










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.30 no.1 La Habana ene.-mar. 2013
RESEARCH
Molecular typing of Streptococcus suis from pigs in Cuba
Tipificación molecular de Streptococcus suis aislado de cerdos en Cuba
Ivette Espinosa, Michel Báez, Belkis Corona, Daine Chong, Evelyn Lobo, Siomara Martínez
División de Biología Molecular, Centro Nacional de Sanidad Agropecuaria, Censa. AP 10, San José de las Lajas, Mayabeque, Cuba.
ABSTRACT
Streptococcus suis is a bacterium commonly carried by pigs in the respiratory tract; thus the infections caused by virulent strains are considered a problem in the swine industry. A successful approach for the identification of virulent strains is the differentiation of capsular serotypes using specific antisera or the corresponding cps types by genotypic assessment, with the subsequent detection of virulence associated factors, namely the extracellular factor, the muramidase-released protein and the hemolysin suilysin. Data regarding serological and molecular identification of S. suis from pigs in Cuba are not available. This study was aimed at identifying the capsular types cps2, 7 and 9, as well as three genes related to virulence using PCR assays. According to the results, 31 isolates were evaluated and classified as cps2 (n = 21) or cps9 (n = 4), while six isolates not were typable. Considering the presence in these isolates of the genes mrp, epf and sly, six different genotypes were differentiated among the cps2 or cps9 strains and there were three non-typable isolates for the genes used in this study. The cps2 isolates were recovered from pigs between 6-12 and 14-17 weeks with pneumonia and systemic infection respectively, whereas the cps9 isolates were exclusively associated with pneumonia.
Keywords: Streptococcus suis, virulence markers, genotypes.
RESUMEN
Streptococcus suis es una bacteria común en el tracto respiratorio de los cerdos; sin embargo las infecciones causadas por cepas virulentas se consideran un problema en la industria porcina. Estas cepas se pueden identificar por la diferenciación de los serotipos capsulares con antisueros específicos o por sus correspondientes tipos cps, mediante ensayos genotípicos. También se pueden reconocer mediante la detección de factores asociados con la virulencia, como el factor extracelular, la proteína liberada por muramidasa y la hemolisina suilisina. Antes de esta investigación no había datos sobre la identificación serológica o molecular de los tipos capsulares de S. suis procedentes de cerdos en Cuba. El objetivo de esta investigación fue la detección de los tipos capsulares cps2, cps7 y cps9, y de los tres genes asociados con la virulencia, mediante ensayos de reacción en cadena de la polimerasa. De 31 aislamientos, 21 se clasificaron con el genotipo capsular cps2, y 4 con el cps9. El genotipo capsular cps7 no se detectó y 6 aislados no se correspondieron con ninguno de los serotipos analizados. Considerando la presencia de los genes sly, epf y mrp, se identificaron seis genotipos en los aislados cps2 y cps9, y 3 en los aislados cuyo genotipo cps no se identificó. Los aislados cps2 se extrajeron de cerdos de 6 a 12 con neumonía y de 14 a 17 semanas con infección sistémica; mientras que los cps9 estaban asociados exclusivamente con neumonía.
Palabras clave: Streptococcus suis, marcadores de virulencia, genotipos.
INTRODUCTION
Streptococcus suis has emerged as an important swine pathogen in most countries with an intensive swine production [1]. S. suis infections can be the cause of meningitis, polyarthritis, polyserositis, endocarditis, septicaemia, pneumonia and sudden death, leading to significant economic losses in pig farms [2-6]. This bacterium is also a zoonotic agent responsible for meningitis and arthritis in humans in close contact with infected pigs [6-8].
S. suis is a heterogeneous species that can be divided into 33 serotypes with respect to the capsule polysaccharides. Worldwide, capsular serotype 2 (cps2) is the most prevalent one among invasive porcine and human isolates [9, 10]. However, the importance of particular serotypes can vary geographically. Serotypes cps9 and cps7 are particularly reported common in Europe [4]. Major parts of the cps loci that are responsible for the biosynthesis of the capsule polysaccharides of the greatest concern serotypes 1 (and 14), 2 (and 1/2), 7 and 9, have been sequenced [11-14]. PCR tests based on sequences of type-specific capsular genes have been developed, for other serotypes diagnostic methods (although unreliable) are available [11-14].
