










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.29 no.4 La Habana oct.-dic. 2012
REPORT
New antigen against avian influenza virus based on the fusion of the exposed domains of the H5 subtype hemagglutinin and the Gallus gallus CD154 protein
Desarrollo de un nuevo antígeno contra el virus de la influenza aviar basado en la fusión de dominios expuestos de la hemaglutinina viral del subtipo H5 y la proteína CD154 de Gallus gallus
Alaín González1, Julia Noda2, Armando Vega2, Alina Venereo1, Elsa Rodríguez1, María P Rodríguez1, Ernesto M González1, Damarys Díaz1, Raquel Montesino3, Elaine Santana1, Carlos Montero1, Alina Rodríguez1, Mario P Estrada1, Carlos Borroto1
1 Departamento de Biotecnología Animal, Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 11 600, La Habana, Cuba.
2 Departamento de Virología, Centro Nacional de Sanidad Agropecuaria, Censa. AP 10, CP 32700, San José de las Lajas, Mayabeque, Cuba.
3 Departamento de Química de Carbohidratos, CIGB, La Habana, Cuba.
ABSTRACT
The highly pathogenic H5N1 avian influenza virus (AIV) has caused important economic losses and public health problems, mainly in Asia, Africa and Europe. Counteracting this virus by vaccination of poultry is crucial. In our study, recombinant antigens based on the fusion of the extracellular segments of both molecules, hemagglutinin H5 of AIV and the chicken CD154, were tested to enhance the immune response in chickens. For this purpose, chickens were immunized with recombinant adenoviral vectors carrying the nucleotide sequence of the hemagglutinin H5 alone (AdHA) or fused to the CD154 recombinant protein (AdHACD). Also, these two proteins (rHA and rHACD) were produced and assayed as antigens. Both, the adenoviral vector AdHACD and the recombinant protein rHACD, induced hemagglutination-inhibiting antibody titers significantly higher than their counterparts carrying HA alone. Similar results were observed when the cellular response was measured by real time PCR. AdHACD and rHACD significantly increased the production of IFN-γ transcripts, which were lower when AdHA or rHA were used. To obtain a higher quality antigen, rHACD was purified by size exclusion chromatography, resulting in the preservation of its immunogenic properties. However, rHACD did not work as immunogen when an immunoaffinity chromatography was performed by eluting at acidic pH or with a chaotropic agent. These results demonstrate that chicken CD154 can significantly enhance humoral and cellular responses against hemagglutinin H5 of AIV when both molecules are administered as a fusion protein in chickens. Therefore, AdHACD or rHACD could become promising vaccine candidates against H5N1 virus.
Keywords: avian influenza virus, hemagglutinin, CD154, influenza vaccine, immune response, adenoviral vector.
RESUMEN
El virus de la influenza aviar (AIV) H5N1 ha causado serios problemas económicos y de salud pública en Asia, África y Europa. Por este motivo, la protección de la masa avícola contra este virus es fundamental. En este estudio se desarrollaron dos antígenos basados en la fusión del segmento expuesto de la hemaglutinina H5 del AIV con el segmento expuesto del CD154 de pollo. Estos se inmunizaron como vectores adenovirales recombinantes que contenían la hemaglutinina H5 simple (AdHA) o la proteína de fusión (AdHACD), o como antígenos proteicos recombinantes rHA y rHACD. Tanto el AdHACD como la rHACD indujeron títulos de anticuerpos inhibidores de la hemaglutinación significativamente superiores a los obtenidos con el AdHA y la rHA. Al medir la respuesta celular por PCR en tiempo real, los resultados fueron similares. El AdHACD y la rHACD incrementaron significativamente la producción de transcritos de IFN-γ. Estos fueron menores con el AdHA y la rHA. Para obtener un antígeno de mayor calidad, se purificó la rHACD por cromatografía de exclusión molecular. Este método mantuvo la inmunogenicidad de la rHACD. Sin embargo, cuando se realizó una cromatografía de inmunoafinidad mediante elución a pH ácido o con un agente caotrópico, la rHACD no funcionó como inmunógeno. Estos resultados demuestran que el CD154 de pollo puede incrementar significativamente la inmunidad humoral y celular contra la hemaglutinina H5 del AIV, cuando ambas moléculas se administran fusionadas en pollos. Por tanto, el AdHACD y la rHACD podrían convertirse en candidatos vacunales prometedores contra el virus H5N1.
Palabras clave: virus de influenza aviar, hemaglutinina, CD154, vacunas contra influenza, respuesta inmune, vector de adenovirus.
