










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl v.27 n.3 La Habana jul.-sep. 2010
REPORT
Demonstration by RNA interference of a new molecular mechanism for resistance to an oomycete in tobacco plants
Ingrid Hernández1, Orlando Borrás1, Osmani Chacón2, Merardo Pujol1, Yunior López1, Raisa Rodrígues1, Roxana Portieles1
1 Plant Division, Laboratory of Vegetal Functional Genomics, Center for Genetic Engineering and Biotechnology, CIGB Ave. 31 / 158 and 190, Cubanacán, Playa, PO Box 6162, Havana, Cuba
2 Institute for Investigations on Tobacco, IIT
ABSTRACT
Oomycetes constitute an important agricultural pathogen for plants of the Solanaceae family. Black shank, caused by Phytophthora parasitica var. nicotianae, is one of the most important diseases affecting tobacco plantations in Cuba and worldwide, constituting an occasional cause of major economic losses. We have constructed a subtractive library with the aim of identifying tobacco genes involved in the resistance to this oomycete. The analysis by realtime PCR confirmed, for the first time, the patterns of differential expression between resistance and susceptibility reactions for the identified genes. This constitutes the first report demonstrating the pattern of expression of the glutathione-S-transferase (GST) gene in varieties and species independently of the degree of resistance or susceptibility to the pathogen. The gene was cloned in a plant transformation vector for gene silencing, and transgenic N. tabacum plants with silenced GST were obtained. They significantly displayed increased resistance to the disease, in highly susceptible plants, in comparison to control plants. This work constitutes the first report on the role of GST as a negative regulator of the defensive response to oomycetes, as well as the first instance where RNA interference technology has ever been used to obtain resistance to diseases produced by Oomycetes.
INTRODUCTION
Black shank, caused by Phytophthora parasitica var. nicotianae, is one of the most important agricultural pathogens for tobacco (Nicotiana tabacum) worldwide (1). This pathogen attacks the roots, stem and leaves in all developmental stages, producing root necrosis, withering and shriveling, chlorosis, stem lesions, atrophy and, ultimately, death. The disease progresses rapidly under conditions of high humidity and temperature (2). Annually, agricultural losses due to this mold are significant, resulting in important financial costs (1).
The disease is usually managed with an integrated approach that includes specific cultural practices, the application of fungicides and the use of resistant cultivars. However, the use of resistant cultivars constitutes by far the most effective measure (3). Both N. longiflora and N. plumbaginifolia have been shown to be resistant to race 0 of P. parasitica var. nicotianae (4), and cultivars inheriting this resistance are immune (i.e. no growth or reproduction of the pathogen takes place).
Nicotiana megalosiphon is a wild tobacco species commonly used in genetic improvement programs for tobacco (5). N. megalosiphon is highly resistant to oomycetes Phytophthora parasitica var. nicotianae and Peronospora hyoscyami f. sp. tabacina (6). However, this species is highly susceptible to viral pathogens such as virus A of potato and TMTV (7).
Several studies have approached the tobacco - P. parasitica var. nicotianae interaction from a molecular perspective. The activity of lipoxygenase (LOX) and the expression of the lox gene upon infection with P. parasitica var. nicotianae, for instance, have been studied in intact tobacco plants, observing a rapid and temporary accumulation of LOX after the inoculation with P. parasitica var. nicotianae zoospores. This result suggests an important role for the lox gene in mediating resistance to the pathogen and establishing a defensive state in the host plant (8). The defensive state against P. parasitica var. nicotianae has also been associated with an extracellular cytotoxic activity originated from the accumulation of salicylic aciddependant PR-1 proteins in the apoplasm, as well as a salicylic acid-independent cytotoxic activity which has not been detected in leaves in which a systemic acquired resistance has been induced (9).
On the other hand, the transition from the vegetative to the reproductive state also produces a transition from susceptibility to resistance to the causal agent of black shank. This resistance manifests as it decreases in infection efficiency and hyphal expansion, and has also been found to be associated to the accumulation of an extracellular cytotoxic activity. Microarray analyses and ribonucleic acid hybridization revealed that during late developmental stages, leaves are characterized by a coordinated regulation of genes involved in the acquired systemic response and the induction of structural proteins of oxidation in the cell wall (10).
