afectación de la identidad de la UR del PsC 23F (figura 3). A medida que aumentó el número de grupos carbonilos generados, los espectros de RMN 1H presentaron menor resolución. El análisis espectroscópico muestra una disminución de la intensidad relativa de la señal correspondiente al grupo metilo de la α-L-ramnosa (δ 1,26 ppm) y un aumento de las señales a δ 1,20; 2,04; 2,14 y 5,20 ppm, respectivamente. Se puede relacionar la señal de 1,20 ppm con el grupo metilo de la ramnosa lateral oxidada y la señal de δ 5,20 ppm con el carbono anomérico de este sustituyente. El conjunto de modificaciones apreciadas evidencian que desde la generación de un carbonilo cada 2,5 UR existe afectación de la identidad de la UR en el PsOx-23F. Por otro lado, a medida que se aumenta la oxidación sobre el Ps 23F ocurre afectación de la antigenicidad, la cual es marcada en el máximo nivel de activación evaluado (C = O:1 UR) (figura 4), por lo que se aprecia una correspondencia con los resultados obtenidos en el análisis espectroscópico. Lo anterior indica que la disminución de la antigenicidad se debe a la oxidación excesiva del residuo de α-L-ramnosa, donde se encuentran los sitios más susceptibles a la oxidación (20) y al mismo tiempo es un determinante antigénico (21).
Se realizaron 5 réplicas de las condiciones de oxidación para obtener un grupo carbonilo entre 5-7 UR. Se obtuvo un rendimiento promedio de 76,4 %; un nivel de oxidación de un grupo carbonilo cada 6,2 y una KD promedio de 0,55. El espectro del PsOx-23F mostró todas las señales características del PsC 23F natural. Además, se observan los cuatros protones anoméricos, con intensidades relativas similares a las del PsFg-23F, que indica que se conservó la cadena lateral.
Por otro lado no se observaron señales relacionadas con la afectación del sustituyente glicerol-fosfato, el cual es imprescindible para mantener la antigenicidad del PsC 23F. No obstante, se apreciaron nuevas señales a δ 1,20; 2,04; 5,20; 6,59 y 9,22 ppm, aunque todas con baja intensidad relativa. Estos corrimientos químicos están relacionados con los cambios estructurales que provocaron la generación de los grupos carbonilos en la estructura del PsOx-23F como se describió anteriormente.
Conjugación del PsOx-23F a la proteína portadora toxoide tetánico
Se seleccionó un PsOx-23F con un carbonilo cada 6 UR y se conjugó a la proteína portadora TT mediante la reacción de aminación reductiva para conocer si estos podían inducir una respuesta inmune específica contra el PsC 23F. Las características de los conjugados obtenidos fueron: un recobrado superior al 48 %, una relación carbohidrato: proteína en el intervalo de 1,1:1 hasta 1:1,2 y un porcentaje de proteína no conjugada promedio de 12,5 %, determinada en columna SUPEROSA 12.
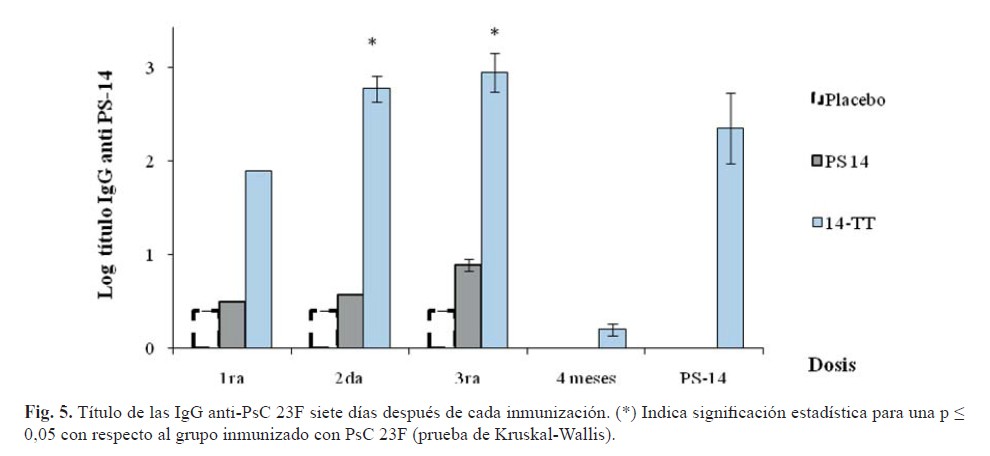
Todos los valores obtenidos de la relación carbohidrato: proteína se encuentran incluidos en el intervalo recomendado por la OMS para la obtención de vacunas conjugadas de neumococo (11). Además, el análisis en cromatografía líquida de alta resolución en columna de exclusión molecular arrojó una KD de 0,33 en columna TSK 5000. Los títulos de IgG contra el PsC 23F inducido por el conjugado fueron superiores con significación estadística a los inducidos por el grupo inmunizado con PsC 23F luego de la segunda y tercera dosis (figura 5). Por otro lado, el conjugado incrementó la generación de IgG después de cada inmunización, aunque solo fue estadísticamente significativa luego de la segunda dosis.
Siete días posteriores a la inmunización con polisacárido capsular, se observó un incremento significativo de los títulos de IgG, solo en el grupo previamente inmunizado con conjugado. Esto indica que la respuesta inmune generada por los conjugados fue capaz de inducir memoria inmunológica, que supone la presencia de poblaciones específicas al PsC de células B de memoria, lo cual es característico de antígenos de tipo T dependientes.
