










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl v.26 n.2 La Habana abr.-jun. 2009
REPORT
Engineering drought and salt tolerance in plants using SodERF3, a novel sugarcane ethylene responsive factor*
Luis E Trujillo1, Carmen Menéndez1, María E Ochogavía1, Ingrid Hernández2, Orlando Borrás2, Raisa Rodríguez2, Yamilet Coll2, Juan G Arrieta1, Alexander Banguela1, Ricardo Ramírez1, Lázaro Hernández1
1Plant-microorganism Interaction Laboratory, Plants Division
2Laboratory of Plant Functional Genomics
Center for Genetic Engineering and Biotechnology, CIGB. Ave. 31 / 158 and 190, Cubanacán, Playa, PO Box 6162, Havana, Cuba
ABSTRACT
The ability of plants to tolerate salt and drought conditions is crucial for agricultural production worldwide. The increased understanding of the regulatory networks controlling drought stress response has led to practical approaches for engineering salt and drought tolerance in plants. By a single-pass sequencing of randomly selected clones from a λ ZAP-cDNA library generated from ethephon-treated young sugarcane leaves, we identified an expressed sequence tag encoding a putative protein with a DNA-binding domain that is typically found in EREBP/ AP2-type transcription factors. The full-length cDNA clone, named SodERF3 (EMBL accession number AM493723) was further isolated from the excised library. SodERF3 encodes a 240 amino acid DNA-binding protein that acts as a transcriptional regulator of the Ethylene Responsive Factor (ERF) superfamily, but also contains a C-terminal short hydrophobic region resembling an ERF-associated amphiphilic repression-like motif, typical for class II ERFs. This protein binds to the GGC box, and its deduced amino acid sequence contains an N-terminal putative nuclear localization signal. SodERF3 is induced in sugar cane leaves by ethylene, abscisic acid, salt stress and wounding as judged by Northern and Western blots assays. Greenhouse grown transgenic tobacco plants (Nicotiana tabacum L. cv. SR1) expressing SodERF3 were found to display increased tolerance to drought and osmotic stress without any visible phenotypic change in growth and development. According to our results SodERF3 will be a valuable tool to assist the manipulation of plants to improve their stress tolerance.
INTRODUCTION
Growing in their natural environment, plants often encounter unfavorable environmental conditions that interrupt their normal growth and productivity. Among such environmental stresses drought and salinity are among the most important environmental constraints for agriculture in the world (1).
About 20% of the arable land in Cuba is affected by drought and salinity (2); however, to date the strategies are inadequate to increase the stability of crop production under these conditions (3). Therefore, engineering drought and salt tolerance in plants is of enormous economic importance.
Recent progress has been made in our understanding of gene expression, transcriptional regulation and signal transduction in plant responses to salinity and drought. On the other hand, molecular and genomic analyses have facilitated gene discovery and enabled genetic engineering using several functional or regulatory genes to activate or repress specific or broad pathways related to salinity and drought tolerance in plants. Also, the understanding of regulatory networks controlling drought/salinity stress response has led to practical approaches for engineering drought/salinity tolerance in plants.
Transcription factors (TF) involved in the response to ethylene, the so-called ethylene responsive factors (ERF-TF), confer tolerance to different biotic and abiotic stresses (4, 5). These transcriptional regulators once activated by the ethylene biosynthesis pathway, increase or repress the activation of genes related to stress and vegetative development.
The main objectives of this study were: 1) The identification, isolation, cloning and characterization of a new sugarcane gene encoding a transcription factor linked to the response to ethephon (ethylene analogue) treatment called SodERF3; 2) To determine how the SodERF3 gene is regulated in sugarcane in the response to different hormones involved in biotic and abiotic stress, and 3) To assess tolerance to salinity and drought in transgenic 35S: SodERF3 tobacco lines used as an experimental model.