INTRODUCTION
Avian influenza is a highly contagious, systemic viral disease affecting chickens, turkeys and other poultry [1]. Its causative agent, the avian influenza virus (AIV), attacks several organs causing 100% of mortality rate. The rapid spread of AIV, beyond its natural barriers, may hinder international trade of birds and their products, also causing economic loses and serious public health problems.
During the twentieth century, there were three influenza pandemics with millions of deaths worldwide. In the last two pandemics, the infectious agent was a chimeric virus which carried genes from human and avian strain. Therefore, AIV is considered a key contributor for human influenza pandemics. In 2009, a pandemic strain originated in Mexico from subtype, H1N1 was identified as a reassortant of one avian, two swine and one human strains. That virus affected several countries in America and Europe. Due to the increased severity and magnitude of AIV outbreaks, the World Health Organization (WHO) has warned on the emergence of a new pandemic caused by a highly pathogenic AIV strain, with devastating effects [2].
Since 2003, the highly pathogenic AIV H5N1 has caused outbreaks in Southeast Asia and has spread across Africa and Europe, with losses of millions of birds and the subsequent negative economic impact. This viral subtype has also been able to infect humans who get into close contact with infected birds or contaminated water [3]. So far, the main transmission pathway is from animals to humans, and the continued presence of the infection in poultry may increase the risk of the emergence of a new viral subtype with potential to spread among humans. Therefore, the protection and control of poultry are critical.
WHO recommended vaccination combined with biosecurity measures to prevent possible outbreaks of the H5N1 virus. Although conventional vaccines currently used are considered effective in the protection against clinical symptoms and decreased the viral excretion to the environment, processing under emergency conditions may be limited by the use of the egg-dependent production systems [4].
Subunit vaccines could reverse this scenario, due to its simple, fast and secure manufacturing. They could also allow differentiation between infected and vaccinated animals. In this sense, hemagglutinin (HA) has been the IAV protein most widely used to develop candidate vaccines, as the major antigenic determinant, and was obtained in insect cells using baculovirus vectors as expression system [5]. However, the immunogenicity of subunit vaccines is lower than that of conventional vaccines. The use of molecular adjuvants is one of the strategies used to circumvent the limited immunogenicity of subunit vaccines.
One of the molecules used for this purpose is the glycoprotein CD154, the ligand of the CD40 co-stimulatory receptor, which is expressed as a type II integral membrane protein and belongs to the superfamily of tumor necrosis factors. The interaction of CD154 with CD40 is essential both for humoral and cellular immune responses [6]. It also provides signals for the activation and maturation of dendritic cells. Moreover, this interaction promotes B cell activation and their clonal expansion, the formation of germinal centers, isotype switching and the generation of memory B cells [7].
A number of studies have shown that murine CD154 may act as molecular adjuvant by enhancing the immune response [8, 9]. Recently, duck and chicken CD154 were cloned and sequenced. Furthermore, it was demonstrated that their functions are equivalent to the mammalian CD154 [10].
So far, studies of HA-based vaccine candidates have focused on obtaining better expression and purification systems of the molecule. A plausible strategy to increase its immunogenicity comprises generating fusion proteins of HA to molecular adjuvants (for example, C3d, used as DNA vaccine [11]). This study was aimed at developing a new antigen against AIV based on the fusion of the exposed domains of both, the viral hemagglutinin subtype H5 and the chicken CD154 molecules. For this purpose, two adenoviral vectors were generated expressing in cell culture the exposed domain of the viral HA subtype H5 alone or fused to the exposed domain of the chicken CD154. The two glycoproteins were purified after been expressed by recombinant adenoviral vectors (HA and HACD), and their immunogenicity evaluated in chickens by detecting hemagglutination-inhibiting antibodies (HIA), and the cellular immune response by measuring IFN-γ expression by transcript real-time PCR. This research was granted the 2011 Award of the Cuban National Academy of Sciences.
RESULTS AND DISCUSSION
Expression and characterization of HA and HACD molecules
The ability of the adenoviral vectors AdHA and AdHACD to produce HA and HACD proteins, respectively, was assessed by the transduction of the murine mammary epithelial cell line HC11. Transduction efficiency was evaluated by direct observation at the fluorescence microscope 24 hours after adenoviral transduction. All the cells were successfully transduced with both adenoviral vectors at a multiplicity of infection (MOI) of 25, and showed an intense fluorescence level under the assayed conditions (Figure 1A).