Additionally, the infection of plant cells by a pathogen usually elicits a battery of disease response genes, which can result in the production of several toxic products that include reactive oxygen species (11). One of such responses observed in plants after infection is the increased expression of genes coding for glutathione-S-transferase (GST) (12).
Although the involvement of GST of N. tabacum on the response to cold, salt stress and aluminuminduced toxicity is established (13), little is known about its role during pathogenic infection. Using binding primers to preserved regions of N. tabacum GST, four different genes for this enzyme were amplified from cDNA of Nicotiana benthamiana leaves infected with Colletotrichum destructivum (14). After cloning each of these genes on the PVX vector for virus-induced gene silencing and obtaining the corresponding transgenic plants, a significant increase in susceptibility to the infection was observed only in individuals bearing a silenced NbGSTU1 gene. These plants showed a larger number of lesions (130%) and more extensive C. orbiculare colonization (67%) than the controls (14), proving that the different GST variants differently respond to fungal infections, and demonstrating the involvement of at least one of them in the development of the disease (14).
Taking into account the importance of dissecting the molecular components in Nicotiana tabacum responsible for establishing a state of resistance or susceptibility to P. parasitica var. nicotianae, as well as the need for exploring new strategies for harnessing disease resistance, the objectives of the present work were: To obtain a cDNA library formed by transcripts derived from genes induced during the resistance interaction with P. parasitica var. nicotianae; to characterize by realtime PCR the expression of genes involved in resistance to the disease; to evaluate the role of resistance- or susceptibility-related genes by using RNA interference technology, and to characterize the expression of a set of defense genes in the tobacco with RNA interference, using realtime PCR.
RESULTS AND DISCUSSION
Obtention of a cDNA library formed by transcripts derived from genes induced during the resistance interaction with P. parasitica var. nicotianae
This work identified a set of genes involved in the response against P. parasitica var. nicotianae in N.
tabacum which had not been previously shown to participate in this phenomenon. Although the functional assignments for these genes were made based only on sequence homologies to known orthologs -and therefore demand a stricter experimental verification-, this step offers, however, a clue about the genetic diversity of this response.
A total of 65 clones were obtained, of which 48 were sequenced. For nearly 40% of the clones no known orthologs were found; these were classified, therefore, as having an unknown function. The genes that did have significant sequence homologies to known orthologs fell into six primary functional categories. Most of these genes (15%) were defense-related, followed by those others involved in signal transduction, metabolism and protein synthesis (10%). Genes involved in energy constituted 8% of the cDNA collection, and those involved with transport constituted 7% of the sequences (15).
Characterization by realtime PCR of the expression of genes involved in resistance to the disease
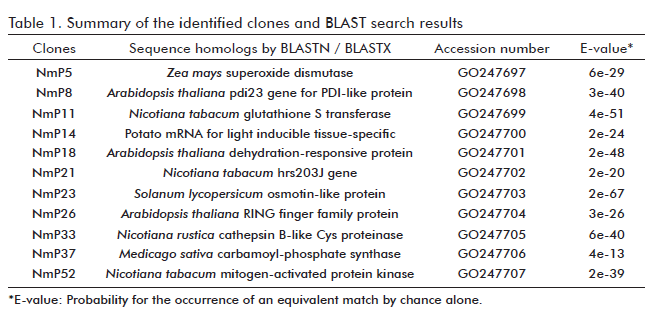
Based on the homologies detected in public sequence databases, 11 genes with a significant E-value in BLAST searches were selected for further analysis of their expression patterns (Table 1). Quantitative PCR assays were performed on RNA extracted from resistant (N. megalosiphon) or susceptible (N. tabacum var. Sumatra) plants at 0, 3, 7 and 10 days post-inoculation with P. parasitica var. nicotianae. This analysis indicated that the levels of expression of the transcripts from all 11 genes were generally higher during the resistance interaction and lower during the susceptible interaction (15).
Gene hrs203J and the one coding for the RING fi nger protein displayed a rapid and intense induction during the resistance interaction, but were undetectable during susceptibility interaction. This indicates that they might be important candidates for the evaluation of disease-resistant genotypes within the framework of crop improvement programs (Table 2). Significantly, a clone homologous to GST showed the highest induction during the resistance and susceptibility interaction in all evaluated time points, generating a hypothesis about its possible involvement in resistance and susceptibility that was later tested through a functional analysis examining its potential role as negative resistance regulator.