Virulence can vary substantially both within and among serotypes, and not all isolates of the same serotype cause the same disease [4]. Although little is known about potential virulence factors, such as the hemolysin suilysin (SLY; encoded by the sly gene), the muramidase-released protein (MRP; mrp), and the extracellular factor (EF, epf) are virulence markers that have been used in elaborated genotypic and phenotypic schemes to try to predict the virulence of a given S. suis strain [15-17]. It is now included in routine diagnosis of S. suis in several laboratories [4].
In Cuba, the pig production has increased since 1997, and an intensive technology is being practiced in a considerable number of farms (30 % of the swine population) [18]. S. suis isolatesHowever, data regarding typing from pigs in Cuba are not available. In this work, the molecular identification of the capsule polysaccharide genes (cps) and the genotypic assessment of virulence markers in Cuban S. suis isolates were carried out for the first time.
MATERIALS AND METHODS
Samples and classification
A total of 31 isolates of S. suis from the collection of the Bacteriology Department of the National Center of Animal and Plant Health (Censa, Cuba) were included in this study. Most of these isolates were collected from diseased or dead animals during diagnostic procedures from farrow-to-finish farms in the Western region of Cuba from 2004 to 2011.
According to their source of isolation and clinical symptoms, the isolates were distributed in three groups as follows: i) Invasive group (n = 3), three isolates from abscess of pigs 10-17 weeks of age with arthritis and one isolate from aborted foetus; ii) pneumonia group (n = 21), isolates from the lung of animals between 10-12 weeks of age; and iii) carrier group (n = 6), comprising two isolates of tonsillar biopsies from healthy animals, one isolate from the vaginal secretion and three isolates of semen ejaculates from mature boars of a pig breeding farm.
PCR
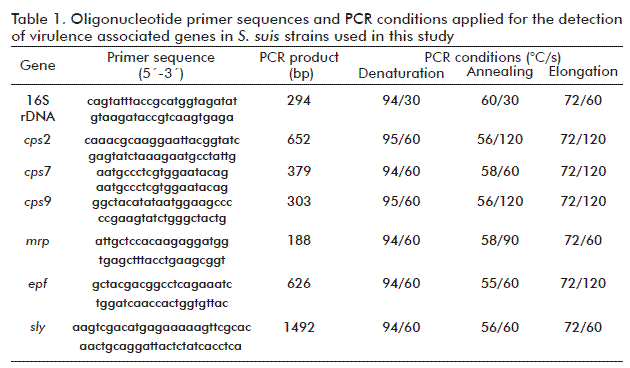
The isolates were identified by conventional biochemical procedures following the methods reported by Berthelot-Herault et al. [19]. The isolates were cultured in Todd Hewitt medium (5 mL) at 37 °C for 24 h and the DNA was extracted as described previously [20]. The S. suis-like strains were identified by amplification of a 294-bp-long 16S rDNA gene fragment by PCR using S. suis species-specific primers [13]. The capsular genotype (cps2) was determined by PCR as previously described by Smith et al. [12]. Capsular genotypes cps7 and cps9, and the mrp (188 bp) gene were analyzed by monoplex PCR assays using the sequence of primers and conditions described previously by Silva et al. [14]. Other two conventional PCR assays were used to screen the presence of sly (1492 bp) [21] and epf (626 bp) genes [17]. Details of all oligonucleotide primers used and PCR conditions applied for the detection are listed in table 1. Each isolate was tested under the same conditions twice.
The reaction was carried out in a 25-µL PCR mixture containing 20 pmol of each primer, 200 μM of dNTPs, 1× PCR buffer, 3 mM MgCl2 and 1 U Taq DNA polymerase (Invitrogen). The amplified DNA was visualized in 2 % agarose gels in TBE buffer (90 mM Tris, 90 mM borate, 2.5 mM EDTA; pH 8) for 1 h at a constant voltage of 125 V. Amplified products were stained with ethidium bromide (0.5 μg/μL) and detected by UV transillumination. The 50 bp ladder (Promega) was used as standard.
DNA sequencing
Partial sequences corresponding to cps2 and 16S rDNA genes were amplified by using chromosomal DNA obtained from sample 13 (designated as isolate SS13) collected in October 2010 from the lung of a pig with pneumonia. The sequences of both genes were determined by an automated sequence analyzer (CEQTM8800, Beckman Coulter, USA) using the same primers as for PCR amplification and Genome LabTM DTCS- Quick start kit. Prior to the sequencing reaction, PCR products were purified with the Wizard PCR purification kit (Promega) according to the manufacturer’s instructions.
