










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.28 no.2 La Habana abr.-jun. 2011
RESEARCH
Association analysis of COMT polymorphisms and Schizophrenia in a Cuban family sample. Preliminary results
Análisis de asociación de polimorfismos del gen COMT y la esquizofrenia en una muestra familiar cubana. Resultados preliminares
Hanlet Camacho1, Marcelo Nazabal1, Mayelín Domínguez2, Raúl Mendoza2, Lester Leal1, Juan Roca1, Alexander García2, Jesús Benítez1, María E Fernández de Cossío1, Adelaida Villarreal1, Alberto Cintado1, Annia Ferrer1, Marta Dueñas1, Tamara Díaz1, Racmar Casalvilla1, Migdyrai Martín2, Lidia I Novoa1
1Centro de Ingeniería Genética y Biotecnología, CIGB Ave. 31 e/ 158 y 190, PO Box 6162, Cubanacán, Playa, La Habana, Cuba.
2Cuban Neurosciences Center, CNEURO.
ABSTRACT
Schizophrenia is a complex disease affecting as much as 1% of the global population, constituting the target of considerable effort to identify disease susceptibility genes. Several lines of evidence point to the catechol-O-methyltransferase (COMT) gene as a schizophrenia susceptibility candidate, not only because it encodes a key dopamine catabolic enzyme but also because it maps to the velocardiofacial syndrome region of chromosome 22q11, which has long been associated with predisposition to the disease. Several case-control and family-based studies have been conducted to examine the possible association of COMT with this disorder; however, these studies have produced conflicting results. To further assess the genetic contribution of COMT variants to schizophrenia susceptibility, three single-nucleotide polymorphisms (rs2075507, rs4680 and rs362204) were investigated in a sample of 74 family trios from the Cuban population. Restriction fragment length polymorphism was used to identify the allelic variants, employing statistical tools based on transmission disequilibrium tests to find possible associations. In this study, the first of its type performed in the Cuban population, we found an association of rs2075507 and rs362204 at allelic levels with a p < 0.05; also finding an association of haplotype 1-2-1 with the disease.
Keywords: candidate genes; single nucleotide polymorphism (SNP); family-based association study, family trios.
RESUMEN
La esquizofrenia es una enfermedad común y compleja que afecta cerca del 1% de la población mundial, por lo que grandes esfuerzos se llevan a cabo para identificar los genes de susceptibilidad a padecer la misma. Varias líneas de evidencias implican al gen que codifica para la catecol-O-metiltransferasa (COMT), como uno de los genes candidatos que confieren susceptibilidad, no solo porque codifica para una enzima clave en el proceso de degradación de la dopamina, sino también porque está ubicado en la región cromosómica 22q11 implicada en el síndrome de velocardio facial, el cual ha sido ampliamente asociado con la predisposición a padecer la enfermedad. Varios estudios de caso-control y basados en familias han sido realizados para determinar la posible asociación de este gen con la esquizofrenia, pero desafortunadamente los resultados de los mismos han sido contradictorios. Para comprobar la relación del gen COMT con la esquizofrenia, 3 polimorfismos de simples nucleótidos presentes en el gen (rs2075507, rs4680, rs362204), fueron estudiados en una muestra de 74 tríos familiares de la población cubana. Se empleo la técnica de polimorfismos por fragmentos de restricción para el genotipaje y la estadística basada en las pruebas de desequilibrio en la transmisión de alelos para la búsqueda de asociación. En este estudio, el primero de su tipo reportado en la población cubana, encontramos asociación de los marcadores rs2075507 y rs362204 a nivel alélico con un valor de p < 0.05 y de igual forma encontramos una asociación a nivel haplotípico del haplotipo 1-2-1 con la entidad patológica.
Palabras clave: Genes candidatos, polimorfismos de simple nucleótidos, estudios de asociación basados en familias, tríos familiares.
INTRODUCTION
Schizophrenia is a severe mental illness with a global incidence close to 1%. Convergent data have shown that both genetic and environmental factors are involved in the development of this disorder (1, 2).