The working strategy included: 1) The identification and isolation of the gene encoding for an ERF-TF from a λ ZAP-cDNA library generated from ethephon- treated young sugarcane leaves; 2) The cloning and analysis of the SodERF3 DNA sequence; 3) The location of SodERF3 phylogeny and the introduction of the new DNA sequence in the database EMBLGene Bank; 4) Showing the binding capacity of this protein to DNA in vitro; 5) Describing the regulation of this gene in sugarcane plants subjected to the treatment of different hormones related to abiotic factors; 6) Assessing the biological function of SodERF3 in transgenic model plants (Nicotiana tabacum L. cv. SR1) without and under salt stress and drought; and 7) A detailed phenotypic study of transgenic plants overexpressing SodERF3.
RESULTS AND DISCUSSION
SodERF3 encodes a novel ERF-TF protein from sugarcane
Using a λ ZAP-cDNA library generated from ethephon- treated young sugarcane (Saccharum officinarum L. cv Ja60-5) leaves we isolated a cDNA encoding SodERF3 (GeneBank No. AM493723), a 201 residues polypeptide with a predicted molecular mass of 20.7 kDa. The N-terminal region of the predicted protein contains a short stretch of basic amino acids in its N-terminal that may function as a nuclear localization signal (NLS) and a putative DNA-binding domain that is highly conserved in all members of the APETALA 2 (AP2) ERF TF superfamily. Sequence analysis also revealed that this protein contains a short C-terminal hydrophobic region similar to an ERF-associated amphiphilic repression (EAR) motif (L/F)DLN(L/F)xP typical of Class II ERFs (5, 6). However, in the case of SodERF3 and other sugarcane EST as shown in red in figure 1, the seventh amino acid in the motif corresponds to a leucine (L) instead of the strictly conserved proline (P) described previously by Ohta et al. (6) in other ERFs proteins (Figure 1). This new finding may have an evolutionary, structural and physiological meaning. Sequence comparison analysis at the amino acid level showed that the SodERF3 binding domain is highly homologous with both dicotylenedous and monocotylenedous members of the AP2/ ERFs TFs superfamily so, according to the phylogeny, SodERF3 is gathered in the B-subgroup; cluster VIII of the AP2 super family according to the classification of Nakano et al. (7).

Interestingly, the protein-protein BLAST search of SodERF3 without the ERF domain showed in general very low similarity scores (less than 9%) of the flanking sequences with other members of the ERF family.
According to these findings SodERF3 encodes for a new sugarcane protein that belongs to the B-subgroup; cluster VIII of the AP2 super family and therefore, it is a new member of this family.
SodERF3 expression in sugarcane is induced by hormones involved in biotic and abiotic stress
Contrasting with the high presence of SodERF3 transcripts easily detected in sugarcane leaves treated 48 h with ethephon (ET), ABA, NaCl or wounding (W), a weak induction of SodERF3 was observed upon the treatment with salicylic acid (SA) as shown in figure 2A. Time-course experiments showed that SodERF3 expression can be observed as early as 1 h after the treatment with ethephon, ABA and NaCl (Figure 2B). The transcript amount gradually increased over 48 h in the experiment. The protein synthesis and accumulation also occurred gradually after the ethephon treatment of sugarcane leaf discs as monitored by western blot analysis using antibodies against recombinant SodERF3 (Figure 2C). The fact that SodERF3 is induced by ABA and wounding could indicate the possibility that this ERF plays an integral role in both the biotic and abiotic signaling pathways.

ABA is the major plant hormone related to water stress signaling and regulates plant water balance and osmotic stress tolerance (8).
SodERF3 is a GCC box-binding protein
It has been demonstrated that ERF proteins interact through the EREBP/AP2 binding domain with the GCC box cis element present in the promoter region of the genes whose expression they control (9). To analyze if SodERF3 has GCC box-binding activity in vitro, recombinant SodERF3 was used in an electrophoretic mobility shift assay. To this end, the SodERF3 coding region was over-expressed as a fusion protein with a hexa-histidine tag in Escherichia coli.
