










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.28 no.2 La Habana abr.-jun. 2011
RESEARCH
Screening of protease inhibitory activity in extracts of five Ascidian species from Cuban coasts
Búsqueda de actividad inhibidora de proteasas en extractos de cinco especies de ascidias de las costas cubanas
Mey Ling Reytor1, Yamile González1, Isel Pascual1, Aida Hernández2, María Á Chávez1, Maday Alonso del Rivero1
2Instituto Nacional de Oceanología Calle 1era #18406, Playa, La Habana, Cuba.
ABSTRACT
One of the most promising strategies to counteract malaria is by controlling the infectivity of Plasmodium parasites with drugs aimed against their main targets. The participation of proteases (belonging to different classes) has been demonstrated in its infection mechanism, hence these proteins have become promising targets in treating this illness. This study involved the screening of crude and clarified extracts of five Cuban ascidian species for inhibitory activity of proteases belonging to different mechanistic classes. Ascidians are invertebrates that present many bioactive molecules, but, as yet, only a few protease inhibitors had been isolated from them. In this research we reported for the first time the presence of inhibitory activity of bovine pancreatic carboxypeptidase A and B, aminopeptidase N from porcine kidney and subtilisin A from Bacillus licheniformis in some ascidian extracts tested. The most promising extracts were selected in terms of percentage of inhibition and they were characterized in terms of the behaviour of the extract concentration against residual enzymatic activity; the determination of IC50 values and Specific Inhibitory Activity (SIA), and the evaluation of their proteolytic activity through zymographic assays.
Keywords: ascidians, proteolytic activity, inhibitors, malaria, proteases.
RESUMEN
Una de las estrategias más promisorias para contarrestar la malaria es controlar la infectividad del parásito Plasmodium, con fármacos dirigidos hacia sus blancos principales. Se ha demostrado que proteasas (pertenecientes a diferentes clases mecanísticas) participan en el mecanismo de infección, por lo que se han convertido en blancos promisorios para el tratamiento de la enfermedad. En este trabajo se investigaron extractos crudos y clarificados de cinco especies de ascidias cubanas en busca de actividad inhibidora de proteasas pertenecientes a diferentes clases mecanísticas. Las ascidias son animales invertebrados que presentan numerosas moléculas bioactivas, aunque solo unos pocos inhibidores de proteasas han sido aislados a partir de ellas. En esta investigación se informa por primera vez la presencia de actividad inhibidora de carboxipeptidasa A y B pancreática bovina, aminopeptidasa N de riñón porcino y subtilisina A de Bacillus licheniformis en algunos de los extractos de ascidias evaluados. Los extractos más promisorios se caracterizaron con relación al comportamiento de la concentración de extracto en función de la actividad enzimática residual, la determinación de los valores de IC
Palabras clave: ascidias, actividad proteolítica, inhibidores, malaria, proteasas.
INTRODUCTION
Malaria is a parasitic disease present in many regions of the world and produces approximately 1.5 - 2.7 million deaths each year (1). The resistance to current antimalarial drugs is spreading rapidly and the development of new vaccines is very difficult. The World Health Organization (WHO) has therefore argued the urgent need to develop new anti-malarial drugs aimed at new therapeutic targets. In this sense, the isolation of several proteases that play important roles during Plasmodium falciparum infection, the parasite that causes the most lethal form of Malaria, has been des-cribed. In its intra-erythrocyte state, P. falciparum degrades almost all of the haemoglobin for its growth and maturation. This process is carried out by proteases belonging to different mechanistic classes such as: aspartic proteases (plasmepsins) (2), cysteine proteases (falcipains) (3) and metallo-amino-peptidases (4). In addition, serine proteases (subtilases) (5) and genes encoding for metallo-carboxy-peptidases (6) have been found in the P. falciparum genome, and they have been related to its infection mechanisms. Certain inhibitors from these enzymes have been tested in in vitro experiments blocking P. falciparum growth and its expansion to other erythrocytes (7-10). Therefore, the search for inhibitors from these proteases has become a new strategy for developing new antimalarial drugs.
