










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl v.26 n.1 La Habana ene.-mar. 2009
REPORT
Analysis of the genetic variability and immunological properties of the NlpB antigen, a novel protein identified in Neisseria meningitidis *
Maité Delgado1, Daniel Yero1,2, Olivia Niebla1, Gretel Sardiñas1, Sonia González1, Evelín Caballero1, Yanet Climent1,2, Darién García1, Karem Cobas1, Isbel Vivar1, Yusleydis Pérez1, Laritza Gorovaya1, Lázaro H Betancourt1, Yasser Perera1, Lila Castellanos1, Gerardo Guillén1
1Center for Genetic Engineering and Biotechnology, CIGB POBox 6162, Havana 10600, Cuba
2Finlay Institute, Ave 27 No. 19805, La Lisa, Havana, AP 16017, POBox 11600, Cuba
INTRODUCTION
Bacterial meningitis is still a pressing public health problem worldwide. Infections with Neisseria meningitidis, in particular, are a major contributor in the incidence of bacterial meningitis and septicemia, two devastating diseases that can claim the lives of children and young adults in a matter of hours. Both the developed and developing countries are equally affected by meningococcal infections, and examples of endemic or epidemic states can be found in both cases regardless of their geographical location (1).
Meningococcal disease, if left untreated, has a mortality rate of 60 to 80%. Although this figure can be reduced to 10% through the use of antibiotics, the antibiotic therapy does not eliminate or decrease the incidence of meningococcal colonization at the nasopharynx within the general population, while at the same time being expensive, and providing only short-term benefits; additionally, meningococcal disease often leaves permanent sequelae among survivors. The use of immunoprophylactic treatment is generally regarded, therefore, as the only effective means for the successful control of this infection (1).
Most cases of meningococcal disease are caused by serogroups A, B or C meningococci. Although efficacious polysaccharide-based vaccines have been available for a long time against serogroups such as A, C, W135 and Y (2), the capsular polysaccharide from serogroup B is poorly immunogenic due to its structural similarity with polysialic acid polymers present on the surface of human embryonic nervous tissue (3), and serogroup B vaccine research has therefore focused instead on subcapsular antigens such as the outer membrane proteins (OMP). Several different OMPs have been evaluated as vaccine candidates, either as individual subunits or as components of outer membrane vesicle (OMV) preparations. Although some of these formulations have shown promising results, their protective spectrum has so far been restricted to a relatively small number of antigenically related strains, and in general they fail to provide cross-protection against representative collections of epidemiologically relevant heterologous meningococcal strains.
The publication in the year 2000 of the genome sequence of a N. meningitidis serogroup B strain spurred a renewed interest in the search for new protein antigens that could induce protective immunity against this serogroup (4), using bioinformatic tools to process genomic and proteomic data. In this context, the present report summarizes the results obtained with these techniques at the Meningococcal Disease Department of the Center for Genetic Engineering and Biotechnology, which resulted in the identification of lipoprotein NlpB (annotated as NMB0928 in the genome sequence) as a component of the OMV from N. meningitidis, used as the active ingredient of the VAMENGOC- BC® vaccine. Additionally, this report presents our results in regard to obtaining and characterizing NlpB as a new vaccine candidate against meningococcal disease.
IDENTIFICATION OF NlpB AS A PUTATIVE EXPOSED OUTER MEMBRANE LIPOPROTEIN
Protein NMB0928 from N. meningitidis was first identified through proteomic techniques in the OMV of strain CU385, which constitutes the active ingredient of the Cuban VA-MENGOC-BC® vaccine (5). NlpB was later shown to be present, at consistent expression levels, in OMV from other meningococcal strains (6-8). An analysis by our group of the previously reported sequence for the nmb0928 gene and its genomic context led to the re-annotation of this gene and the potential cellular localization of its putative product (9), which had escaped previous in silico screenings for vaccine candidates against the meningococcus by other groups (10) due to an error in the original annotation (11). The polypeptide sequence of NMB0928 shows a typical Gram-negative secreted lipoprotein signal peptide and Leu-X-X-Cys motif, and although its physiological role is not known, it shares sequence homology with a family of bacterial lipoproteins that includes NlpB from Escherichia coli (a member of an outer membrane protein complex involved in the sorting and assembly of other secreted proteins (12)). This meningococcal NlpB homologue was, therefore, considered an attractive vaccine candidate.
