










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBiotecnología Aplicada
versão On-line ISSN 1027-2852
Biotecnol Apl vol.29 no.1 La Habana jan.-mar. 2012
RESEARCH
Stabilization of glucose oxidase with cyclodextrin-branched carboxymethylcellulose
Estabilización de la glucosa oxidasa con carboximetilcelulosa ramificado con ciclodextrina
Madyu Matos1, Benjamin K Simpson2, Héctor L Ramírez1, Roberto Cao3, Juan J Torres-Labandeira4, Karel Hernández1
1Center for Enzyme Technology, Universidad Camilo Cienfuegos. Autopista a Varadero Km 3½, CP 44 740, Matanzas, Cuba.
2Food Science and Agricultural Chemistry Department, McGill University (Macdonald Campus). 21 111 Lakeshore Road, Ste Anne de Bellevue, Québec, Canada H9X 3V9.
3Laboratory of Bioinorganic Chemistry, Faculty of Chemistry, University of Havana, UH, PO Box 10 400, Havana, Cuba.
4Pharmaceutical Technology Department, Faculty of Pharmacy, Santiago de Compostela University Campus Vida, PO Box 15 782, Santiago de Compostela, Spain.
ABSTRACT
We present a methodology for improving some enzymatic characteristics of glucose oxidase (GOx) (EC 1.1.3.4). The enzyme was chemically modified with a β-cyclodextrin-branched carboxymethylcellulose polymer (CMC-CD), using carbodiimide as coupling agent. The obtained neoglycoenzyme had 0.78 mol of polysaccharide per mol of GOx and retained 67% of its initial activity. Comparison of some characteristics of the modified and free enzymes showed a higher Km for derivatized GOx and better thermostability, which increased from 45 °C to 51 °C. In addition, derivatization of GOx with CMC-CD increased its resistance to inactivation at 45 °C by 2.2-fold, protected the molecule against inactivation with the anionic surfactant sodium dodecylsulphate to the point that it retained 75% of its activity after an incubation of 3 h, and extended its pH tolerance toward alkaline pH (7.5). Covalent glycosidation of glucose oxidase with CD-branched carboxymethylcellulose polymer constitutes therefore an effective strategy for enhancing the stability of this enzyme.
Keywords: Carboxymethylcellulose, cyclodextrin, enzyme stability, glucose oxidase.
RESUMEN
La glucosa oxidasa (GOx; EC 1.1.3.4) se modificó químicamente con el polímero carboximetilcelulosa ramificado con unidades de β ciclodextrina (CMC-CD) utilizando una carbodiimida como agente acoplante. La neoglicoenzima contenía 0.78 mol de polisacáridos por mol de GOx y retuvo, después de la transformación, el 67% de su actividad inicial. Se compararon algunas características de la enzima modificada y la nativa. La enzima modificada mostró valores de Km superiores a la enzima libre. La termoestabilidad de la glucosa oxidasa modificada con el polímero CMC-CD se incrementó de 45 ºC hasta 51 ºC. A través de esta metodología, se logró una mayor estabilidad de la enzima modificada con CMC-CD, la cual mostró 2.2 veces más resistencia a la inactivación térmica a 45 ºC. Por otra parte, este derivado retuvo el 75% de su actividad inicial después de tres horas de incubación lo cual demuestra una remarcada estabilidad en presencia del surfactante aniónico duodecilsulfato de sodio y reveló una máxima actividad a valores de pH alcalinos (7.5). La glicosilación covalente de la glucosa oxidasa con el polímero CMC-CD puede ser utilizada efectivamente para el mejoramiento de la estabilidad de esta enzima.
Palabras clave: Carboximetilcelulosa, ciclodextrina, estabilidad de enzimas, glucosa oxidasa.
INTRODUCTION
Enzymes are useful tools for designing new and more efficient industrial processes. However, practical use of enzymes is often limited by their rapid inactivation by extreme temperature, pH or the presence of surfactants upon exposure to elevated temperatures [1].
Various approaches have been described for improving the stability of enzymes in aqueous media; chemical modification using water-soluble molecules constitutes one of the most promising [1]. In particular, the covalent derivatization of enzymes with carbohydrate moieties has received considerable attention in the field of enzyme technology [2]. This strategy has been based on the structural and functional stability that oligosaccharide chains confer to natural occurring glycoenzymes [3]. The structural and functional properties of carbohydrates, such as water solubility, biocompatibility and non-toxicity, have favored the use of these compounds for preparing artificially modifi ed enzymes, called ‘neoglycoenzymes’ [4].