SDS-PAGE and Western blot analyses of transduced mammalian cells showed that HA and HACD proteins were secreted to the culture medium [13]. In the case of HA, multimers were barely detected. Under reducing conditions, the HA protein generated three bands, the upper one corresponding to the native precursor protein HA0 and the other two to the HA1 and HA2 subunits resulting from the partial cleavage of HA0 by endogenous cellular proteases. In the case of HACD, intense multimeric bands were observed, with only two immunoreactive bands present after SDS-PAGE. These results were in agreement both with the cleavage pattern of HA and CD154 multimerization, the latter being essential for the biological function of CD154 [14].
For a better characterization, HA and HACD proteins were N-deglycosylated by PNGase-F treatment [12]. The size of the N-deglycosylated precursor protein and its subunits were lower than that observed in the untreated proteins. There was a three-band pattern both for the HA and HACD molecules, consistent with the predicted molecular sizes. This result demonstrated that glycosylation was responsible for the increased sizes of the native proteins. It also altered the migration pattern of HACD, in which the faster of the two migrating bands actually contained the fusions of HA1 and HA2 subunits to CD154, which co-migrate together.
Immunogenicity evaluation of the adenoviral vectors AdHA and AdHACD in chickens
Both adenoviral constructs were assayed for immunogenicity in chickens. Primary immunization with the adenoviral vectors AdHA or AdHACD induced low titers of HIA [12]. However, the booster immunization on day 35 produced a rapid increase in HIA titers a week later, which began to decrease on days 49 and 56 for AdHA and AdHACD, respectively. HIA titers induced by AdHACD were significantly higher than those obtained with AdHA (p < 0.01 and p < 0.05, respectively).
The cellular response was determined by measuring the relative level of IFN-γ transcripts in peripheral blood mononuclear cells (PBMCs) of chickens immunized with 1 × 107 virus particles of either AdHA or AdHACD, by real-time PCR (Figure 1B) [12]. The expression of IFN-γ in PBMCs was induced in both groups. The levels of IFN-γ transcripts of chickens vaccinated with AdHACD were significantly higher than those obtained with AdHA (p < 0.01). This is consistent with a potentially better activation of CD4+ T cells and B cell cooperation in the group immunized with AdHACD than in animals only receiving HA, also resulting in an increased humoral response [8]. Moreover, CD154 has been demonstrated to induce CD8+ T cell responses in its soluble form [9], also contributing to viral clearance. These results demonstrate that CD154 can act as a molecular adjuvant by significantly increasing the humoral and cellular immune responses.
Establishing an HACD expression system and evaluation of its immunogenicity
Since the immunogenicity of the HACD protein was higher compared to that of HA molecule alone in the previous experiment, the HACD protein was selected for optimizing its expression system in cell lines of different species and at a range of viral concentrations. As determined by ELISA, the highest HACD concentration was obtained in the SiHa cell line, which was able to produce more than 4.5 mg/mL of the protein [13]. This value was significantly higher than those obtained in the other cell lines (p < 0.001), with HACD concentrations in the MDBK and HC11 cell lines about 2 mg/mL each. Subsequently, the SiHa cell line was subjected to several viral concentrations (MOI) in the range of 25 to 800. It was shown that the amount of HACD produced under these conditions was not proportional to MOI augmentation, increasing from MOI 25 to 200 and decaying at 400 and 800 [13]. During medium harvesting, dead cells started to appear at MOI 200. This phenomenon was more evident at MOIs 400 and 800. Therefore, MOI 100 was selected to produce the HACD protein, since it rendered the highest HACD concentration in the absence of short term cytopathic effects in host cells.
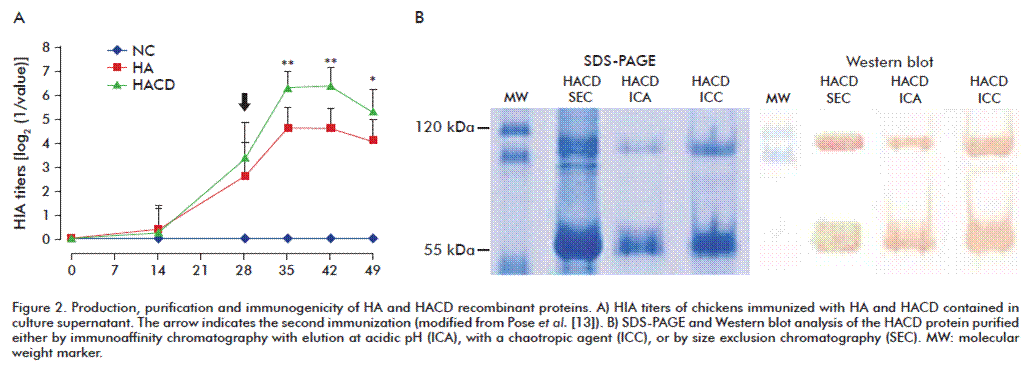
Then, the effect of HA and HACD proteins produced in cell culture on the immune system in chickens was evaluated, by administering the culture supernatant of either protein formulated in Montanide 888. The observed HIA kinetics was similar to that obtained after immunizing with the recombinant adenoviral vectors. Low HIA titers were induced for both proteins after the first immunization (Figure 2A), followed by increased titers after the booster dose for both proteins until day 42, which began to decline on day 49. Nevertheless, HIA antibody titers in animals immunized with HACD were significantly higher than those obtained with HA for all the determinations after the booster immunization (p < 0.01 vs. p < 0.05, respectively).