SEQUENCE CONSERVATION OF THE NLPB GENE AND ITS DEDUCED AMINO ACID SEQUENCE
In order to study the degree of sequence conservation of this antigen, the gene coding for the NMB0928 NlpB homolog was amplified by PCR from a collection of 45 Cuban N. meningitidis strains isolated either from meningococcal disease patients or asymptomatic carriers, spanning a diverse range of serotypes, serogroups, serosubtypes and immunotypes that together represent all clonal complexes detected in Cuba before and after the countrywide immunization campaign against serogroups B and C, and including international reference strains for each serogroup. The analysis of the sequences generated in this study and the phenotypic characteristics of the source strains can be found at GenBank under access numbers EF688409 to EF688427 and FJ824137 to FJ824162. The coding sequence of nlpB was found in all strains analyzed, with a degree of sequence identity ranging from 90 to 100%. There was also a high degree of conservation for the amino acid sequence (Table 1), and although variable sites were detected at the N- and C-termini of the protein (9), the proportion of synonymous and non-synonymous codon changes in the gene (dn/ds) suggests that there is evolutionary pressure for the conservation of its sequence (Table 1).
CLONING AND EXPRESSION OF THE NlpB GENE IN A NOVEL EXPRESSION SYSTEM
Cloning and expression of the neisserial nlpB in E. coli was facilitated by the development, within our group, of a new expression system for this host. This system was based on plasmid pM83 (CIGB, Cuba), designed for the expression of heterologous proteins under the tryptophan promoter, which was engineered to contain a system for the selection of recombinant clones based on lacZ as a reporter gene within a bicistronic mRNA (Figure 1), an expanded multiple cloning site, and a sequence coding for a His(6) tag that can be fused, if desired, to the C-terminus of the recombinant polypeptide for easy downstream purification by immobilized-metal affinity chromatography (IMAC). The final plasmid obtained after these manipulations was named pM238 (13) (Figure 1) and has been widely used for obtaining a number of recombinant proteins in different departments and projects of CIGB. In our case, the gene coding for the NlpB protein was amplified by PCR from the CU385 vaccine strain (B:4:P1.19,15, ST=33) and inserted into pM238, thereby obtaining plasmid pM238-MB0928.
PURIFICATION OF RECOMBINANT NlpB
As mentioned above, plasmid pM238-NMB0928 codes for the synthesis of NlpB with an N-terminal tag consisting of the first 47 amino acids of the LpdA (P64k) protein from N. meningitidis and a C-terminal hexahistidine tag. After the transformation of E. coli W3110 with this plasmid, followed by experiments for the optimization of culture conditions, recombinant NlpB was obtained at expression levels of 30% of total protein contents. The relative mobility of the recombinant protein in denaturing SDS-polyacrylamide electrophoresis matches a molecular weight of 50 kDa, as expected for NlpB bearing the previously mentioned N- and C-terminal fusion tags. After the resuspension of the cell pellet on the TE buffer (10 mM Tris-Cl, 1 mM EDTA, pH 8) and a rupture by sonication (4 one-minute long cycles of sonication with two-minute intervals, on ice), recombinant NlpB was found to be associated to the insoluble fraction, probably as inclusion bodies. The protein was purified to 90% purity (as estimated by densitometry) by solubilization in a buffer containing 2 M urea, followed by denaturing chromatography on chelating Sepharose previously charged with copper sulfate and refolding by the dialysis of the purified fractions against carbonate-bicarbonate buffer pH 10, eliminating the chaotropic agent and the imidazole.
PREPARATION OF THE IMMUNOGENS AND IMMUNIZATION OF ANIMALS WITH RECOMBINANT NlpB
The immunological evaluation of recombinant NlpB was performed with 2 immunization schemes implemented in different animal models. The first scheme used outbred 7 day-old (neonate) OF1 mice of both sexes split into 4 groups of 8 animals each, maintained under controlled conditions throughout the experiment. The animals were inoculated intraperitoneally with 10 µg of the recombinant protein in a total volume of 50 µL/individual, withdrawing blood samples for evaluation 7 days after the last inoculation. In this case, groups 1 and 2 were immunized with NlpB adjuvanted either in aluminum hydroxide (1:40) or Freunds complete/incomplete adjuvant (1:1) respectively at days 7 and 14 after birth; groups 3 and 4 were immunized similarly but received three doses at 7, 10 and 14 days after birth.
The second scheme used 40 female Balb/c (H-2d) mice 6 to 7 weeks old with a weight ranging from 18 to 20 g, supplied by CENBALAB, which were split into 4 groups of 10 animals each immunized subcutaneously. Group 1 received 20 µg of NlpB with Freunds adjuvant, group 2 received 20 µg of NlpB adjuvanted with aluminum hydroxide, group 3 received 20 µg of NlpB inserted into multilamellar liposomes and adjuvanted into aluminum hydroxide, and group 4 received 20 µg of protein P6 from Haemophilus influenzae (unrelated control) in Freunds adjuvant.