Evaluation of the role of genes implicated in resistance and susceptibility by using RNA interference technology
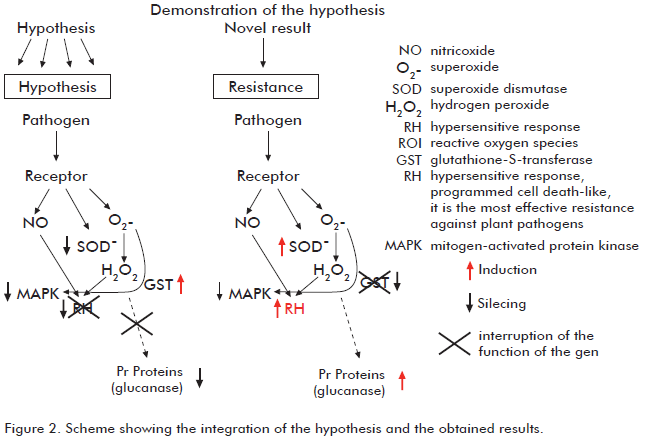
This study analyzed the biological relevance of the GST gene during the susceptibility interaction between P. parasitica var. nicotianae - tobacco by silencing this gene in transgenic tobacco plants. Surprisingly, the plants with silenced GST displayed higher levels of resistance to the pathogen, with decreased withering of the stem (Figure 1). One of the possible reasons for this result is that the expression of GST during the susceptibility interaction might be limiting the hypersensitive response (HR), according to the expression of some genes involved in this type os response such as SOD and Hsr203J (18).
A number of studies on the HR in a variety of experimental systems have demonstrated the involvement of reactive oxygen species such as the superoxide anion (O2 -) and hydrogen peroxide (H2O2) as important signals or effectors of programmed cell death. O2- can promote the HR through the production of H2O2 or repress it through the elimination of nitric oxide or the induction of antioxidants. In this scheme, SOD is an important mediator to compensate these functions.
Characterization of the expression of a set of defense genes in tobacco plants with RNA interference using realtime PCR
Relative expression levels of several genes associated to disease resistance in plants were evaluated in transgenic tobacco plants where GST had been silenced. The genes coding for beta-1,3-glucanase, superoxide dismutase (SOD), mitogen-activated protein kinase (MAPK) and the cell-death associated protein from N. tabacum (Hsr203J) displayed a higher level of expression in the plants bearing RNAi construction compared to the control plants, where their expression was lower or delayed in time. On the other hand, the expression of the gene for phenylalanine-ammonia lyase (PAL) was repressed in the RNAi-transformed lines, but induced in the controls. Results showed that silencing the GST gene contributes to the disease-resistant phenotype as negative regulator of innate immunity to black shank in susceptible plants (18).
Silencing GST led to reduced expression of the PAL gene, which codes for a key enzyme in the phytoalexin and salicylic acid synthesis pathways (18). Also, as mentioned above, the plants with silenced GST displayed increased expression not only of the genes coding for Hsr203J, SOD and MAPK, but also for beta- 1,3-glucanase, a well-known gene coding for a protein involved in pathogenesis. Possibly, the resistant phenotype observed in this study as a result of GST silencing is actually the outcome from the combined action of several gene products under direct or indirect control by GST (17).
Finally, it should be pointed out that during the compatible (susceptible) interaction the pathogen induces what is known as a hypersensitive response in the host plant through the production of reactive oxygen species and nitric oxide. However, there is a strong activation of GST, which is a key enzyme in the synthesis of glutathione, a strong antioxidant. GST participates in the detoxification of reactive oxygen species in order to avoid cell death induced by free radicals produced during the hypersensitive response of the susceptibility interaction (18), explaining the successful invasion of the pathogen into the cells of the host. In plants where GST is silenced, however, hypersensitivity response is active throughout the entire range of evaluated time points; and as a result, the plants display a strong resistance to the pathogen (Figure 2).
The dissection of new molecular mechanisms for pest resistance will allow the modification of highly susceptible plants into disease-resistant cultivars, taking advantage of the activation of more efficient resistance mechanisms. It will, additionally, provide genes that can be used as molecular markers within ongoing genetic crop improvement programs.
