










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkVaccimonitor
versión On-line ISSN 1025-0298
Vaccimonitor vol.26 no.2 Ciudad de la Habana may.-ago. 2017
ARTÍCULO ORIGINAL
Heterologous protein production in the moss Physcomitrella patens
Producción de proteínas heterólogas en el musgo Physcomitrella patens
Evelien Cools 1, Patrick Van Dijck 1,2, Nelson Avonce 1,2,3*
1 Department of Molecular microbiology, VIB, KasteelparkArenberg 31, 3001. Heverlee Belgium.
2 Laboratory of Molecular Cell Biology, Institute of Botany and Microbiology, K.U. Leuven, KasteelparkArenberg 31, B-3001. Heverlee Belgiuma.
3 Centro de Investigación en Dinámica Celular, Instituto de Investigación en Ciencias Básicas y Aplicadas, Universidad Autónoma del Estado de Morelos, México.
email: nelson.avonce@uaem.mx
* Profesor Investigador.
ABSTRACT
Over the last decades plants have been used for the heterologous production of pharmaceuticals, industrial enzymes and edible vaccines. The moss Physcomitrella patens is considered as an experimental model of choice for basic molecular, cytological and developmental questions in plant biology, as well as an outstanding plant model system for heterologous protein production. Here we use P. patens to produce osmotin, a tobacco protein with fungicidal properties. We have generated a transgenic plant able to synthesize and secrete a biologically active osmotin protein.
Keywords: moss, bioreactor, recombinant protein, osmotin.
RESUMEN
Durante las últimas décadas las plantas han sido utilizadas para la producción heteróloga de farmacéuticos, enzimas de uso industrial y vacunas. El musgo Physcomitrella patens es considerado como un modelo de experimentación de elección para abordar preguntas en las áreas de biología molecular, citología y de biología del desarrollo en plantas; así como un excelente sistema modelo para la producción de proteínas heterólogas. En este trabajo se utilizó P. patens para la producción de osmotina, una proteína de tabaco con propiedades fungicidas. Se generó una planta transgénica capaz de sintetizar y secretar osmotina biológicamente activa.
Palabras clave: musgo, bioreactor, proteína recombinante, osmotina.
INTRODUCTION
The search for low cost methods to produce pharmaceutical proteins, vaccines and other industrial-related enzymes has been one of the major goals for pharmaceutical industry during the last years. Several methods such as microbial cell cultures, animal cell cultures, plant tissue cultures, transgenic animals and plants have been explored and are used today. Plants are now considered to be an equally viable and competitive large-scale production platform for the production of pharmaceutical proteins.
Major factors in favor of plant-based systems for the production of animal-derived proteins are its potential for large-scale production, low-cost production, low risk of contamination by viruses and mycoplasmas, correct folding and the possibility to assemble multimeric proteins, often low downstream processing needed and the absence of ethical problems associated with the use of transgenic animals or animal-derived materials (1). A drawback for the use of plants is its specific glycosylation pattern, which is different from the human one, often resulting in allergic reactions when plant produced glycosylated proteins are administered to humans. In many cases proteins are produced in seeds (e.g. rice, corn) and the downstream processing becomes a limiting factor (2).
The moss Physcomitrella patens is becoming widely recognized as an experimental model of choice for basic molecular, cytological and developmental questions in plant biology (3). This moss can be genetically engineered to knock out the genes responsible for undesirable post-translational modifications (4). The dominant phase of its life cycle is in the gametophyte phase, which is haploid. Moreover, it is feasible to target any gene of interest to a well-known naturally highly expressed locus. P. patens can be cultivated in many different ways including bioreactors. Production cost is reduced by the use of simple growth media of inorganic salts with airborne CO2 as sole carbon source.
The vegetative stage of the life cycle in these bioreactors makes sure that there are no genetic instabilities and allows a stable production. For all these reasons, P. patens provides an ideal system for the heterologous expression of proteins and/or metabolites (4). Here we show the production of the plant pathogen-related protein (PRP) osmotin in P. patens. Osmotin is a small (26 kDa) class 5 PRP protein originally isolated from tobacco plants (5). Very low concentrations (4 µM) can kill S. cerevisiae (BWG1-7a strain) cells in a matter of minutes. It binds to the S. cerevisiae Pho36 receptor thereby inducing apoptosis in a Ras2-dependent manner. Its capacity to induce apoptosis makes osmotin an interesting antifungal protein and it has previously been proven to inhibit the growth of several other fungal species (6).