Several key neurotransmitters, especially dopamine and serotonin, have become the focal point of schizophrenia research along the quest to understand the causes of the disease. This situation arises, to a large extent, from the fact that most drugs used for the treatment of schizophrenia interfere with the function of a neurotransmitter(s) or its receptor(s) (2). Neurotransmitters have been implicated in the etiology of schizophrenia since the time the dopamine hypothesis was conceived, postulating that the symptoms of the disease were caused by excess dopaminergic neurotransmission, particularly in mesolimbic and striatal brain regions, that led to positive symptoms and dopaminergic deficits in prefrontal brain regions that, in turn, are responsible for the negative symptoms (3).
One of the most studied genetic loci in the context of the dopaminergic hypothesis has been the gene coding for catechol-O-methyltransferase (COMT), the enzyme catalyzing the first step in one of the major degradative pathways of the catecholamine neurotransmitters, dopamine, epinephrine, and norepinephrine (4, 5). Dopamine is degraded mainly by monoamine oxidase in most brain regions, aided by COMT and phenosulfotransferase activities. COMT, however, becomes much more preponderant in the prefrontal cortex, where it is responsible for more than 60% of dopamine degradation (6-8). There are two major isoforms of the COMT protein; the soluble cytoplasmic type (S-COMT), transcribed by promoter p1 and common in the peripheral nervous system, and the membrane-binding type (MB-COMT), transcribed by promoter p2 and predominant in the central nervous system (9).
The COMT gene has constituted an intriguing target for neuropsychiatric genetic studies, not only for its pharmacological properties but for its genomic location on 22q11. The deletion of the region containing COMT (22q11) is associated with velocardiofacial syndrome, and since patients suffering from this syndrome have an elevated rate of psychosis, this region has consequently also been tagged as a possible “schizophrenia locus” by several linkage and association studies (10, 11).
In our study, the first of its type conducted in our country, we genotyped three restriction site polymorphisms (RSP) reported by DeMille (12) in the COMT gene: the P2 promoter region HindIII RSP at –1217 (rs2075507), a useful marker located in the estrogen-sensitive portion of COMT (13); the exon 4 Nla III RSP (rs4680), a functional variation that has been studied in a large number of populations (14); and the single-base deletion/insertion immediately 3′ to the stop codon in exon 6, which is also detected as the BglI RSP (rs362204) (15).
This genetic family study was conducted as previously reported for other populations (16, 17) and forms part of ongoing research to examine the association of these polymorphisms to schizophrenia in our population sample. In addition, it is intended to establish a methodological foundation for examining other candidate genes for this disorder in the Cuban population. The analysis will be relevant to further understanding of the genetic basis of this disease in our sample, finding differences or commonalities with populations in other parts of the world.
MATERIALS AND METHODS
Family selection
Parent-offspring trios consisting of well characterized paranoid schizophrenic probands and their biological parents were selected in Havana city. Patients and parents were contacted through the Mental Health Community Centers of each municipality, using a group of psychiatrists to confirm, in every case, the diagnosis of Paranoid Schizophrenia following the criteria of the Diagnostic and Statistical Manual of Mental Disorders (DSM IV) (18) and the International Classification of Illnesses (CIE-10) (19). In addition, the Scale for the Assessment of Positive (SAPS) and Negative (SANS) Symptoms was used to characterize the predominant clinical symptoms (20, 21). All patients had a positive family history defined as first- or second-degree relative with schizophrenia or paranoid personality disorder (multiplex family).
All interviews and samples were collected with the informed consent of the participants and approved by the appropriate institutional review boards.
Collection and processing of blood samples
Blood samples were collected in two 8 mL vacutainers per sample with sodium citrate as anticoagulant (Beckton and Dickinson, Franklin Lakes, NJ, 07417-1885). Genomic DNA was extracted from whole blood as starting material using the Promega Wizard DNA Purification Kit (Promega Corporation, Madison, WI, USA).