REFERENCES
1. Melton TA, Broadwell A, Wilson J. Disease management. In: Flue-Cured Tobacco Information. N. C. Coop. Ext. Serv. Bull, North Carolina State University, Raleigh, 2005, pp. 150-1.
2. Lucas G. B., Diseases of Tobacco, in: Biological Consulting Associates, Raleigh, NC, 1975.
3. Sullivan JM, Melton TA, Shew HD. Fitness of races 0 and 1 of Phytophthora parasitica var. nicotianae. Plant Dis 2005; 89:1220-8.
4. Todd FA. Flue Cured Tobacco- Producing a Healthy Crop, Tobacco Consultants, Wendell, NC, 1981.
5. Espino E, Rey X. Nuevas variedades de tabaco negro para cultivo bajo tela resistentes al moho azul (Peronospora tabacina). Agrotecnia Cuba 1987;19:247-60.
6. Borrás-Hidalgo O, Thomma BPHJ, Collazo C, Chacón O, Borroto CJ, Ayra C, et al. EIL2 transcription factor and glutathione synthetase are required for defense of tobacco against tobacco blue mold. Mol Plant Microbe Interact 2006; 19:399-406.
7. Collazo C, Ramos PL, Chacón O, Borroto CJ, López Y, et al. Phenotypical and molecular characterization of the TMTV - Nicotiana megalosiphon interaction. Physiol Mol Plant Pathol 2006; 67:231-6.
8. Race R, Fournier J, Esquerre-Tugaye MT. The incompatible interaction between Phytophthora parasitica var. nicotianae race 0 and tobacco is suppressed in transgenic plants expressing antisense lipoxygenase sequences. Proc Natl Acad Sci USA 1998; 95:6554-9.
9. Hugot K, Aime S, Conrod S, Poupet A, Galiana E. Developmental regulated mechanisms affect the ability of a fungal pathogen to infect and colonize tobacco leaves. Plant J 1999;20:163-70.
10. Hugot K, MP Riviere, Moreilhon C, Dayem MA, Cozzitorto J, Arbiol G, Barbry P, Weiss C, Galiana E. Coordinated Regulation of Genes for Secretion in Tobacco at Late Developmental Stages: Association with Resistance against Oomycetes. Plant Physiol 2004:134;858-70.
11. Lamb C, Dixon RA. The oxidative burst in plant disease resistance. Annu Rev Plant Physiol Plant Mol Biol 1997:48;251-75.
12. Sappl PG, Carroll AJ, Clifton R, Lister R, Whelan J, Millar AH, et al. The Arabidopsis glutathione transferase gene family displays complex stress regulation and cosilencing multiple genes results in altered metabolic sensitivity to oxidative stress. Plant J 2009;58:53-68.
13. Ezaki B, Katsuhara M, Kawamura M, Matsumoto H. Different mechanisms of four aluminum (Al)-resistant transgenes for Al toxicity in Arabidopsis. Plant Physiol 2001;127:918-27.
14. Dean JD, Goodwin PH, Hsiang T. Induction of glutathione S-transferase genes of Nicotiana benthamiana following infection by Colletotrichum destructivum and C. orbiculare and involvement of one in resistance. J Exp Bot 2005;56:1525-33.
15. Chacón O, Hernández I, Portieles R, López Y, Pujol M, Borrás-Hidalgo O. Identification of defense-related genes in tobacco responding to black shank disease. Plant Sci 2009;177:175-80.
16. Li BC, Bass WT, Cornelius PL. Resistance to Tobacco Black Shank in Nicotiana Species. Crop Sci 2006;46:554-60.
17. Csinos AS. Relationship of isolate origin to pathogenicity of race 0 and 1 of Phytophthora parasitica var. nicotianae on tobacco cultivars. Plant Dis 2005:89 332-7.
18. Hernández I, Chacón O, Rodríguez R, Portieles R, López Y, Pujol M, Borrás- Hidalgo O. Black shank resistant tobacco by silencing of glutathione S-transferase. Biochem Biophys Res Commun 2009:387; 300-4.
Orlando Borrás, Plant Division, Laboratory of Vegetal Functional Genomics, Center for Genetic Engineering and Biotechnology, CIGB Ave. 31 / 158 and 190, Cubanacán, Playa, PO Box 6162, Havana, Cuba. E-mail: orlando.borras@cigb.edu.cu

{kind=link}
{kind=link}
{kind=link}
{kind=link}