Sequence analysis
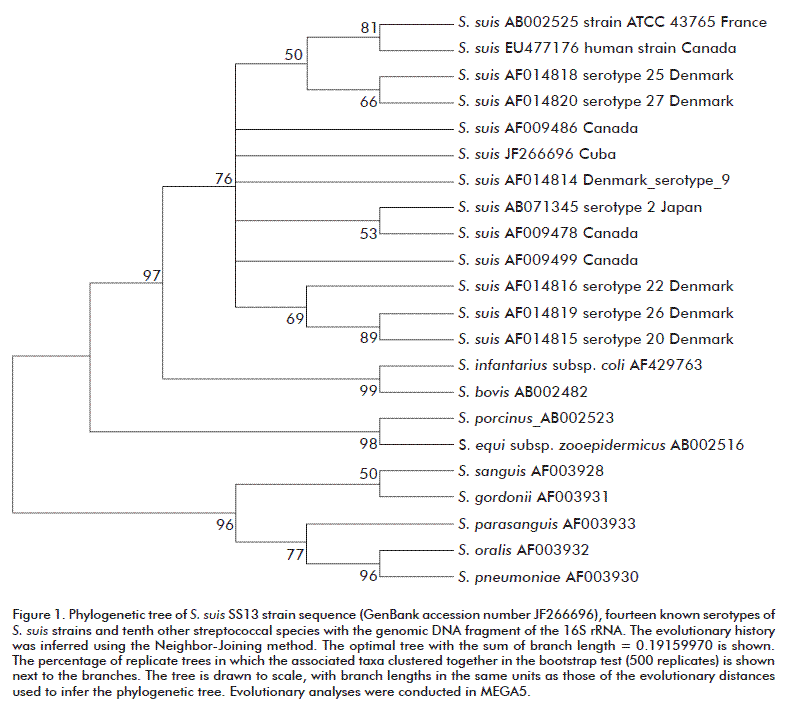
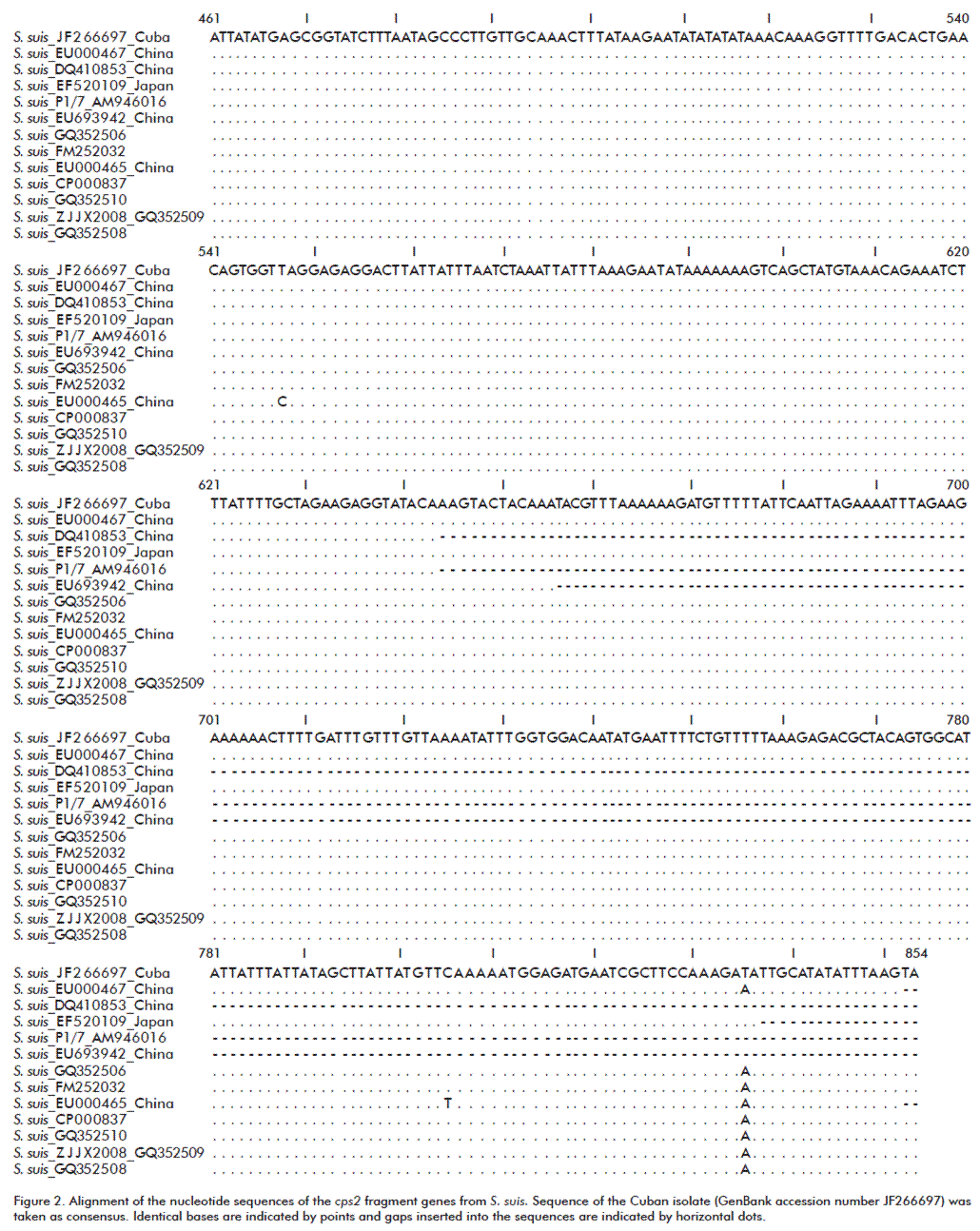
Sequences of both fragments corresponding to cps2 and 16S rDNA genes were aligned and assembled using the Vector NTI® software (Invitrogen, USA) to obtain two sequences of 260 and 637 bp, which were deposited in GenBank under accession numbers JF266696 and JF266697, respectively. They were aligned and studied using the MEGA5 software [22] for phylogenetic inference by using the Neighbor-Joining method [23]. The evolutionary distances were computed using the Kimura 2-parameter method [24]. The analysis involved 22 nucleotide sequences. To obtain a confidence value for the aligned sequence dataset, a bootstrap analysis [25] of 500 replications was done.
RESULTS AND DISCUSSION
All isolates included in this study showed a 294 bp fragment corresponding to 16S rDNA under the conditions described by Marois et al. [13]. The four isolates originating from animals with invasive procedures and the 21 isolates from pig lungs with pneumonia were isolated in pure culture. S. suis is an invasive pathogen and a very successful colonizer of mucosal surfaces, in particular of the upper respiratory tract. Reports about the isolation of this bacterium from pigs suffering arthritis and pneumonia are frequent [4]; thus, a potential causative role of thisorganism in respiratory diseases is also suggested [26]. For instance, most clinical S. suis isolates in Canada were from piglet lungs [27].
In addition, S. suis was identified in samples of tonsil, semen and vaginal discharge in healthy animals (Table 2). Dee and Corey [28] reported that S. suis strains can maintain their viability in different porcine fluids, such as urine or semen, up to 10 days. When S. suis is isolated from the tonsils and the genital and alimentary tracts of carrier pigs, those animals are discarded for being a source of S. suis transmission in herds [29, 30].