Glucose oxidase (β-D-glucose:oxygen-oxidoreductase, EC 1.1.3.4, GOx) is a flavoprotein that catalyzes the oxidation of β-D-glucose to D-glucono-β-lactone and hydrogen peroxide, using molecular oxygen as electron acceptor [5, 6]. This redox enzyme is a dimeric glycoprotein of two identical subunits, having a molecular weight of 160 kDa [6]. Under denaturing conditions, the enzyme dissociates into its constituent subunits, releasing the catalytic flavin adenine dinucleotide (FAD) cofactor [7, 8].
Microbial GOx is currently receiving much attention due to its varied applications in chemical, pharmaceutical, food, beverage, clinical chemistry, biotechnological and other industries [9]. Novel applications of GOx in biosensors have increased demand for this molecule in recent years [10-12]. In general, highly robust GOx forms are desirable for these applications, which justify the development of strategies for increasing the stability of this redox enzyme. Several methods have been reported for stabilizing GOx, including the use of additives [13], immobilization in different supports [14], and engineering of recombinant GOXs through site-directed or random mutagenesis, coupled to expression in heterologous hosts [11].
Due to their exceptional properties as supramolecular host compounds, cyclodextrins have often been used to stabilize catalytically active enzyme conformations by covalent modification [15]. In fact, cyclodextrins are capable of forming stable inclusion complexes with hydrophobic guest compounds [16, 17], such as the amino acid side chains of surface-exposed residues in different enzymes, enhancing their stability and bioavailability [18-23].
The conjugation of carboxymethylcellulose (CMC) to β-Cyclodextrin (CD) moieties yields a water-soluble CMC-CD macromolecular structure. This polysaccharide can be easily linked to the ε-amino groups of protein lysine residues through a carbodiimide-catalyzed reaction [18]. This approach was employed for preparing neoglycoforms of trypsin [18] and α-amylase [19] in previous reports, and has also been shown to improve the pharmacological and pharmacokinetic characteristics of antioxidant enzymes [20, 21]. The remarkable stabilization conferred to these biocatalysts after glycosidation with CMC-CD has been attributed to the cooperative contribution of several factors, including covalent, ionic and supramolecular interactions between the protein backbone and polymer chains [19].
In this manuscript we report the covalent glycosidation of GOx with CD-branched CMC polymers, and the effects of this transformation on the catalytic properties and stability of this redox enzyme.
MATERIALS AND METHODS
Materials
GOx (E.C. 1.1.3.4) from Aspergillus niger, horseradish peroxidase (HRP, E.C. 1.11.1.7), and 2,2-azinobis(3-ethylbenzathiazoline-6-sulfonic acid) (ABTS) were from Sigma Chemical Co. (USA). Fractogel EMD BioSEC (S) and 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (EDAC) were purchased from Merck (Germany). Low-viscosity CMC, sodium salt (MW = 3 × 104, substitution degree = 0.7) was obtained from BDH (UK). CM-Sephadex C-25 was provided by Pharmacia Biotech (Sweden). CD was purchased from Amaizo (USA). All other chemicals were of analytical-grade quality.
Preparation of GOx-CMC-CD conjugate
CMC-CD was prepared by the reductive alkylation of peryodate-oxidized CMC with the mono-6-butylenediamine-6-deoxy-β-cyclodextrin derivative, as previously described [18, 19]. Coupling of the CD-branched polysaccharide to GOx was achieved by dissolving 10 mg of polymer in 5 mL of 20 mmol/L sodium phosphate buffer, pH 7 containing 10 mg of enzyme. The solution was stirred at 4 ºC during 1 h, and then 10 mg of 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide were added. The reaction mixture was stirred for 1 h at 25 °C and then for 16 h at 4 ºC. Afterwards, the solution was exhaustively dialyzed against 20 mmol/L sodium phosphate buffer, pH 7 and loaded on a Fractogel EMD BioSEC (S) (2.6 ×60 cm) gel filtration column equilibrated in the same buffer plus 100 mmol/L NaCl. The fractions containing the polymer-enzyme complex were pooled and stored at 4 °C.