Purification of the HACD protein
In order to obtain a highly pure and cell culture impurity-free antigen, the HACD protein was subjected to three independent purification processes: immunoaffinity chromatography using elution 1) at acidic pH (ICA) or 2) with a chaotropic agent (ICC), and 3) size exclusion chromatography (SEC) (Figure 2B). SDS-PAGE protein fractionation and immunoidentification by Western blot confirmed the presence of the HACD protein after its purification by each of the three methods. The immunoaffinity chromatography, regardless the solutions used for the elution, yielded about 98% of purity. In the SEC method, HACD protein purity ranged 75-80%.
Immunogenicity evaluation of the HACD protein purified by different methods
The first immunization with the SEC-purified HACD antigen generated HIA titers in 40% of immunized chickens on day 28. The second immunization induced an immune response in 80% of immunized chickens on day 35. All chicks of this experimental group induced a humoral immune response on days 42 and 49. Although the HACD purified by ICA and ICC was about 98% pure, they did not induce antibody responses.
During the infection of the host cell by AIV, the HA protein undergoes conformational changes induced by the low pH at the endosomal compartment, which facilitates the fusion of the viral membrane and the escape of the virus to the cytoplasm [15]. Furthermore, chaotropic agents tend to distort the molecules by interfering with the non-covalent intramolecular interactions, further increasing entropy and unfolding the proteins on their primary structures. This total loss of native configuration caused by the acidic conditions or the chaotropic agent during immunoaffinity chromatography may hinder the HACD protein from gaining its native structure during renaturation. This issue could constitute an obstacle for the recovery of haemagglutinating epitopes. Otherwise, only the HACD protein obtained by SEC induced a humoral response in vaccinated chickens (4 out of 10 on day 28, 8/10 on day 35 and all the animals seroconverted on day 49). HACD separation by molecular weight does not damage its structure and helps to preserve the native configuration, which is responsible of inducing HIA. The results of cellular responses developed in the groups immunized with culture supernatants containing the antigens HA and HACD, the HACD purified by SEC or the negative control, showed a significantly higher level of stimulation in the groups immunized with HACD compared to the group immunized with HA or the control group (p < 0.01). However, no significant differences were observed between the groups immunized with culture supernatants or purified HACD. Although purity levels of HACD showed no differences in PBMC secretion of IFN-γ, both were significantly higher than the levels observed for the HA protein alone, which, at the same time, showed no significant differences compared to the negative control [13]. In summary, the presence of the chicken CD154 domain in the chimeric protein demonstrates its potential as molecular adjuvant in the induction of a superior cellular response against the antigen HA.
RELEVANCE OF THE STUDY
The experiments with adenoviral vectors and recombinant proteins shown herein demonstrated that the fusion of the exposed segment of the viral hemagglutinin subtype H5 to the exposed domain of the chicken CD154 allows an increased humoral and cellular immunity against the hemagglutinin of avian influenza virus. Moreover, the versatility of the recombinant adenoviral vectors enables the production of the vaccine antigen in tissue culture or the use of these adenoviral vectors as antigens for immunization of chickens in emergency conditions. This adenoviral platform also allows the replacement of the hemagglutinin nucleotide sequence in an easy and quick way. Hence, a new adenoviral construction with the HA of the circulating strain can always be obtained. In addition, the presence of the exposed segment of the chicken CD154 assures an increased cellular response with preserved T epitopes of HA, which might confer some protection against heterologous strains. Also, the ability of the fused antigen to increase the adaptive immune response could reduce the cost of a recombinant vaccine, based on the induction of a protective immune response with a lower antigen amount.