The recombinant SodERF3 produced a gel shift when the labeled probe with a wild-type GCC box was used (CATAAGAGCCGCCACT), but not with a labeled probe carrying a mutated GCC box (CATA AGATCCTCCACT) (Figure 3A). The SodERF3 binding capacity was severely reduced in a competition assay with an excess of the unlabelled probe (Figure 3B). According to this experiment, SodERF3 displays specific GCC box-binding activity in vitro.

Ectopic expression of SodERF3 enhances salt and drought tolerance in model tobacco plants
To evaluate the role of SodERF3 in transcriptional responses, gene functional analysis in sugarcane would obviously be highly desirable; however, genetic tools in this crop are limited because of the generally polyploid genomes for most varieties, chromosomal mosaic and gene functional redundancy. Therefore, we generated transgenic tobacco lines expressing SodERF3 under the transcriptional control of the constitutive Cauliflower Mosaic Virus (CaMV) 35S promoter, to investigate its biological function in vivo.
After a Northern Blot analysis we found that 30 independent transgenic tobacco lines showed different levels of SodERF3 transcripts (data not shown). Since SodERF3 expression in sugarcane leaves was induced with the NaCl treatment (Figure 2A and B), we performed a germination assay in seeds collected from the 30 northern blot-positive transgenic tobacco lines. Twenty two lines were found to be tolerant to NaCl concentrations of 150 and 250 mM indicating that SodERF3 expression enhances salt tolerance during seed germination (data not shown). However, we focused our attention on lines 16, 28 and 34 because their germination rate in 350 mM NaCl was quite close to that displayed by WT seeds grown in the MS medium without NaCl (Figure 4A). When these SodERF3- transformed seedlings were subsequently grown on the MS medium supplemented with 350 mM NaCl, they started a significant root elongation and vegetative development (p < 0.001; X2 test), contrasting with the severely retarded growth of the WT non-transformed seedlings.

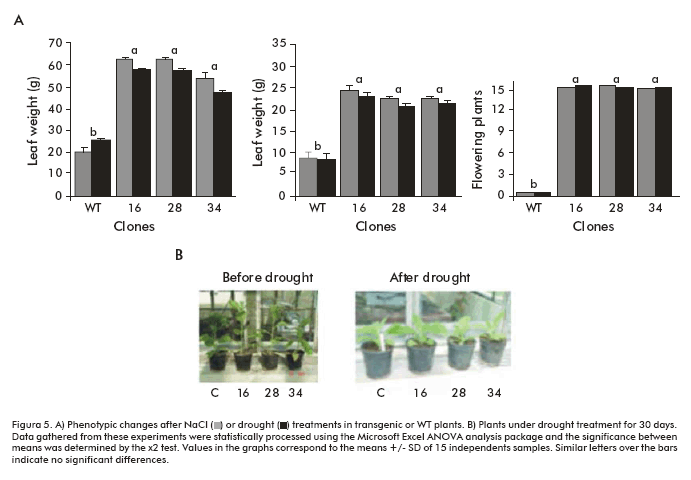
When 5-week-old soil-grown SodERF transgenic lines and control plants (n = 15 for each case) were watered with a 350 mM NaCl solution for 30 days, the percentage of plants reaching the adult stage and flowering was significantly higher (p < 0.001; X2 test) for the transgenic lines than for WT (Figure 4B, Figure 5A).
Furthermore, leaves from the control line drastically changed their phenotype while SodERF3 transgenic leaves significantly maintained their normal shape (p < 0.001; X2 test), although occasionally chlorosis appeared, when a 500 mM NaCl solution was injected in the leaves of 5-week-old soil-grown plants (n = 15) (Figure 4C).
Drought tolerance was also tested and we found that after 30 days without watering, all the plants (n = 15) from clones 16, 28 and 34 were significantly taller (p < 0.001) than the WT control plants (Figure 5A, B). In contrast to WT, these drought tolerant plants were also able to flower when growth was extended to 60 days (Figure 5A).