The design of the immunogen for group 3 is based on the fact that NlpB is a membrane protein, and therefore the insertion into multilamellar liposomes may promote the correct folding of this molecule by mimicking its native physical environment. The liposomes were obtained by the method of vesicle reconstitution by dehydration-hydration (14), incorporating the recombinant antigen as described previously (9). Briefly, the empty liposome suspension was mixed with 6 mg of recombinant protein and dehydrated by freezing 3 times. Finally, the vesicles were centrifuged at 100 000 x g for 2 h at 4ºC and the resulting pellet was resuspended into 1 mL of PBS.
CHARACTERIZATION OF THE IMMUNE RESPONSE IN DIFFERENT ANIMAL MODELS
The analysis of the humoral immune response against the homologous antigen in the murine neonate model described above (which tries to simulate the immune status of newborns) showed an antibody titer larger than 1000 in all individuals, which did not increase after a third dose. These results suggest that the antigen is immunogenic even in infants, which constitute one of the age groups with the highest incidence of meningococcal disease. The protein was also immunogenic in the model of adult mice, with titers ranging from 170 000 to 400 000 depending on the specific adjuvant (9). An analysis by Western blot of the serum samples from immunized mice against cell extracts of N. meningitidis showed that the induced antibodies specifically recognize the natural protein, with a relative mobility corresponding to ~40 kDa. The sera also showed bactericidal activity against the homologous strain (CU385) with titers ranging from 1:16 to 1:32. The evidence suggests that bactericidal activity is a good correlate of protection in humans for OMV-based anti-meningococcal vaccines specific to serogroup B (15), and this criterion as been extended to the evaluation of new protein-based vaccine candidates in mice as well.
The sera were also assayed in a model of passive immunization and challenge in infant rats. In this experiment, newborn rats with an age of 5 to 6 days were intraperitoneally inoculated with a 1:10 dilution of sera from mice immunized with the recombinant protein, and were then challenged 1 hour later with approximately 107 colony-forming units (7 log10 cfu) of the homologous strain (CU385) in the presence of an external iron source (iron-dextran). Four hours after the challenge, blood samples were withdrawn from each rat and bacteremia was measured by cfu counts in BHI-blood agar plates. The results of these analyses showed that the immunization of mice with recombinant NlpB inserted into liposomes induces a protective response, since the sera from these animals promoted a significant reduction in bacteremia for the challenged rats compared to a control serum obtained before the inoculation (5.26 ± 0.94 vs. 6.16 ± 0.34 log10 cfu/mL, P < 0.05). The presentation of this antigen in liposomes, therefore, probably leads to the adoption of a native-like structure that exposes B-cell epitopes to the immune system, which are required for a protective humoral response (9).
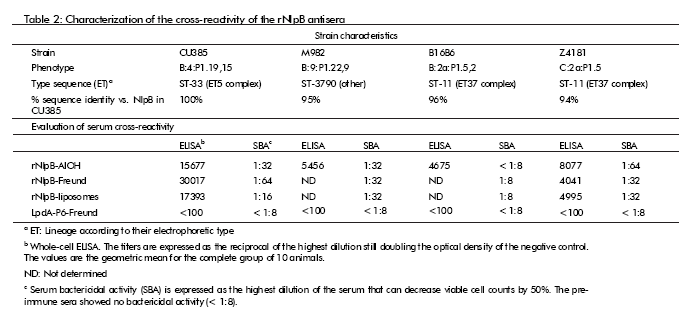
CROSS-REACTIVITY OF THE ANTI-rNlpB SERA
The cross-reactivity of the sera generated by immunizing mice with recombinant NlpB was examined in whole-cell ELISA and bactericidal activity assays against 3 heterologous strains with differing serogroups, serotypes, serosubtypes and genotypes and varying degrees of divergence in their nlpB gene compared to the sequence found in strain CU385. The antisera were able to recognize the natural protein exposed in intact cells from all three strains in the ELISA assay, as well as to mediate complement-dependent bacterial killing with bactericidal titers from 1:8 to 1:64 in all the strains examined (Table 2). The control samples from animals immunized with an unrelated protein (antigen P6 from Haemophilus influenzae) did not show any reactivity in these assays. Additionally, we were able to demonstrate the presence of the NlpB gene product in all strains by Western blot (data not shown) (9).