DNA integrity was analyzed in 0.8% agarose gel electrophoresis, visualizing the fluorescence of ethidium bromide under 320 nm ultraviolet light. The concentration of the genomic DNA was determined in an Ultrospec 2000 Pharmacia Biotec spectrophotometer.
Genotyping
Three polymorphisms of the COMT gene were genotyped using the primer pairs previously designed by DeMille et al. 2002, taking into account the characteristics described for this RSP at the Allele Frequency Database (ALFRED) site (12, 22).
Polymerase chain reaction (PCR) amplifications contained 1U GoTaq DNA Polymerase (Promega), GoTaq Buffer, 1.5 mM MgCl2, 50 ng of each primer, 200 ng genomic DNA, 0.2 mM dNTPs, in 20 µL of final volume, and were subjected to the following cycling protocol: 95 °C for 10 min; then 94 °C for 30 s, 54 °C for rs2075507 or 60 °C for rs4680 and rs362204 for 30 s, and 72 °C for 30 s, for 35 cycles; followed by 72 °C for 10 min.
The examination of restriction fragment length polymorphisms (RFLP) at each of three polymorphic sites was performed according to the manufacturer’s protocol for each enzyme, analyzing the resulting fragments in agarose gels visualized with ethidium bromide.
The size of the bands obtained by PCR and RFLP was determined with Kodac 1D Image Analysis Software Version 3.6, comparing the results with the expected values according to the available sequences of the COMT gene in the databases of the National Center for Biotechnology Information (NCBI, USA) (23).
The expected allelic variants were: 1 = A (absence of restriction site) and 2 = G (presence of restriction site) for rs2075507, 1 = G (absence of restriction site) and 2 = A (presence of restriction site) for rs4680 and 1 = C insertion (absence of restriction site) and 2 = No C insertion (presence of restriction site) for rs362204.
Statistical analysis
Hardy-Weinberg ratios were calculated for each of the three polymorphic sites through chi-squared tests to determine if they were in equilibrium.
Haplotype frequency estimates were calculated using HAPLO (24), which implements the expectation maximization (EM) algorithm.
In our study, the first of its type conducted in our country, we used a family trios-based association design (25, 26).
The family trios association was analyzed with the Family-Based Association Test (FBAT), introduced by Rabinowitz and Laird (26), and Laird et al. (25); based on the transmission disequilibrium test (TDT) (25-27). FBAT analysis was applied under two main hypotheses (Ho: Non-association, non-linkage and Ho: Non-association in the presence of linkage), at allelic, genotype and haplotype levels, always assuming an incidence of 1% for the disease in the population.
A precise correction method for multiple comparisons was applied using the formula alpha = 1-0.95 1/N where N is the number of comparisons, one or more of which shows a significant result (28).
RESULTS
We selected a total of 74 family trios (222 samples) after interviewing approximately 700 families, for an 89.45% rejection rate. The DNA obtained from the 222 samples matched the expected quality according to the kit manufacturer.
Genotyping
Table 1 contains the results obtained after genotyping the three selected polymorphisms in the 74 family trios. The genotype proportions did not deviated significantly from those expected under conditions of HW equilibrium p > 0.05 (p = 0.664 for rs2075507; p = 0.4715 for rs4680 and p = 0.0993 for rs362204), and allele transmission from parents to descendents followed Mendel’s laws in every tested family.
The frequencies of the eight possible haplotypes that could be derived from the combination of all three polymorphisms, determined using the EM method, appear in table 2.
Family-Based Association Test
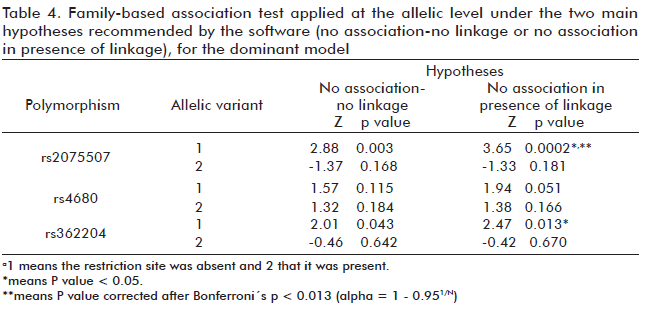
Under the two main null hypotheses (no association-no linkage and no association in presence of linkage), FBAT finds a positive allelic level association of rs2075507 and rs362204 with the disease (p < 0.05; tables 3 and 4). In consequence, the results were similar under the additive and dominant model analyses, although statistical significance was lost for marker rs362204 after applying the Bonferroni correction.