Los resultados experimentales permiten concluir que se puede utilizar la hidrólisis ácida para la fragmentación del PsC 23F de manera que se conserve la identidad de la UR y la antigenicidad del Ps. El sustituyente glicerol-fosfato presente en el PsC 23F es importante para el reconocimiento por los anticuerpos, por lo cual debe ser conservado. La oxidación peryódica sobre el polisacárido 23F debe ser controlada para evitar la disminución de la antigenicidad por la afectación del residuo de L-ramnosa. Los conjugados obtenidos por esta vía generan una respuesta T-dependiente específica contra el polisacárido capsular serotipo 23F.
REFERENCIAS
1. Simonsen L, Taylor RJ, Young-Xu Y, Haber M, May L, Klugman KP. Impact of pneumococcal conjugate vaccination of infants on pneumonia and influenza hospitalization and mortality in all age groups in the United States. MBio 2011;2:e00309-10.
2. Pilishvili T, Lexau C, Farley M, Hadler J, Harrison L, Bennett N, et al. Sustained reductions in invasive pneumococcal disease in the era of conjugate vaccine. J Infect Dis 2010;201:32-41
3. Lees A, Nelson B, Mond J. Activation of soluble polysaccharides with 1-cyano-4-dimethylaminopyridinium tetrafluroborate for use in protein–polysaccharide conjugate vaccines and immunological reagents. Vaccine 1996;14:190–8.
4. Gildersleeve J, Oyelaran O, Simpson J, Allred B. Improved Procedure for Direct Coupling of Carbohydrates to Proteins via Reductive Amination. Bioconjug Chem 2008;19(7):1485-90.
5. Paterson G, DunKan J. Recent advances in the field of Salmonella Typhi vaccines. Human Vaccine 2010;6(5):379-84.
6. Qingrui H, Dongxia L, Aijun K, Wenqi A, Bei F, Xiaowei M, et al. PEG as a spacer arm markedly increases the immunogenicity of meningococcal group Y polysaccharide conjugate vaccine. Journal of Controlled Release 2013;172:382-9.
7. Pawlowski A, Kallenius G, Svenson S. Preparation of pneumococcal capsular polysaccharide-protein conjugate vaccines utilizing new fragmentation and conjugation technologies. Vaccine 2000;18(18):1873-85.
8. Soubal J, Peña L, Santana D, Valdés Y, García D, Pedroso J, et al. Procedure for the conjugation of the Streptococcus pneumoniae serotype 6B capsular polysaccharide to the tetanus toxoid. Biotecnología Aplicada 2013;30:208-15.
9. Kamerling J. Pneumococcal polysaccharides: A chemical view. En: Tomasz A, editor. Streptococcus pneumoniae: molecular biology and mechanisms of disease. New York: Mary Ann Lierbert, Inc.; 2000. p.85–93.
10. John S, Laskowich E, Michon F, Kaiser R, Arumugham R. Monitoring activation sites on polysaccharides by GC–MS. Anal Biochem 2006;358(1):136-42.
11. World Health Organization. Recommendations for the production and control of pneumococcal conjugate vaccine. Technical Report Series No 927. Geneva: WHO; 2005.
12. Bruckner J. Estimation of monosaccharides by the orcinol-sulfuric acid reaction. Biochemical Journal 1995;60:200-5.
13. Porro M, Vitti S, Antoni G, Neri P. Modifications of Park-Johnson ferricyanide submicromethod for the assay of reducing groups in carbohydrates. Anal Biochem. 1981;118:301-6.
14. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951;193:265-75.
15. Chang J, SerranoY, Garrido R, Pedroso J, Cardoso F, García D, et al. Caracterización de conjugados inmunogénicos de polisacárido capsular Streptococcus pneumoniae serotipo 14. VacciMonitor 2013;22(1):15-21.
16. World Health Organization. Training Manual for Enzyme-Linked Immunosorbent Assay for the Quantitation of Streptococcus pneumoniae Serotype-Specific IgG (Pn Ps ELISA). London: World Health Organization Pneumococcal Serology Reference Laboratories; 2016. Disponible en: https://www.vaccine.uab.edu/ELISA%20protocol.pdf.
17. Richards J, Perry M. Structure of the specific capsular polysaccharide of Streptococcus pneumoniae type 23F (American type 23). Biochem Cell Biol 1988;66:758-71.
18. Chang J, Serrano Y, Garrido R, Rodríguez L, Pedroso J, Cardoso F, et al. Relevance of O-Acetyl and phosphoglycerol groups for the antigenicity of Streptococcus pneumoniae serotype 18C capsular polysaccharide. Vaccine 2012;30:7090-6.
19. Park S, Nahm M. L-rhamnose is often an important part of immunodominant epitope for pneumococcal serotype 23F polysaccharide antibodies in human sera immunized with PPV23. PLoSOne 2013;8:e83810. doi: 10.1371/journal.pone.0083810.
20. Charlotte C, Yu I, Manam V, Hepler R, PHennessey J. Carbohydrate composition analysis of bacterial polysaccharides: optimized acid hydrolysis conditions for HPAEC-PAD analysis. Anal Biochem 1992;201:343-9.
21. Kim J, Laskowich E, Michon F, Kaiser R, Arumugham R. Monitoring activation sites on polysaccharides by GC-MS. Anal Biochem. 2006;358:136-42.
Recibido: Noviembre de 2016 Aceptado: Enero de 2017


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}