On the other hand, marine organisms are very important sources of leader compounds in current drug development. Among them, the ascidians (Phylum Chordata, Subphylum Urochordata) are animals found in every ocean, which are fixed onto rocks or shells (11). Several bioactive molecules have been isolated from ascidians, but only a very small fraction corresponds to protease inhibitors (12-14) and only two of them have been isolated and characterized until now. Taking into account, the importance of ascidians as a source of bioactive compounds, as well as the need to find active inhibitors against Plasmodium proteases, we carried out this study where we screened extracts of five Cuban ascidian species for protease inhibitors, against model proteases belonging to the same classes as those found in the parasite, such as: bovine pancreatic carboxypeptidase A (CPA) and B (CPB) (metallo-carboxy-peptidases), pancreatic trypsin and Bacillus licheniformis subtilisin A (serine proteases), porcine pepsin (aspartic protease), Carica papaya papain (cysteine protease) and porcine kidney amino-peptidase N (APN) (metallo-amino-peptidase). This is a preliminary characterization of the inhibitory activity shown by these ascidian extracts that are useful for the future selection and purification of the most promissory extracts.
MATERIALS AND METHODS
Materials
Enzymes: CPA (EC 3.4.17.1), CPB (EC 3.4.17.2), trypsin from bovine pancreas type I (EC 3.4.21.4), porcine pepsin (EC.3.4.23.1) and subtilisin A from B. licheniformis (EC 3.4.21.62) were purchased from Sigma Chemical Company (EUA). Papain from C. papaya (EC 3.4.22.2) was purchased from Calbiochem-Novabiochem (EUA). APN was partially purified from porcine kidney following the procedure described by Golich and colleagues (15). A partially purified mixture of aminopeptidases, containing APN, aminopeptidase A (APA) and aminopeptidase B (APB) was obtained, which was termed APN. Synthetic substrates: N-(4-Metoxiphenylazoformyl)-L-phenylalanine (AAFP), Metoxyphenylazoformyl)-L-arginine (AAFR)-L-arginine (AAFR), Benzoyl-arginine-p-nitro-aniline-HCl (BAPA), t-butyloxycarbonyl-glycyl-glycyl-leucil-p-nitroaniline (Boc-Gly-Gly-Leu-pNA), Leucine-p-nitroaniline (Leu-pNA) and leucine-serine-nitrophenilalanine-norleucine-leucine-methyl ester (Leu-Ser-Phe(NO2)-Nle-Ala-Leu-OMe) were purchased from the Bachem Company (Germany).
Preparation and clarification of ascidian crude extracts
Five ascidian species were studied: Ascidia nigra, A. sydneiensis, Ecteinascidia turbinata, Diplosoma listerianum and Poticlinum constellatum. The specimens were collected from the Northwestern coast of Havana City (Cuba). To obtain the crude extract, whole animals were homogenized in distilled water at a 1:2 proportion (w/v) at 4 oC using a blender (10 sec for 5 repeated cycles). Then, the extracts were centrifuged at 10 000 x g for 30 min at 4 oC and the supernatant was filtered on fiberglass (16).
Clarified extracts were obtained by heating or with the acid treatment of crude extracts. Heating was done for 20 min at 60 oC followed by extract centrifugation at 12 100 x g for 30 min at 4 oC. Alternatively, extracts were treated with 2.5% trichloroacetic acid (TCA) and incubated for 1 h at room temperature. Then, they were centrifuged at 17 400 x g for 30 min at 4 oC. The supernatant was adjusted to pH 7.0 with 1M NaOH and extensively dialyzed against distilled water (1:100, supernatant/water) using membranes with 500 Da of the cutoff (Spectra/Por®, USA).
Determination of protein concentration
Protein concentration of all extracts was determined by absorbance at 280 nm (Ultrospect 4000 spectrephotometer) assuming a x280nm0.1% = 1 (17) and enzyme concentration was calculated using their extinction coefficient (18). In the case of APN, total protein concentration was determined using the Bradford method (19).