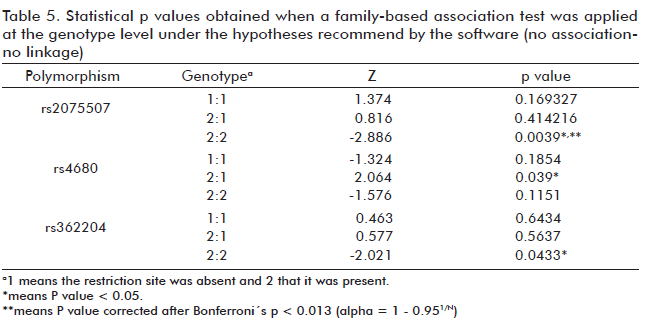
The no association-no linkage hypothesis was used to analyze association at the genotype level, obtaining a possible association of different genotypes with different effects on the disease. Genotype 2-2 for markers rs2075507 and rs362204 has shown a positive association, which was different from the negative association displayed by genotype 1-2 of marker rs4680 (Table 5). The only association still showing statistical significance after the application of Bonferroni correction to this analysis was that of genotype 2-2 for rs2075507.
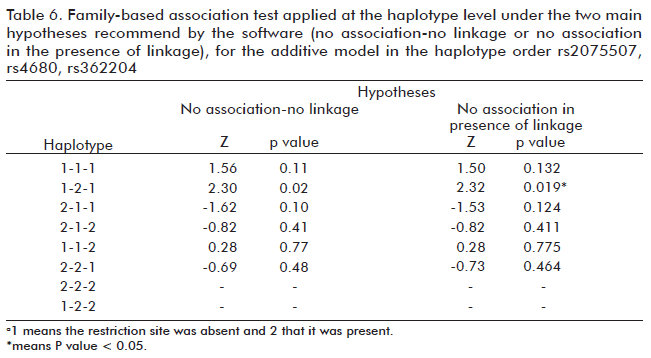
Haplotype analysis was conducted under the two hypotheses mentioned above, and the positive associations obtained for the 1-2-1 haplotype were equal under the additive model for two hypotheses (Table 6). After the corrections for multiplex test, these associations at the haplotype level were lost.
DISCUSSION
Different genetic studies have been carried out with the aim at finding genes associated to schizophrenia. These are mostly linkage analyses, case-control association studies and linkage disequilibrium studies (29).
Family trio sampling was applied in this case to avoid the effect of co-founding events, the admixture phenomenon and population stratification, according to Rabinowitz and Laird (26) and Laird et al. (25). The high percentage of rejected families in our case is due to failure to comply with the inclusion criteria described in Materials and methods.
Family-based association test at allelic level
Statistical variable Z provides a measure of the association status of the marker variant. If Z > 0 with a significant p value, the possible association will then be positive, implying that the presence of the marker variant could confer susceptibility to the disease. If, on the other hand, Z < 0, then the possible association will be negative, meaning that the presence of the marker could protect against the disease.
When performing an allelic-level analysis under the hypothesis No association-No linkage, we found a statistically significant linkage (p < 0.05) with the disease for allele A in the rs2075507 polymorphism and non-insertion of C in the rs362204 polymorphism (Tables 3 and 4). Then, under the hypothesis No association in presence of linkage, we confirmed a significant difference in the transmission of allele A for the rs2075507 polymorphism and not insertion of C allele for the rs362204 polymorphism, with p < 0.05. This represents an association with schizophrenia for these two alleles in our sample. After the application of the Bonferroni correction (p < 0.013), however, statistical significance was lost for rs362204. This may imply that statistical significance may be regained with larger sample sizes or, alternatively, that there is simply no association in this case.