The 31 isolates of S. suis tested in this study demonstrated a high genetic diversity, being grouped into nine different genotypes with the aid of the previously described PCR assay (Table 2). Only genotypes cps2 (68 %) and cps9 (13 %) were detected, and six isolates from tonsils and semen of asymptomatic animals were non-typable for the three capsular genotypes investigated. The cps9 genotype was identified exclusively in isolates from lungs, while genotype cps2 was found in different isolates regardless of their origin (i.e., lungs, abscess in joints, aborted foetus, vaginal discharges or semen).
Epidemiological data have revealed a high diversity of serotypes among S. suis isolates, with serotype cps2 as the most associated with invasive diseases in humans [4, 14, 16, 31]; and serotypes 2, 9 and 14 are considered to be more invasive than others in pigs [14]. For S. suis serotype 2, it has been demonstrated that the capsule is an important virulence factor; however, unencapsulated S. suis strains might also invade host tissues, though to a lower degree [12]. The relevance of particular serotypes can vary geographically, even temporally. In China, cps2 accounts for more than 70 % of the systemic S. suis diseases in piglets [32], while in the Netherlands and Germany, cps9 emerged as the most common serotype responsible for a major fraction of invasive S. suis diseases of piglets [19]. On the other hand, cps7 strains have frequently been associated with bronchopneumonia in Scandinavia and Germany [11].