Assays
GOx was determined spectrophotometrically by recording absorbance at 405 nm during the oxidation of ABTS by a coupled peroxidase-catalyzed reaction [24]. The reaction mixture consisted of 0.5 mL of 1 mol/L D-glucose in 100 mmol/L sodium phosphate buffer pH 6.0, 0.5 mL of ABTS (1 mg/mL) in distilled water, and 0.1 mL of HRP solution (2 mg/mL) in 100 mmol/L sodium phosphate buffer, pH 6.0. One unit of GOx was defined as the amount of enzyme that causes the oxidation of 1 μmol of ABTS per minute at 25 ºC under the described conditions.
Protein concentration was estimated by the Lowry method [25], using bovine serum albumin as standard. Total carbohydrate was determined by the phenol/sulfuric acid method, using D-glucose as standard [26]. The percentage of modifi ed amino groups was determined by using o-phtalaldehyde to determine free amino groups in native and CMC-CD modified protein, with glycine as standard [27].
Determination of kinetics parameters
Apparent Michaelis-Menten kinetic constants for free and modified GOx were determined by measuring enzyme reaction rates at substrate concentrations ranging from 0.004 to 20 μmol/L, at pH 6.0. The Michaelis constants were calculated by processing the data in Prism (GraphPad Software, USA), a software application designed for the analysis of enzyme kinetics.
Thermostability
Solutions of native and modifi ed GOx preparations (50 μg/mL of protein) were incubated at temperatures ranging from 25 ºC to 65 ºC in 100 mmol/L sodium phosphate buffer, pH 6.0. Aliquots were removed after 10 min of incubation, chilled quickly and assayed for enzyme activity. The values of T50 for each enzyme form, where T50 is defined as the temperature at which 50% of the initial activity remains, were determined from plots of residual activity (%) vs temperature.
Kinetics of thermal inactivation
Native and modified GOx preparations (50 μg/mL of protein) were incubated in 100 mmol/L sodium phosphate buffer pH 6.0 at temperatures ranging from 40 ºC to 55 ºC during 1 h. Aliquots were removed at different time points, chilled quickly and assayed for enzyme activity.
Stability in the presence of sodium dodecyl sulphate
Native and modified enzyme preparations (50 μg/mL of protein) were incubated at 30 ºC with 0.3% (w/v) sodium dodecyl sulphate (SDS) in 100 mmol/L sodium phosphate buffer, pH 6.0. Aliquots were removed at different time points, diluted 10-fold in 100 mmol/L sodium phosphate buffer, pH 6.0 and assayed for enzyme activity.
pH stability
Native and modifi ed enzyme forms (50 μg/mL protein) were incubated at 30 ºC in the following buffers: 50 mmol/L sodium acetate, pH 2.0-6.0; 50 mmol/L sodium phosphate, pH 7.0-7.5; and 50 mmol/L glycine-HCl, pH 8.0-11.0. Aliquots were removed after 30 min of incubation, diluted 10-fold in 100 mmol/L sodium phosphate buffer pH 6.0, and assayed for GOx activity.
Statistical analysis
All experiments were carried out in triplicate under identical conditions, using at least three replicate samples for each analytical determination. Mean values and standard deviations, and statistical charts, were calculated using Microsoft® Excel.
RESULTS AND DISCUSSION
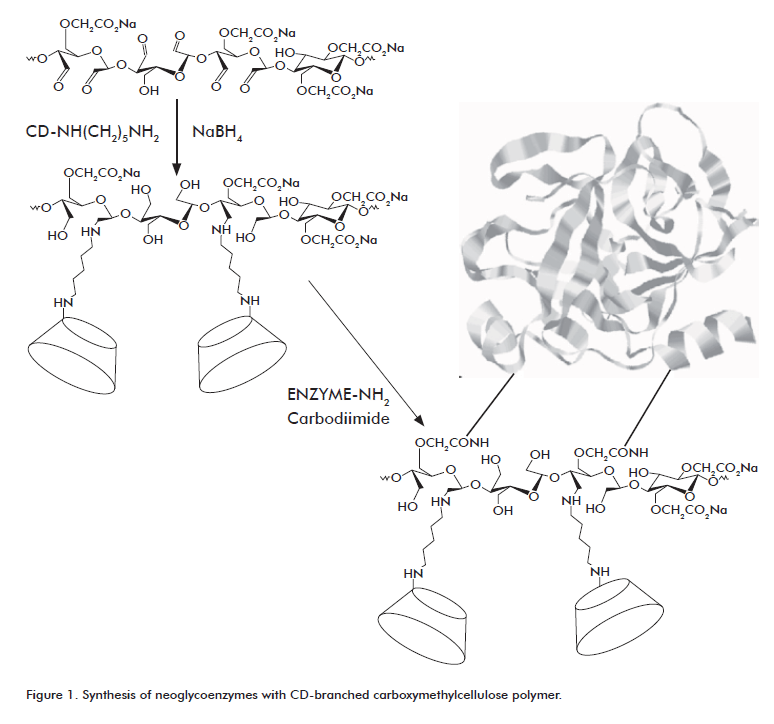
The polysaccharide used as modifying agent for GOx was prepared by oxidizing CMC with sodium metaperiodate, followed by conjugation to mono-6-hexylenediamino-6-deoxy-β cyclodextrin in the presence of NaBH4. The conjugation of CMC (molecular weight = 3.0 x 104, degree of carboxymethylation of 70%) with CD moieties yielded a water-soluble macromolecular structure with a 28:1 CD-polymer ratio, as reported by Ramírez [20]. This polysaccharide was covalently linked to free amino groups located at the surface of GOx using a water-soluble carbodiimide as coupling agent (Figure 1).