The associations obtained for allele A in the case of rs2075507 were maintained after the corrections for multiplex tests. In this case, according to the Z value (Z > 0), the presence of this allele increases susceptibility to the disease, a finding that matches those of Palmatier et al. (14) when analyzing these polymorphisms as part of a haplotypic study in 38 populations distributed worldwide (14). Our finding also concurs with the results of Chen et al. during the study of Irish families (30) and differs with family based association tests results obtained by Fan et al. (16), and Kremer et al. (31) when analyzed Chinese and Palestinian Arabs populations, respectively.
The association effect of these polymorphisms may have a direct cause in variations in gene transcription, protein synthesis or biological activity mediated by the polymorphism itself, or may actually be traced to the effects of other markers in linkage disequilibrium with the target polymorphism, as reported e.g. by Shifman et al. (32) when studying a population of Ashkenazi Jews.
On the other hand, the location of the rs2075507 polymorphism is very close to the estrogen binding site, and an identical change in that position has been shown to alter the estrogenic regulation of gene transcription at this locus (13).
FBAT at genotype level
At the genotype level only non-association, non-linkage can be used as null hypothesis by the software. In this case, we observed a linkage and possible association of homozygous genotypes for the presence of the restriction site in polymorphisms rs2075507 and rs362204 (p < 0.05), with Z < 0. This would indicate that these genotypes, if indeed associated, would be protective (Table 5).
The results for these two polymorphisms at the genotype level were congruent with those at the allelic level. The presence of genotype 2-2 for both polymorphisms had shown a possible negative association with the disease. Additionally, the presence of allelic variant 1 at the allelic level in our study showed a possible positive association with the illness.
We also found a positive linkage and a possible association among a heterozygous genotype for the rs4680 marker and the disease during the trio study (p < 0.05). The possible linkage or associations, however, were lost for genotypes 2-2 and 1-2 of rs362204 and rs4680 respectively after the application of the Bonferroni correction (p < 0.013).
In the future, it would be interesting to confirm the association of the rs4680 polymorphism for our population, since other published studies have already shown an association of this polymorphism with the disorder (33, 34).
FBAT at haplotype level
When we analyzed the possible association of the haplotypes formed by the three markers, we found, under the first null hypothesis, a positive (Z > 0) linkage for haplotype 1-2-1 (Table 6). Although the possible association of this haplotype was confirmed under the second hypothesis, the associations were lost in both cases after applying the corrections for multiplex tests.
The results achieved before the corrections confirm those obtained at the allelic level and coincide with those reported by Palmatier et al. (14), reinforcing the theory of the effect of the three polymorphisms on the susceptibility to the disease, whether by themselves of through some other polymorphism with which they are in linkage disequilibrium (14,32). Such a polymorphism might be, for instance, the 67 bp insertion-deletion polymorphism or the tetranucleotide repeats (STR) located to the right of marker rs2075507 within the COMT coding sequence. While these markers do not constitute single base-pair changes (SNP) but longer sequence variations (Macro satellites), their study is becoming relevant due to the statistical power conferred to haplotype analysis by the combination of SNP with STR, according to the 2006 report by Komura et al. (35).
Despite the sample size of our study, we have been able to replicate some of the findings made by other association studies examining the association of the COMT gene with schizophrenia in several populations worldwide. We found possible associations for the rs2075507 and rs362204 markers that would confer susceptibility to the disease at the allelic level, and also found a possible association of the haplotype rs2075507-rs4680-rs362204 with the pathology.
Our results provide a preliminary characterization of the association of these markers with schizophrenia in a Cuban sample, in addition validate the employed sampling and genotyping procedures with a focus on their application for futures studies.
REFERENCES
1. Gejman PV, Sanders AR, Duan J. The role of genetics in the etiology of schizophrenia. Psychiatr Clin North Am. 2010; 33(1):35-66.
2. Shin JK, Malone DT, Crosby IT, Capuano B. Schizophrenia: a systematic review of the disease state, current therapeutics and their molecular mechanisms of action. Curr Med Chem. 2011;18(9):1380-404.