The identification of the S. suis SS13 cps2 isolate was confirmed by sequencing a fragment of the 16S rDNA gene and a fragment of cps2 gene. BLAST analysis of the sequences revealed a 99 % identity to S. suis type strain, which is consistent with the values expected for strains of a single species (99 % for 16S rDNA). The similarity level for the query sequence never exceeded 97 % with other Streptococcus species; these values consistent with distinct species in the same genus [25]. The phylogenetic relationship among the strains was shown with a tree (Figure 1), indicating a tight clustering of the SS13 isolate (JF266696) with known serotypes of S. suis. Other non-S. suis streptococcal strains were observed clearly separated from the S. suis cluster.
Multiple alignment of the nucleotide sequences of the cps2 fragment (JF266697) revealed that the sequence corresponded to strains of S. suis (Figure 2), only differing in one nucleotide change at position 836 (A→T), this mutation resulting in a possible amino acid variation from Lys279279 by Ile at the C-terminus.
The 1492 bp PCR product corresponding to the suilysin gene was identified in three cps2 isolates from animals with pneumonia and only in one non-typable isolate from tonsils of an asymptomatic animal (Table 2). Suilysin is a pore-forming cholesterol-dependent cytotoxin. Though suilysin is not essential for virulence of S. suis cps2 isolates in pigs, in vitro experiments suggest that it may have important functions for the interactions with host cells [5]. Tarradas et al. [33] detected suilysin production by isolated from diseased and healthy carrier pigs in Spain.
The 626 bp fragment of the gene encoding EF was detected only in two cps2 isolates. The EF was identified as an extracellular protein associated with virulence in serotypes 1, 2, 1/2, 14 and 15 [17]. The function of EF is still unknown, since isogenic epf mutants were as virulent as the wild-type in experimental infections, indicating that EF is associated but not essential for virulence [34].
The gene fragment encoding the MRP protein was amplified in cps2 isolates from cases of pneumonia and asymptomatic animals; this fragment was de-tected in the four cps9 isolates The function of the 136 kDa MRP protein is unknown. It was discovered as a factor released from virulent cps2 strains after muramidase treatment but expression of the MRP was found among strains of serotypes 1, 2, 1/2, 14 and 15 [17]. Different studies have shown that the presence of the gene did not always correlate with actual expression of the respective protein. Fittipaldi et al. [35] detected the mrp gene in 92 isolates of S. suis but the expression of MRP by Western blotting was only positive in 46 isolates, due, in most cases, to frameshift mutations in the 5´ end of the gene, re-sulting in premature stop codons. The effective production of the virulence markers by the tested isolates specifically in the genotype cps2 mrpepfslywould be also verified.
As mrp seems to be a silent gene in some strains, it might also be discussed that phenotyping rather than genotyping should be performed in diagnostic laboratories [4]. Although not ideal, typing methods based on protein expression of these markers confirm the phenotype because the PCR assays only detect a partial sequence of a gene. However, protein expression levels may be affected by many factors, as growth conditions, or even induced at specific stages of the infection process as it is known for many important, highly regulated virulence factors [36, 37]. The most promising candidate for improvement of S. suis diagnostics is the use of probes as the ivs-25/iri-1, which encodes a putative general virulence regulator. It discriminates between virulent and avirulent strains [37].
The analysis of the three virulence genetic markers showed the cps2+ mrp- epf- sly- as the most frequent genotype, in three isolates from abscesses of pigs with arthritis, one isolate from an aborted foetus and eight isolates from lungs. Only an isolate from lung showed the genotype cps2+ mrp+ epf + sly+. Despite the lack of evidence for a critical role of one or more of these putative virulence factors, they may nonetheless serve as virulence markers, since MRP, EF, and SLY are typical of Eurasian strains while they are almost absent among less virulent North American strains. [1, 26, 35]. Strains with the mrp+ epf+ sly+ genotype are known to be highly virulent, from European epidemiological studies [17] and pig experimental infections [4]. However, the percentage of S. suis cps2 strains recovered from diseased pigs and the number of cases of human disease is lower in North America than in other parts of the world [36]. Therefore, it is very probable that the virulence of the Cuban S. suis cps2 isolates associated with invasive processes is similar to that reported for North American strains. Further studies with a larger number of Cuban isolates are required to definitively confirm this hypothesis.
Serotype determination remains a valuable tool used by veterinary practitioners and diagnosticians to understand the epidemiology of a particular outbreak and/or to increase the possibility for success of a vaccination program for a given herd [35]. The PCR assays used in this study can differentiate the three cps genotypes (2, 7 and 9) and additionally detect the important virulence associated factors genes mrp, epf and sly (Table 2). Taking into account methods for serotyping, the PCR assays are rapid, reliable, and sensitive. Moreover, these PCR assays could be used directly with tonsillar specimens from infected or carrier animals so that isolation of single colonies could be omitted. Therefore, these tests will undoubtedly contribute to a more rapid and reliable diagnosis of S. suis and may facilitate control and eradication programs [12].