The carbohydrate content of the modified enzyme was estimated to be 244 mol (as D-glucose) per mol of protein. According to the chemical structure of CMC-CD [18, 19] and the sugar content of the native enzyme, this result represents an average of 0.78 mol of polysaccharide attached to each mol of GOx. The enzyme retained 67% of its specific activity after conjugation with CMC-CD. Loss of specific activity is probably associated to poorer accessibility of the active site due to steric hindrance from the attached polyanionic macromolecules. Chemical treatment per se may also play a role in this loss, as previously described [20].
The approach employed here was used previously to obtain a neoglycoform of Bacillus subtilis α-amylase that contained an average of 0.9 mol of polysaccharide per mol of protein and retained about 90% of the initial amylolytic activity [19]. Similar results were reported for trypsin [18].
The literature contains a number of different GOx stabilization strategies. Immobilization in glyoxylagarose, glutaraldehyde-agarose, MANAE-agarose and epoxide sephabeads resulted in activities ranging from 45 to 85% of initial values [14]. GOx has also been stabilized with LiCl as additive monovalent cation, producing relative enzyme activities of 78%. In this case, the GOx derivative was found to have a more compact dimeric structure, but lower enzyme activity than the native form [28].
An average of 21 mol of amino groups was found for the GOx-CMC-CD conjugate, as estimated with the o-phtaldehyde method [27]. In this complex, 27 mol of amino groups per mol of protein were modified by the same polysaccharide molecule, implying that the conjugate has a high degree of intramolecular crosslinking.
Native and modifi ed forms yielded Km values of 0.06 μmol/L and 1.09 μmol/L, respectively (Figure 2). Glycosidation decreased the affi nity of GOx for D-glucose by 18-fold. Values of kcat/Km for the native and modified enzymes were 11.33 μmol/L•s and 1.31 μmol/L•s respectively. As mentioned above, poorer catalytic performance of the modifi ed enzyme may be caused by steric hindrance from the attached polymer chains, resulting in lower accessibility of the active site for the substrate. In addition, the localized electrostatic gradient may affect Km by facilitating or obstructing interactions between charged groups in the enzyme or enzyme-substrate complex during binding and catalysis. A large number of small energetic gains and losses may complicate the analysis of such overall effects [29]. Similar effects on catalytic activity have been previously described for other enzyme-polysaccharide conjugates [23, 30] and for GOx entrapped in liposomes [31].
Importantly, hydrophobic interactions between CD and hydrophobic amino acid residues may reduce the dielectric constant of the enzymatic environment. Such a phenomenon would modify the dissociation constants of acid and basic groups in enzyme, substrate and buffer. Partition could cause changes in local concentrations of these molecules, affecting in turn the apparent kinetic parameters of the enzyme [29].
Studies on the reduced and oxidized holoenzyme as well as the apoenzyme revealed that in the native conformation of GOx, FAD is in close proximity to seven tryptophan residues. FAD plays an important role in the activity and structure of this enzyme [13]. Taking into account all of these factors, the existence of host-guest interactions between attached CD moieties and hydrophobic amino acid residues may affect the kinetics parameters of the GOx derivative.