CONCLUSIONS
The findings here reported lead us to consider the inclusion of this antigen into a meningococcal vaccine preparation, given the degree of sequence conservation of NlpB in the collection of strains studied. The changes that were detected in the amino acid sequence do not eliminate the cross-recognition of intact cells from heterologous strains by the sera obtained by immunization with NlpB, and the antibodies also have bactericidal activity and protect against a meningococcal challenge in the infant rat model. To be considered a wide-spectrum vaccine candidate, however, the evaluation of NlpB-specific antisera should include a larger collection of meningococcal strains. Such an investigation is currently underway in our department not only for this antigen, but for other proteins that have also generated protective immunity in preclinical assays.
REFERENCES
1. Stephens DS. Conquering the meningococcus. FEMS Microbiol Rev 2007;31(1):3-14.
2. Pace D. Quadrivalent meningococcal ACYW-135 glycoconjugate vaccine for broader protection from infancy. Expert Rev Vaccines 2009;8(5):529-42.
3. Finne J, Bitter-Suermann D, Goridis C, Finne U. An IgG monoclonal antibody to group B meningococci cross-reacts with developmentally regulated polysialic acid units of glycoproteins in neural and extraneural tissues. J Immunol 1987;138(12):4402-7.
4. Pizza M, Scarlato V, Masignani V, Giuliani MM, Arico B, Comanducci M, et al. Identification of vaccine candidates against serogroup B meningococcus by whole-genome sequencing. Science 2000;287(5459):1816-20.
5. Uli L, Castellanos-Serra L, Betancourt L, Domínguez F, Barbera R, Sotolongo F, et al. Outer membrane vesicles of the VAMENGOC- BC® vaccine against serogroup B of Neisseria meningitidis: Analysis of protein components by twodimensional gel electrophoresis and mass spectrometry. Proteomics 2006;6(11):3389-99.
6. Vipond C, Suker J, Jones C, Tang C, Feavers IM, Wheeler JX. Proteomic analysis of a meningococcal outer membrane vesicle vaccine prepared from the group B strain NZ98/254. Proteomics 2006;6(11):3400-13.
7. Vaughan TE, Skipp PJ, OConnor CD, Hudson MJ, Vipond R, Elmore MJ, et al. Proteomic analysis of Neisseria lactamica and Neisseria meningitidis outer membrane vesicle vaccine antigens. Vaccine 2006;24(25):5277-93.
8. Ferrari G, Garaguso I, Adu-Bobie J, Doro F, Taddei AR, Biolchi A, et al. Outer membrane vesicles from group B Neisseria meningitidis delta gna33 mutant: proteomic and immunological comparison with detergent-derived outer membrane vesicles. Proteomics 2006;6(6):1856-66.
9. Delgado M, Yero D, Niebla O, González S, Climent Y, Perez Y, et al. Lipoprotein NMB0928 from Neisseria meningitidis serogroup B as a novel vaccine candidate. Vaccine 2007;25(50):8420-31.
10. Pizza M, Scarlato V, Masignani V, Giuliani MM, Arico B, Comanducci M, et al. Identification of vaccine candidates against serogroup B meningococcus by whole genome sequencing. Science 2000;287(5459):1816-20.
11. Tettelin H, Saunders NJ, Heidelberg J, Jeffries AC, Nelson KE, Eisen JA, et al. Complete genome sequence of Neisseria meningitidis serogroup B strain MC58. Science 2000;287(5459):1809-15.
12. Misra R. First glimpse of the crystal structure of YaeTs POTRA domains. ACS Chem Biol 2007;2(10):649-51.
13. Yero D, Pajon R, Niebla O, Sardinas G, Vivar I, Perera Y, et al. Bicistronic expression plasmid for the rapid production of recombinant fused proteins in Escherichia coli.Biotechnol Apply Biochem 2006;44(Pt (1)):27-34.
14. Kirby CJ, Gregoriadis G. Preparation of liposomes containing factor VIII for oral treatment of haemophilia.J Microencapsul 1984;1(1):33-45.
15. Borrow R, Balmer P, Miller E. Meningococcal surrogates of protectionserum bactericidal antibody activity. Vaccine 2005;23(17-18):2222-7.
*This work received the Award of the National Academy of Sciences of Cuba for the year 2008.
Maité Delgado. Center for Genetic Engineering and Biotechnology, CIGB. POBox 6162, Havana 10600, Havana, Cuba. E-mail: maite.delgado@cigb.edu.cu

{kind=link}
{kind=link}
{kind=link}