Inhibitory activity of the extracts
The enzymatic activity for CPA (9.0 x 10-7 M) and CPB (9.0 x 10-7 M) was assayed using a specific substrate for each enzyme: AAFP (0.1 mM, 1 KM (Michaelis-Menten constant)) (20) and AAFR (0.1 mM, 1 KM) (21), respectively. Trypsin (3.1 x 10-7M) and papain (3.0 x 10-8 M) were tested using BAPA (1 mM, 1 KM) as the substrate (22-23). The enzymatic activity of subtilisin A (3.0 x 10-7 M) was determined using Boc-Gly-Gly-Leu-pNA (1 mM, 1 KM) (24) and for pepsin (7.1 x 10-9 M) it was monitored using Leu-Ser-Phe(NO2)-Nle-Ala-Leu-OMe (0.22 mM, 11.3 KM) as the substrate (25). APN (0.2 mg/mL) was evaluated against Leu-pNA (0.3 mM), a specific substrate for this enzyme and not for APA and APB (26). In all cases the enzyme and substrate concentrations are referred to the assay.
The enzymatic assays were carried out in an Ultrospec 4000 spectrophotometer. The inhibition assays included a 10 min pre-incubation of the sample with the enzymes on the activity buffer at 25 oC or 37 oC before the substrate was added (temperature and buffer used depended on the enzyme). Inhibition was detected through the decrease in residual activity, which is the ratio of vi/vo, where vi and vo are the initial rates of the reaction in the presence and absence of the inhibitor, respectively. The behavior of residual activity for each extract was evaluated through the increase in extract concentration. To calculate the IC50 values (concentration of the inhibitor producing 50% inhibition), the experimental data were adjusted to the IC50 equation using the GRAFIT program (version 3.01).
Additionally, in all cases, the percentage of inhibition (%I) and the initial inhibitor ([Io]) and enzyme ([Eo]) concentration ratio ([Io]/[Eo]) were calculated in the assay. For these determinations we used the extract concentration producing the highest inhibition (lower residual activity values) and we assumed total initial protein concentration as [Io]. We considered the extracts that could produce more than 65% inhibition with [Io]/[Eo] values lower than 50 as the positive selection criteria for a more thorough characterization.
Also, the proteolytic activity of the most promissory extracts was evaluated in order to avoid false positive inhibition due to proteolytic enzymes that could hydrolyze the enzyme used in the inhibitory assay. Enzymatic activity against the different chromogenic substrates (specific for each type of enzyme) was measured under the same conditions described above for each enzyme, but without the enzyme used as the control in the inhibitory assay.
Zymography
Proteolytic activity of the extracts was also evaluated through zymographic assays using gelatine (16%) as the substrate included in 12.5% polyacrylamide gel (27). The crude and clarified extracts were run through the gel. Gels were washed with Triton X100 (twice) and incubated overnight with 20 mM Tris-HCl, 0.5 M NaCl buffer, pH 8.0. Subsequently, they were stained and de-stained according to the Lantz and Ciborowski procedure (27).
RESULTS
Screening of protease inhibitory activity
The inhibitory activity was evaluated in the crude and clarified extracts. The crude extracts did not show inhibitory activity against proteases tested, with the exception of the D. listerianum extract, which was active against APN, reaching 80.3% of inhibition (result not shown). Hence, we only show the results obtained with clarified extracts because most of them were able to inhibit some of the enzymes tested in this screening.
Regarding the trypsin and papain enzymes,s, only the heated extract of D. listerianum inhibited trypsin and the TCA treated extract of E. turbinata inhibited papain with the inhibitory activity percentage above 65%.However the [Io]/[Eo] ratio was very high (taking into account our selection criteria; table 1). No extract showed inhibitory activity for pepsin (Table 1). For this reason, we did not continue working with these three enzymes and focused only on the results related to the inhibition of the enzymes CPA, CPB, APN and subtilisin by certain clarified extracts.