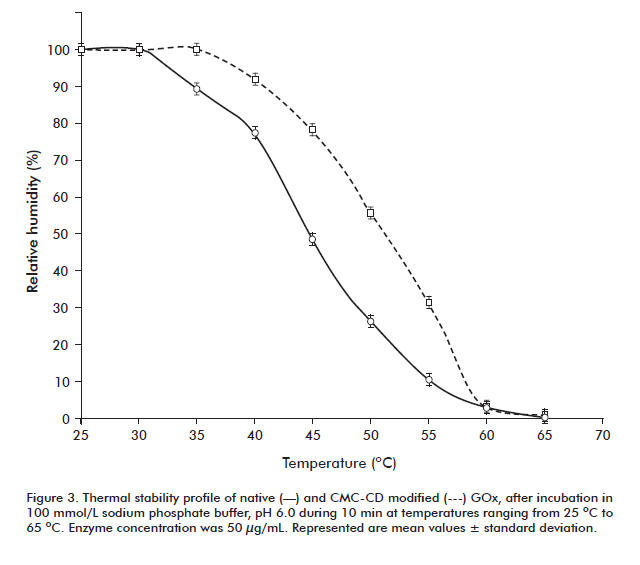
The thermal stability of the modified enzyme was investigated by heating the samples for 10 min at temperatures ranging from 25 ºC to 65 ºC and measuring their activity. Modified GOx was more resistant to temperatures above 30 ºC (Figure 3) than the native enzyme, as T50 increased from 45 ºC before glycosidation to 51 ºC afterwards. Similar results have been reported for GOx stabilized through the addition of KCl [28] and for different GOx immobilization variants [32-34].
The improved resistance of derivatized GOx against heat-induced denaturation is due, to a significant extent, to the multipoint nature of the enzyme-polymer linkage. Surface complementarity between enzyme and modifying agent has also been found to be crucial for the stabilization of GOx [32]. Previous studies have pointed at aggregation as the main cause of inactivation of GOx [13], which can be prevented by modifying the microenvironment of the enzyme. The multipoint interactions present in multimeric enzymes may prevent subunit dissociation by intersubunit cross-linking while simultaneously reducing conformational inactivation [33].
In a different set of experiments, the time course of the inactivation of free and modifi ed GOx was examined by incubation at temperatures ranging from 40 ºC to 55 ºC during 1 h. Compared to native GOx, the CMC-CD modified enzyme exhibited longer halflives (t1/2) at these temperatures (Figure 4). Maximum stabilization was achieved at 40 ºC, where t1/2 of CMC-CD-modified GOx was 5-fold higher than that of its native counterpart. Stability increases of 7.7- fold have been reported for GOx immobilized with lysozyme in the presence of additives and salts [34]. Considering the advantages associated with the use of enzymes at high temperatures, the higher thermal stability of GOx after glycosidation with CMC-CD is an important result.
Thermal denaturation of GOx proceeds mainly through the destabilization of ionic and hydrophobic interactions and the breakage of hydrogen bonds and van der Waals forces, leading to conformational changes in tertiary structure that inactivate the enzyme [5, 13]. The conformational stability observed for GOx modified with CD polymers results from the cooperative contribution of several factors, including the covalent cross-linking to macromolecular structures and the formation of new hydrogen bonds and electrostatic interactions at its surface. However, it can support the results due to the occurrence of host-guest interactions between the attached CD moieties and the amino acid residues at the surface of the modified proteins are also associated with the thermal stabilization [19, 23]. Similar results were obtained for CD-branched CMC derivatives of α-amylase [19], α-chymotrypsin [22], trypsin [18] and superoxide dismutase [20, 21].
GOx activity is markedly reduced in the presence of surfactants such as SDS, due to the rupture of non-polar interactions essential for maintaining the active conformation [7]. However, CMC-CD-modified GOx was remarkably more stable in the presence of SDS (Figure 5), retaining about 75% of its initial activity after incubation for 3 h. Improvement stabilization was found for CMC-CD modified GOx in the presence of SDS.
Similarly, the modified enzyme was more stable after incubation in solutions at different pH values (Figure 6). Optimum pH was 6.0 for both native and CMC-modified GOx, as reported earlier [28]. However, CMC-CD-modified GOx retained 70% of its activity at pH 7.5, compared with 20% for the native enzyme. The fact that the polymer-enzyme complex was more stable at this pH than its native counterpart suggests that the attached polymer chain protects the active enzyme conformation at alkaline pH and avoids protein unfolding. CMC-CD derivatization would thus represent an effective tool for improving enzyme stability at non-optimal pH.