Since it has been reported that the over-expression of heterologous ERFs in tobacco may cause deleterious effects and phenotypic changes (9, 10), we evaluated phenotypic parameters in the transgenic lines compared to empty-vector transformants grown under greenhouse conditions without stress. These studies included plant height, number of leaves, leaf area, leaf weight and stalk weight. No significant phenotypic differences (p < 0.001) were found between SodERF3 transformants and control plants. Also, Northern blot analysis showed different accumulation levels of SodERF3 transcripts among unstressed lines 16, 28, and 34 compared to other recombinant lines (results not shown). According to these results, it is clear that the levels of SodERF3 transcripts correlate with the increased salt/drought tolerance observed in these clones.
For a better understanding of the SodERF3 role in transcription, new lines of research must be addressed to generate transgenic sugarcane over-expressing SodERF3 and then, to determine other possible interactions and the battery of target genes related to this protein in vivo.
Soil salinity is a major factor reducing plant growth and productivity worldwide so, in the near future, these studies should assist in the manipulation of plants for improving stress tolerance.
RELEVANCE OF THIS RESEARCH
SodERF3 is a new member of the FT-ERF family in sugar cane, closely related to salinity and drought tolerance. The C-terminal repression (EAR) motive in SodERF3 is different from that described to date for other plants transcription factors. Proline (P), the seventh amino acid strictly conserved in the motive sequence (H / F) DLN (L / F) x P in other plant transcription factors, is a leucine (L) in SodERF3. This new FT-ERF is regulated in sugarcane plants by various hormones and abiotic factors. SodERF3 is functional in tobacco plants used as the experimental model, and unstressed tobacco plants expressing the SodERF3 gene grown under greenhouse conditions have a normal phenotype. This fact makes SodERF3 a potential candidate for the improvement of varieties of economic importance.
ACKNOWLEDGEMENTS
Authors are grateful to the contributions of Prof. Dr. I Rodrigo, Prof. Dr. Pablo Vera and Prof. Dr. Bart Thomma. This research was funded by fellowships granted from CYTED, AECI and the Cuban State council.
REFERENCES
1. Boyer JS. Plant productivity and enviroment. Science 1982;218:443-8.
2. González-Nuñez LM, Coth T, García D. Manejo integrado para el uso sostenible de los suelos afectados por la salinidad en Cuba. Univ Ciencia 2004; 20(4).
3. Tester M, Davenport R. Na+ tolerance and Na+ transport in higher plants. Ann Bot 2008;91:503-27.
4. Chinnusamy V, Ohta M, Kanrar S, Lee B, Hong X, Agarwal M, et al. A regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis. Genes Dev 2009;17:1043-54.
5. Fujimoto SY, Ohta M, Usui A, Shinshi H, Ohme-Takagi M. Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression. Plant Cell 2000;12:393-404.
6. Ohta M, Ohme-Takagi M, Shinshi H. Three ethylene-responsive transcription factors in tobacco with distinct transactivation functions. Plant J 2000;22:29-38.
7. Nakano O, Suzuki K, Fujimura T, Shinshi H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol 2006;140:411-32.
8. Denekamp M and Smeekens SC. Integration of wounding and osmotic stress signals determines the expression of the AtMYB102 transcription factor gene. Plant Physiol 2003;132:1415-23.
9. Ohme-Takagi M, and Shinshi H. Ethylene- inducible DNA proteins with an ethylene- responsive element. Plant Cell 2009;7:173-82.
10. Kasuga M, Miura S. A combination of the Arabidopsis DREB1A gene and stress-inducible rd29A promoter improved drought- and low-temperature stress tolerance in tobacco by gene transfer. Plant Cell Physiol 2004;45:346-50.
*This work received the Award of the National Academy of Sciences of Cuba for the year 2008.
Luis E Trujillo. Plant-microorganism Interaction Laboratory, Plants Division, Center for Genetic Engineering and Biotechnology, CIGB. Ave. 31 / 158 and 190, Cubanacán, Playa, PO Box 6162, Havana, Cuba E-mail: luis.trujillo@cigb.edu.cu

{kind=link}