MATERIALS AND METODS
Plant Material and growth conditions
Wild type Physcomitrella patens Gransden strain was used for transformation experiments. Plants were propagated vegetatively as protonema homogenates on cellophane-overlaid BCD-AT(3) solid medium or as individual plants regenerated after protoplast transformation. Plants were incubated at 24°C under continuous light conditions.
Gene constructs and plant transformation
We have created an insertional construct targeted to the lambda 108 locus of P. patens (NCBI Accession: GQ250943). The construct contains Lambda 108 sequences on both sides, a highly expressed actin promoter, a signal peptide, a histidine tag, the tobacco osmotin gene, an alginate-binding domain, terminator sequence and a kanamycin cassette.
The plasmid pSAL-lambda 108 was created by cloning a 2kb fragment previously amplified from genomic DNA from P. patens by using the primers A5475 (5’ GGGTGATCAGTGTTTCGTTTAAACGAG TACCATTAAAGATGTTTC 3’) and A5476 (5’ GGGCTGCAGGTTGACTGTTTAAACTGACTTGGGAAATGAAATG 3’), the amplified Lambda108 sequence was inserted in the pSAL4 backbone by using the PstI and BclI restriction sites. The actin promoter sequence was amplified from genomic DNA of P. patens by using primers A5025 (5’ ATATGAAGCTTTCCTTGTGACTTTTGTGCATTATTG 3’) and A5026 (5’ CGGAATTCTATAGGATCCTATTTAATACGGACCTGCACAACA 3’).
The kanamycin cassette sequence was amplified by PCR with primers A5027 (5’ CGGAATTCCTACTCCAAAAATGTCAAAGATACAGTC 3’) and A5028 (5’ CCCAAGCTTAGTTTTGATCTTGAAAGATCTTTTATCTTT 3’) from plasmid DNA (pMBL5 plasmid). Both actin promoter and km cassette were cut at HindIII and EcoRI sites, and ligated to the HindIII site of the pSAL4-lambda 108 plasmid.
A synthetic construct was ordered with GENEART (Regensburg, Germany), containing in a single open reading frame, the signal peptide sequence (1-30 aa) from the xyloglucanendotransglycosylase/hydrolase precursor gene from P. patens (NCBI Accesion: CAH58715), a 6X histidine tag, the osmotin gene from Nicotianatabacum (NCBI Accesion: AAB23375) and the alginate binding domain (1-203 aa) of the protein P5 from Sphingomonas Sp. A1 (NCBI Accesion: LC043070). This synthetic construct was cloned at the sites BamHI and EcoRI of the plasmid pSAL4-lambda 108 containing the actin promoter and the Km cassette, generating the plasmid pSal4-Osmotin P5-BD. The plasmid pSAL4-Osmotin P5-BD was cut with PmeI enzyme, the 7452 bp containing the whole construct was gel-purified and used to target it to the lambda 108 genomic region by using the protoplast transformation method as described (7).
Transgenic selection was performed by using the antibiotic G418, and the correct insertion of the construct was verified by PCR and its expression by RT-PCR. One plant clone was selected for further experiments.
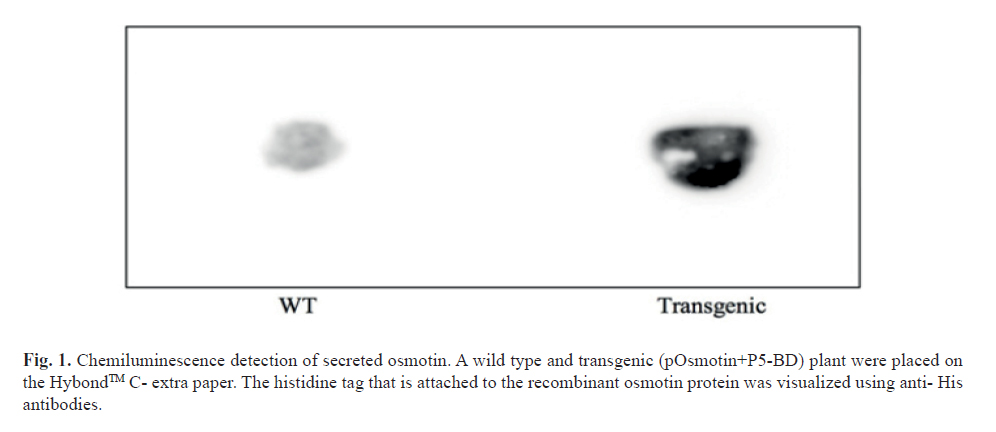
Detection of the protein by placing the plant on the HybondTM C- extra
Transgenic and Wt plants of approximately the same size were placed on a piece of HybondTM C- extra paper. The plants were left on this Hybond paper for two days at 25°C under continuous light conditions. Afterwards, plants were removed and the membrane was incubated for 1 hr at RT in 2% skimmed milk + TBST solution (20 mMTrisHCl pH 8, 150 mMNaCl, 0.05% Tween). Primary antibody (mouse anti-His antibody, GE Healthcare) was added at 1:3000 dilution in TBST and incubated overnight at 4°C. The membrane was washed 3X with TBST for respectively 5, 10 and 15 minutes; secondary antibody (anti-mouse antibody, GE Healthcare) was added at 1:5000 dilution and incubated 1hr at RT and washed 3X with TBST for 5 min and 1X with TBS for 5 min. 1 ml of Luminol and 1 ml of peroxide solution were added and incubated for 1-5 min, the paper was removed from the solution and visualized using the Fujifilm Las 4000 mini Chemiluminescence reader.