Our results on the use of CD-branched CMC polymers as modifying agents for GOx suggest that they stabilize significantly the active conformation of the enzyme, preserving enzyme activity upon exposure to different inactivating/denaturing agents. This conformational stabilization may have its basis on the high degree of intramolecular cross-linking of the conjugate, caused by covalent multipoint attachment of the polymer chains to the enzyme protein structure, as mentioned above. Additional stabilization may be provided by multipoint anionic and supramolecular interactions between the enzyme and the CD-branched polymer CMC-CD polymer, as previously reported for similar neoglycoconjugates [18, 19]. This strategy may be advantageous for a number of GOx applications, considering the advantages of this enzyme and its use in different industries.
CONCLUSIONS
The present paper reports the covalent derivatization of GOx with CD-CMC. Our results demonstrate the feasibility of this strategy for improving the stability of GOx in a number of denaturing conditions. We suggest that covalent glycosidation of redox enzymes with CD-branched CMC polymers might be a useful alternative for conferring functional resistance to these biocatalysts.
ACKNOWLEDGMENTS
The author appreciates the valuable technical assistance of Dr Reynaldo Villalonga Santana.
REFERENCES
1. Iyer PV, Ananthanarayan L. Enzyme stability and stabilization—Aqueous and non-aqueous environment. Process Biochem. 2008;43(10):1019-32.
2. Villalonga ML, Reyes G, Villalonga R. Metal-induced stabilization of trypsin modified with alpha-oxoglutaric acid. Biotechnol Lett. 2004;26(3):209-12.
3. Wang C, Eufemi M, Turano C, Giartosio A. Influence of the carbohydrate moiety on the stability of glycoproteins. Biochemistry. 1996;35(23):7299-307.
4. Mislovicova D, Masarova J, Svitel J, Mendichi R, Soltes L, Gemeiner P, et al. Neoglycoconjugates of mannan with bovine serum albumin and their interaction with lectin concanavalin A. Bioconjug Chem. 2002;13(1):136-42.
5. Zoldak G, Zubrik A, Musatov A, Stupak M, Sedlak E. Irreversible thermal denaturation of glucose oxidase from Aspergillus niger is the transition to the denatured state with residual structure. J Biol Chem. 2004;279(46):47601-9.
6. Tsuge H, Natsuaki O, Ohashi K. Purification, properties, and molecular features of glucose oxidase from Aspergillus niger. J Biochem. 1975;78(4):835-43.
7. Jones MN, Manley P, Wilkinson A. The dissociation of glucose oxidase by sodium n-dodecyl sulphate. Biochem J. 1982;203(1):285-91.
8. Caves MS, Derham BK, Jezek J, Freedman RB. The mechanism of inactivation of glucose oxidase from Penicillium amagasakiense under ambient storage conditions. Enzyme Microb Technol. 2011;49(1):79-87.
9. Bankar SB, Bule MV, Singhal RS, Ananthanarayan L. Glucose oxidase--an overview. Biotechnol Adv. 2009;27(4):489-501.
10. Du Y, Luo XL, Xu JJ, Chen HY. A simple method to fabricate a chitosan-gold nanoparticles film and its application in glucose biosensor. Bioelectrochemistry. 2007;70(2):342-7.
11. Courjean O, Mano N. Recombinant glucose oxidase from Penicillium amagasakiense for efficient bioelectrochemical applications in physiological conditions. J Biotechnol. 2011;151(1):122-9.
12. Oztekin Y, Ramanaviciene A, Yazicigil Z, Solak AO, Ramanavicius A. Direct electron transfer from glucose oxidase immobilized on polyphenanthroline-modified glassy carbon electrode. Biosens Bioelectron. 2011;26(5):2541-6.
13. Gouda MD, Singh SA, Rao AG, Thakur MS, Karanth NG. Thermal inactivation of glucose oxidase. Mechanism and stabilization using additives. J Biol Chem. 2003;278(27):24324-33.
14. Betancor L, Lopez-Gallego F, Hidalgo A, Alonso-Morales N, Dellamora-Ortiz G, Guisan JM, et al. Preparation of a very stable immobilized biocatalyst of glucose oxidase from Aspergillus niger. J Biotechnol. 2006;121(2):284-9.
15. Wenz G. Cyclodextrins as building-blocks for supramolecular structures and functional units. Angew Chem Int Ed Engl. 1994;33(8):803-22.