{kind=link}
In the case of A. sydneiensis and P. constellatum, only TCA treated extracts showed inhibition, in particular against CPA (Table 1). The Specific Inhibitory Activity (SIA) calculated for them was 1499 U/mg and 725 U/mg, respectively (with [Io]/[Eo] ratio of 2.5 and 1.9) and the former had the largest SIA value of all in the screening. For these extracts, the effect of their concentration against CPA residual activity presented a concave behaviour, which is characteristic of reversible inhibition, with IC50 values of 13 mg/mL and 38 mg/mL, respectively (Figure 1).
{kind=link}
On the other hand, clarified extracts of D. listerianum showed a broad specificity against metallo and serine proteases (Table 1). They were able to reach a high inhibition of CPA, CPB, APN and subtilisin showing a concave behaviour for these enzymes (Figure 2). The TCA treated extract inhibited CPA and APN with IC50 values of 24 mg/mL and 340 mg/mL, whereas the heated extract inhibited APN and CPB, showing IC50 values of 480 and 426 mg/mL, respectively. The heated extract also inhibited subtilisin, but produced total inhibition with an IC50 value of 40 mg/mL. The SIA of the heated extract against APN, CPB and subtilisin was 81 U/mg, 41 U/mg and 75 U/mg, respectively; while the SIA obtained for the TCA treated extract against APN and CPA were 118 U/mg and 435 U/mg, being the latter the largest value obtained for this ascidian specie.
{kind=link}
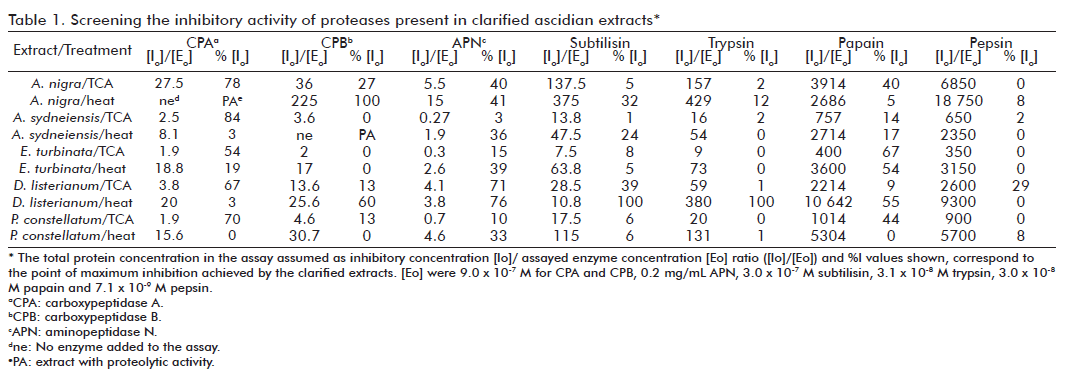
On the other hand, interesting results related to the metallo-carboxy-peptidases tested in this screening were obtained with clarified A. nigra extracts. The TCA treated extract only showed CPA inhibitory activity with a concave behaviour when plotting extract concentration against residual activity, an IC50 value of 168 mg/mL and a SIA value of 197 U/mg (Table 1; Figure 3A). Nevertheless, the heated extract was not able to inhibit CPA; in contrast, it has CPA-like enzymatic activity, because it is able to hydrolyze AAFP, a specific substrate for CPA. Moreover, the heated extract showed CPB inhibitory activity and was able to reach 100% inhibition but with a [Io]/[Eo] = 225 (Table 1).
{kind=link}
Proteolytic activity and zymography of clarified extracts
In order to evaluate the presence of false positive inhibitors in the extract mentioned above due to proteases (which could degrade the enzyme used as the control), we evaluated the proteolytic activity of all crude and clarified extracts through colorimetric assays and zymography described in the experimental section. Only the specie A. nigra (both crude and heated extracts) showed enzymatic activity against chromogenic substrates, in particular against AAFP, a typical substrate for CPA. However, this activity was not found when the extract was treated with TCA. Figure 3B shows the linear relationship between the initial rate of the enzymatic reaction and extract concentration obtained for the heated extract. This reaction was inhibited by the potato carboxypeptidase inhibitor, an inhibitor of a metallo-carboxy-peptidase belonging to the M14A subfamily (data not shown).