3. Miyamoto S, LaMantia AS, Duncan GE, Sullivan P, Gilmore JH, Lieberman JA, et al. Recent advances in the neurobiology of schizophrenia. Mol Interv. 2003;3:27-39.
4. Sagud M, Mück-Seler D, Mihaljević-Peles A, Vuksan-Cusa B, Zivković M, Jakovljević M, et al. Catechol-O-methyl transferase and schizophrenia. Psychiatr Danub. 2010;22(2):270-4.
5. Kvetnansky R, Sabban EL, Palkovits M. Catecholaminergic systems in stress: structural and molecular genetic approaches. Physiol Rev. 2009;89(2):535-606.
6. Volk DW, Lewis DA. Prefrontal cortical circuits in schizophrenia. Curr Top Behav Neurosci. 2010;4:485-508.
7. Murray RM, Lappin J, Di Forti M. Schizophrenia: from developmental deviance to dopamine dysregulation. Eur Neuropsychopharmacol. 2008;18 Suppl 3:S129-34.
8. Remington G. Alterations of dopamine and serotonin transmission in schizophrenia. Prog Brain Res. 2008;172:117-40.
9. Lundström K, Tenhunen J, Tilgmann C, Karhunen T, Panula P, Ulmanen I. Cloning, expression and structure of catechol-O-methyltransferase. Biochim Biophys Acta. 1995;1251(1):1-10.
10. Lajiness-O’Neill R, Beaulieu I, Asamoah A, Titus JB, Bawle E, Ahmad S, et al. The neuropsychological phenotype of velocardiofacial syndrome (VCFS): relationship to psychopathology. Arch Clin Neuropsychol. 2006;21(2):175-84.
11. Murphy KC, Owen MJ. Velo-cardio-facial syndrome: a model for understanding the genetics and pathogenesis of schizophrenia. Br J Psychiatry. 2001;179: 397-402.
12. DeMille MM, Kidd JR, Ruggeri V, Palmatier MA, Goldman D, Odunsi A, et al. Population variation in linkage disequilibrium across the COMT gene considering promoter region and coding region variation. Hum Genet. 2002;111(6):521-37.
13. Xie T, Ho SL, Ramsden D. Characterization and implications of estrogenic down-regulation of human catechol-O-methyltransferase gene transcription. Mol Pharmacol. 1999;56(1):31-8.
14. Palmatier MA, Pakstis AJ, Speed W, Paschou P, Goldman D, Odunsi A, et al. COMT haplotypes suggest P2 promoter region relevance for schizophrenia. Mol Psychiatry. 2004;9(9):859-70.
15. Chen CH, Lee YR, Liu MY, Wei FC, Koong FJ, Hwu HG, et al. Identification of a BglI polymorphism of catechol-O-methyltransferase (COMT) gene, and association study with schizophrenia. Am J Med Genet. 1996;67(6):556-9.
16. Fan JB, Chen WY, Tang JX, Li S, Gu NF, Feng GY, et al. Family-based association studies of COMT gene polymorphisms and schizophrenia in the Chinese population. Mol Psychiatry. 2002;7(5):446-7.
17. Paweł K, Hauser J, Skibinska M, Szczepankiewicz A, Dmitrzak-Weglarz M, Gorzkowska K, et al. Family based association study of DRD1, DRD2, DRD3, DRD4, DAT, COMT gene polymorphism in schizophrenia. Psychiatr Pol. 2010;44(3):405-13. Polish.
18. Fuller AK. Diagnostic and Statistical Manual of Mental Disorders; DSM-IV Sourcebook; Study Guide to DSM-IV; DSM-IV Casebook: A Learning Companion to the Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition; The Clinical Interview: Using DSM-IV. N Engl J Med. 1994; 331:1163-6.
19. Organización Mundial de la Salud. Trastornos mentales y del comportamiento. Clasificación Internacional de Enfermedades. 10ma ed. Madrid: Meditor; 1993.
20. Andreasen NC. Improvement of negative symptoms: Concepts, definition and assessment. Int Clin Psychopharmacol. 1997;12 Suppl 2:S7-10.