CONCLUSIONS
Our study demonstrates increased humoral and cellular immune responses of the HACD molecule, which could be used as a potential subunit vaccine candidate against the highly pathogenic AIV subtype H5N1. This could avoid several limitations of conventional vaccines, such as the limited production capacity due to the use of embryonated eggs and costs associated to high contention facilities. Although competent systems for protein expression and purification are needed for manufacturing an efficient subunit vaccine, and protection by challenging vaccinated chickens with an H5 highly pathogenic avian influenza strain has to be demonstrated, the strategy of enhancing antigen immunogenicity by its fusion to the molecular adjuvant CD154 could lower vaccine production costs and the administered dose per chicken. Moreover, as the efficacy of unpurified antigen was also demonstrated, it could be used for chicken vaccination in an emergency situation if its safety is demonstrated.
ACKNOWLEDGEMENTS
The authors thank the colleagues from the Center for Genetic Engineering and Biotechnology of Sancti-Spíritus for providing the immunochemicals for antigen detection. They are also grateful for their contribution to this study to Enrique Pérez and Consuelo Pérez from Censa, and Sonia González, Yanaysi Ceballos and Anays Álvarez from the CIGB.
REFERENCES
1. Buscaglia C. Influenza Aviar. InVet. 2004;6(1):71-84.
2. Suguitan AL, Jr, Cheng X, Wang W, Wang S, Jin H, Lu S. Influenza H5 hemagglutinin DNA primes the antibody response elicited by the live attenuated influenza A/Vietnam/1203/2004 vaccine in ferrets. PLoS One. 2011;6(7):e21942.
3. Horimoto T, Kawaoka Y. Pandemic threat posed by avian influenza A viruses. Clin Microbiol Rev. 2001;14(1):129-49.
4. Hoffmann E, Krauss S, Perez D, Webby R, Webster RG. Eight-plasmid system for rapid generation of influenza virus vaccines. Vaccine. 2002;20(25-26):3165-70.
5. McPherson CE. Development of a novel recombinant influenza vaccine in insect cells. Biologicals. 2008;36(6):350-3.
6. Pentcheva-Hoang T, Corse E, Allison JP. Negative regulators of T-cell activation: potential targets for therapeutic intervention in cancer, autoimmune disease, and persistent infections. Immunol Rev. 2009;229(1):67-87.
7. Gray D, Siepmann K, Wohlleben G. CD40 ligation in B cell activation, isotype switching and memory development. Semin Immunol. 1994;6(5):303-10.
8. Li W. Synergistic antibody induction by antigen-CD40 ligand fusion protein as improved immunogen. Immunology. 2005;115(2):215-22.
9. Stone GW, Barzee S, Snarsky V, Kee K, Spina CA, Yu XF, et al. Multimeric soluble CD40 ligand and GITR ligand as adjuvants for human immunodeficiency virus DNA vaccines. J Virol. 2006;80(4):1762-72.
10. Fischer KP, Gares SL, Wang D, Lorne Tyrrell D, Gutfreund KS. Identification and characterization of functional CD154 (CD40 ligand) in the Pekin duck. Dev Comp Immunol. 2007;31(1):61-71.
11. Ross TM, Xu Y, Bright RA, Robinson HL. C3d enhancement of antibodies to hemagglutinin accelerates protection against influenza virus challenge. Nat Immunol. 2000;1(2):127-31.
12. Sanchez Ramos O, Gonzalez Pose A, Gomez-Puerta S, Noda Gomez J, Vega Redondo A, Aguila Benites JC, et al. Avian CD154 enhances humoral and cellular immune responses induced by an adenovirus vector-based vaccine in chickens. Comp Immunol Microbiol Infect Dis. 2011;34(3):259-65.
13. Pose AG, Gomez JN, Sanchez AV, Redondo AV, Rodriguez ER, Segui RM, et al. Subunit influenza vaccine candidate based on CD154 fused to HAH5 increases the antibody titers and cellular immune response in chickens. Vet Microbiol. 2011;152(3-4):328-37.
14. Peitsch MC, Jongeneel CV. A 3-D model for the CD40 ligand predicts that it is a compact trimer similar to the tumor necrosis factors. Int Immunol. 1993;5(2):233-8.
15. Gaudin Y, Ruigrok RW, Brunner J. Low-pH induced conformational changes in viral fusion proteins: implications for the fusion mechanism. J Gen Virol. 1995;76(Pt 7):1541-56.
Alaín González. Departamento de Biotecnología Animal, Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 11 600, La Habana, Cuba. E-mail: alain.pose@cigb.edu.cu.

{kind=link}
{kind=link}