On the other hand, as a result of zymographic assays we found that crude and heated extracts of P. constellatum, A. sydneiensis and A. nigra displayed clear bands in the gels, which represent gelatine degradation by the presence of proteolytic enzymes in the extracts. In contrast, when the extract was treated with TCA the proteolytic activity disappeared and no clear bands were observed (Figure 4A-C). In the case of the D. listerianum crude extract, two clear bands were observed (with different molecular weights), and only one band disappeared with each treatment.
{kind=link}
DISCUSSION
One of the most important strategies to counteract malaria is to design a drug composed of natural inhibitors against the main protease parasite targets. In this paper we used model proteases to screen ascidian extracts for new natural inhibitors that in the future could be purified and characterized for this purpose.
Ascidians constitute a very important group of organisms for the isolation of bioactive molecules, being the most promising of them the drug Yondelis, isolated from E. turbinata and used against different types of cancer (28). However, only a few protease inhibitors have been isolated from these organisms. The first one was an inhibitor of trypsin (enzyme belonging to PA clan, S1 family) isolated from the Halocynthia roretzi (14). The second were two inhibitors active against trypsin and elastase, respectively, isolated from Polyandrocarpa misakiensis (13). The third is an inhibitor of matrix metalloprotease 2 (MA clan, M10 family), isolated from a tunicate of the Polyclinidae family (12).
The ability of marine invertebrates to inhibit enzymess belonging to the M1 family of the MA clan was described by our group in the case of HcPI, an inhibitor isolated from Hermodice carunculata that is able to inhibit pyroglutamyl aminopeptidase II (29). Nevertheless, in this study we report for the first time the inhibition of APN (MA Clan, M1 family) by extracts of D. listerianum. It is important to emphasize that the molecules responsible for APN inhibition in this species are stable under both clarification conditions used in this work.
In P. falciparum a metallo-amino-peptidase (PfAM1) belonging to the same family of APN was found, which had broad substrate specificity and was involved in haemoglobin degradation (30). This fact suggests that PfAM1 constitutes a crucial target for the inhibition of parasite growth and development (31). It is possible that APN inhibitors also could be able to inhibit PfAM1, which could be very promising in obtaining new antimalarial drugs (31).
Also for D. listerianum we reported for the first time the inhibitory activity of CPA, CPB (MC clan, M14 family) and subtilisin (SB clan, S8 family). Recently it was reported that the P. falciparum genome contains a gene of a metallo-carboxy-peptidase like enzyme belonging to the Nna1 family (6). Its recombinant Caenorhabditis elegans homologous presented broad substrate specificity and is inhibited by inhibitors of metallo enzymes (6). These authors suggest that these types of enzymes could have tubulyn carboxypeptidase activity and could therefore be involved in the infection mechanism of the parasite (6). In addition, the expression of two midgut carboxypeptidase B gene (cpbAg1 and cpbAg2) in Anopheles gambiae has been found to be up-regulated by P. falciparum infection and the addition of antibodies against CPBAg1 to a P. falciparum-containing blood meal, inhibited CPB activity and blocked parasite development (32). Therefore, the inhibition of both CPA and CPB like enzymes could result in a promissory strategy for developing drugs and transmission-blocking vaccines against the parasite (6, 32).
TCA treated extracts of A. Nigra, A. sydneiensis, P. constellatum and D. listerianum inhibit CPA in a reversible way, taking into account the behaviour of the extract concentration against CPA residual activity. For CPA, the analysis of IC50 and SIA values suggests that A. sydneiensis represents the most efficient extract in CPA inhibition. The biological activity (inhibitory or proteolytic) of A. nigra and A. sydneiensis extracts found in this work could be related to the recently reported inhibition of P. falciparum growth by these extracts (33).