Evaluating biological activity of the protein
Normal BCDATG medium (7) was supplemented with 0,59 g/L of yeast complete supplement solution (Mpbio) containing essential nutrients for the yeast cells (leucine, adenine, histidine, uracyl and tryptophan) and 2% glucose. Yeast cells (S. cerevisiae BWG1-7a strain) were taken from a cell culture and grown overnight in 3 ml YPD at 30°C; 500 µl of this culture were centrifuged at 14,000 rpm for 10 seconds, the pellet was re-suspended in 100 µl and plated out on the supplemented BCDAT plates. Transgenic and a wild type plant tissue of approximately the same size were placed on the plates. The plates were incubated at room temperature for one week.
RESULTS AND DISCUSSION
Detection of secreted recombinant protein
To test if recombinant osmotin protein was produced and secreted by the transgenic plant, a wild type and transgenic plant were placed on HybondTM C-extra paper. In Figure 1 we can clearly see that the osmotin-construct containing plant showed a much higher chemiluminescence signal than the wild type plant. The wild type plant did show a low signal, probably due to non-specific binding of the antibodies to proteins produced (and secreted) by the moss. However, from this assay, we could conclude that the recombinant osmotin was present and was produced and secreted by the transgenic moss plant.
Evaluating biological activity of the protein
To test antifungal activity of the recombinant osmotin produced by the P. patens transgenic plant, tissue of a wild type and transgenic Physcomitrellaplant were placed on a thin layer of Saccharomyces cerevisiae BWG1-7a cells previously inoculated on BCDAT media. Growth and development of the moss was evaluated after 7 days. As is visible in Figure 2, the wild type plant became brown after one week and completely invaded by the yeast cells whereas the transgenic plant clearly shows an antifungal activity due to the production and secretion of a functional osmotin. Heterologous protein production in plants is becoming an increasing approach by pharmaceutical industry. Physcomitrella as a bioreactor has already shown many advantages over other plant systems (4). As a proof of concept, here we show the heterologous production of osmotin, a protein with fungicidal properties (6). Although it is necessary to further optimize the production and purification of the protein, we have clearly shown that the selected transgenic plant is able to secrete a biologically active osmotin protein as shown by the antibiogram assay against S. cerevisiae BWG1-7a strain (Fig. 2). These results demonstrate the capability of P. patens as a plant model system for heterologous protein production, supporting its use as bio-factory for the production of vaccines or other immunotherapeutic products.
REFERENCES
1. Desai PN, Shrivastava N, Padh H. Production of heterologous proteins in plants: Strategies for optimal expression. Biotechnol Adv. 2010;28(4):427–35.
2. Reski R, Frank W. Moss (Physcomitrella patens) functional genomics -- Gene discovery and tool development, with implications for crop plants and human health. Brief Funct Genomic Proteomic. 2005;4(1):48–57.
3. Cove D. The Moss Physcomitrella patens. Annu Rev Genet. 2005;39(1):339–58.
4. Decker EL, Reski R. Current achievements in the production of complex biopharmaceuticals with moss bioreactors. Bioprocess Biosyst Eng. 2008;31(1):3–9.
5. Singh NK, Nelson DE, Kuhn D, Hasegawa PM, Bressan RA. Molecular cloning of osmotin and regulation of its expression by ABA and adaptation to low water potential. Plant Physiol. 1989;90(3):1096–101.
6. Narasimhan ML, Damsz B, Coca MA, Ibeas JI, Yun DJ, Pardo JM, et al. A plant defense response effector induces microbial apoptosis. Mol Cell. 2001;8(4):921–30.
7. Kamisugi Y, Cuming AC, Cove DJ. Parameters determining the efficiency of gene targeting in the moss Physcomitrella patens. Nucleic Acids Res. 2005;33(19):1–10.
Recibido: Julio de 2017 Aceptado: Agosto de 2017


{kind=link}
{kind=link}