21. Jablensky A. The diagnostic concept of schizophrenia: its history, evolution, and future prospects. Dialogues Clin Neurosci. 2010;12(3):271-87.
22. Rajeevan H, Osier MV, Cheung KH, Deng H, Druskin L, Heinzen R, et al. ALFRED: the ALelle FREquency Database. Nucleic Acids Res. 2003;31:270-1.
23. Pruitt KD, Tatusova T, Maglott DR. NCBI Reference Sequence (RefSeq): a curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 2005;33(Database issue):D501-4.
24. Hawley ME, Kidd KK. HAPLO: a program using the EM algorithm to estimate the frequencies of multi-site haplotypes. J Hered. 1995;86(5):409-11.
25. Laird NM, Horvath S, Xu X. Implementing a unified approach to family-based tests of association. Genet Epidemiol. 2000;19 Suppl 1:S36-42.
26. Rabinowitz D, Laird N. A unified approach to adjusting association tests for population admixture with arbitrary pedigree structure and arbitrary missing marker information. Hum Hered. 2000;50(4):211-23.
27. Spielman RS, McGinnis RE, Ewens WJ. Transmission test for linkage disequilibrium: the insulin gene region and insulin-dependent diabetes mellitus (IDDM). Am J Hum Genet. 1993;52(3):506-16.
28. Sham PC. Statistical methods in psychiatric genetics. Stat Methods Med Res. 1998;7(3):279-300.
29. Duan J, Sanders AR, Gejman PV. Genome-wide approaches to schizophrenia. Brain Res Bull. 2010;83(3-4):93-102.
30. Chen X, Wang X, O’Neill AF, Walsh D, Kendler KS. Variants in the catechol-o-methyltransferase (COMT) gene are associated with schizophrenia in Irish high-density families. Mol Psychiatry. 2004;9(10):962-7.
31. Kremer I, Pinto M, Murad I, Muhaheed M, Bannoura I, Muller DJ, et al. Family-based and case-control study of catechol-O-methyltransferase in schizophrenia among Palestinian Arabs. Am J Med Genet B Neuropsychiatr Genet. 2003;119B(1):35-9.
32. Shifman S, Bronstein M, Sternfeld M, Pisanté-Shalom A, Lev-Lehman E, Weizman A, et al. A highly significant association between a COMT haplotype and schizophrenia. Am J Hum Genet. 2002;71(6):1296-302.
33. Costas J, Sanjuán J, Ramos-Ríos R, Paz E, Agra S, Ivorra JL, et al. Heterozygosity at catechol-O-methyltransferase Val158Met and schizophrenia: new data and meta-analysis. J Psychiatr Res. 2011;45(1):7-14.
34. Hoenicka J, Garrido E, Martínez I, Ponce G, Aragüés M, Rodríguez-Jiménez R, et al.; PARGPARG. Gender-specific COMT Val158Met polymorphism association in Spanish schizophrenic patients. Am J Med Genet B Neuropsychiatr Genet. 2010;153B(1):79-85.
35. Komura D, Shen F, Ishikawa S, Fitch KR, Chen W, Zhang J, et al. Genome-wide detection of human copy number variations using high-density DNA oligonucleotide arrays. Genome Res. 2006;16(12):1575-84
Received in December, 2010.
Accepted for publication in June, 2011.
Hanlet Camacho, Centro de Ingeniería Genética y Biotecnología, CIGB Ave. 31 e/ 158 y 190, PO Box 6162, Cubanacán, Playa, La Habana, Cuba, E-mail: hamlet.camacho@cigb.edu.cu

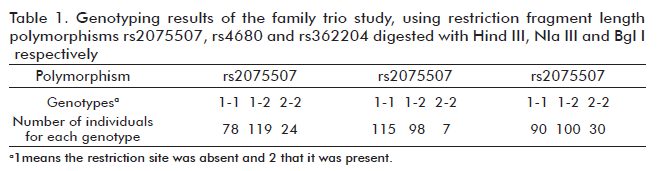{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}