16. Atwood JL, Davies JED, MacNicol DD, Vögtle F, Lehn J-M, editors. Comprehensive supramolecular chemistry. Oxford: Pergamon; 1996. p. 451-82.
17. Fulton DA, Fraser Stoddart J. Neoglycoconjugates based on cyclodextrins and calixarenes. Bioconjugate Chem. 2001;12(5):655-72.
18. Villalonga ML, Fernandez M, Fragoso A, Cao R, Villalonga R. Functional stabilization of trypsin by conjugation with beta-cyclodextrin-modified carboxymethylcellulose. Prep Biochem Biotechnol. 2003;33(1):53-66.
19. Darias R, Herrera I, Fragoso A, Cao R, Villalonga R. Supramolecular interactions mediated thermal stabilization for α-amylase modified with a β-cyclodextrin-carboxymethylcellulose polymer. Biotechnol Lett. 2002;24(20):1665-8.
20. Ramirez HL. Cyclodextrin-grafted polysaccharides as carrier systems for naproxen [dissertation]. Havana City: Havana University; 2008.
21. Valdivia A, Pérez Y, Cao R, Baños M, García A, Villalonga R. Bienzymatic supramolecular complex of catalase and superoxide dismutase: Stability and anti-Inflammatory properties. Macromol Biosci. 2007;7(1):70-5.
22. Fernández M, Villalonga ML, Fragoso A, Cao R, Baños M, Villalonga R. -Chymotrypsin stabilization by chemical conjugation with O-carboxymethyl-poly-β-cyclodextrin. Process Biochem. 2004;39(5):535-9.
23. Villalonga R, Tachibana S, Cao R, Matos M, Asano Y. Glycosidation of L-phenylalanine dehydrogenase with O-carboxymethyl-poly-β-cyclodextrin. Enzyme Microb Technol. 2007;40(3): 471-5.
24. Betancor L, Fuentes M, Dellamora-Ortiz G, López-Gallego F, Hidalgo A, Alonso-Morales N, et al. Dextran aldehyde coating of glucose oxidase immobilized on magnetic nanoparticles prevents its inactivation by gas bubbles. J Mol Catal B Enzym. 2005;32(3):97-101.
25. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem. 1951;193(1):265-75.
26. Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F. Colorimetric method for determination of sugars and related substances. Anal Chem. 1956;28(3):350-6.
27. Bruneel D, Schacht E. End group modification of pullulan. Polymer. 1995;36:169-72.
28. Ahmad A, Akhtar MS, Bhakuni V. Monovalent cation-induced conformational change in glucose oxidase leading to stabilization of the enzyme. Biochemistry. 2001;40(7):1945-55.
29. Chaplin MF. The preparation and kinetics of immobilized enzymes [Internet]. In: Chaplin M, Becke C, editors. Enzyme Tecnhology. Cambridge: Cambridge University Press; 1990 [cited 2011 Aug 27]. p. 80-89. Available from: http://www.lsbu.ac.uk/biology/enztech.
30. Gómez L, Ramírez HL, Villalonga R. Chemical modification of α-amylase by sodium alginate. Acta Biotechnol 2001;21(3):265-73.
31. Rodriguez-Nogales JM. Kinetic behaviour and stability of glucose oxidase entrapped in liposomes. J Chem Technol Biotechnol. 2004;79(1):72-8.
32. Rauf S, Ihsan A, Akhtar K, Ghauri MA, Rahman M, Anwar MA, et al. Glucose oxidase immobilization on a novel cellulose acetate-polymethylmethacrylate membrane. J Biotechnol. 2006;121(3):351-60.
33. Fernandez-Lafuente R, Rodríguez V, Mateo C, Penzol G, Hernandez-Justiz O, Irazoqui G, et al. Stabilization of multimeric enzymes via immobilization and post-immobilization techniques. J Mol Catal B Enzymatic. 1999;7:181-90.
34. Gulla KC, Gouda MD, Thakur MS, Karanth NG. Enhancement of stability of immobilized glucose oxidase by modification of free thiols generated by reducing disulfide bonds and using additives. Biosens Bioelectron. 2004;19(6):621-5.
Received in September, 2011.
Accepted for publication in January, 2012.
Madyu Matos. Center for Enzyme Technology, Universidad Camilo Cienfuegos. Autopista a Varadero Km 3½, CP 44 740, Matanzas, Cuba. E-mail: madyu.matos@umcc.cu.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}