CONCLUSIONS
So far, this is the first study on the molecular identification of S. suis isolates in Cuba from disease cases in pigs. S. suis isolates belonging to the capsular serotypes cps2 and cps9 were detected by genotyping of the cps2 and cps9 genes, respectively. This is also the first report indicating the occurrence of at least two capsular types and nine different genotypes among the S. suis isolates tested. A larger scale sampling among pig farms from different Cuban regions could enable us to design a strategy to minimize the risk of exposure to S. suis.
ACKNOWLEDGMENTS
We thank Dr. Marcelo Gottschalk from Faculty of Veterinary Medicine, University of Montreal, Canada and Dr. Eduardo Sistach from National Center of Animal and Plant Health (Censa, Cuba) for their critical review of the manuscript.
REFERENCES
1. Berthelot-Herault F, Gottschalk M, Morvan H, Kobisch M. Dilemma of virulence of Streptococcus suis: Canadian isolate 89-1591 characterized as a virulent strain using a standardized experimental model in pigs. Can J Vet Res. 2005;69(3):236-40.
2. Kim D, Han K, Oh Y, Kim CH, Kang I, Lee J, et al. Distribution of capsular serotypes and virulence markers of Streptococcus suis isolated from pigs with polyserositis in Korea. Can J Vet Res. 2010;74(4):314-6.
3. Baums CG, Verkuhlen GJ, Rehm T, Silva LM, Beyerbach M, Pohlmeyer K, et al. Prevalence of Streptococcus suis genotypes in wild boars of Northwestern Germany. Appl Environ Microbiol. 2007;73(3):711-7.
4. Baums CG, Valentin-Weigand P. Surface-associated and secreted factors of Streptococcus suis in epidemiology, pathogenesis and vaccine development. Anim Health Res Rev. 2009;10(1):65-83.
5. King SJ, Heath PJ, Luque I, Tarradas C, Dowson CG, Whatmore AM. Distribution and genetic diversity of suilysin in Streptococcus suis isolated from different diseases of pigs and characterization of the genetic basis of suilysin absence. Infect Immun. 2001;69(12):7572-82.
6. Gottschalk M, Segura M. The pathogenesis of the meningitis caused by Streptococcus suis: the unresolved questions. Vet Microbiol. 2000;76(3):259-72.
7. Tang J, Wang C, Feng Y, Yang W, Song H, Chen Z, et al. Streptococcal toxic shock syndrome caused by Streptococcus suis serotype 2. PLoS Med. 2006;3(5):e151.
8. Wertheim HF, Nghia HD, Taylor W, Schultsz C. Streptococcus suis: an emerging human pathogen. Clin Infect Dis. 2009;48(5):617-25.
9. Heath PJ, Hunt BW. Streptococcus suis serotypes 3 to 28 associated with disease in pigs. Vet Rec. 2001;148(7):207-8.
10. Hill JE, Gottschalk M, Brousseau R, Harel J, Hemmingsen SM, Goh SH. Biochemical analysis, cpn60 and 16S rDNA sequence data indicate that Streptococcus suis serotypes 32 and 34, isolated from pigs, are Streptococcus orisratti. Vet Microbiol. 2005;107(1-2):63-9.
11. Smith HE, van Bruijnsvoort L, Buijs H, Wisselink HJ, Smits MA. Rapid PCR test for Streptococcus suis serotype 7. FEMS Microbiol Lett. 1999;178(2):265-70.
12. Smith HE, Veenbergen V, van der Velde J, Damman M, Wisselink HJ, Smits MA. The cps genes of Streptococcus suis serotypes 1, 2, and 9: development of rapid serotype-specific PCR assays. J Clin Microbiol. 1999;37(10):3146-52.
13. Marois C, Le Devendec L, Gottschalk M, Kobisch M. Detection and molecular typing of Streptococcus suis in tonsils from live pigs in France. Can J Vet Res. 2007;71(1):14-22.
14. Silva LM, Baums CG, Rehm T, Wisselink HJ, Goethe R, Valentin-Weigand P. Virulence-associated gene profiling of Streptococcus suis isolates by PCR. Vet Microbiol. 2006;115(1-3):117-27.
15. Beineke A, Bennecke K, Neis C, Schroder C, Waldmann KH, Baumgartner W, et al. Comparative evaluation of virulence and pathology of Streptococcus suis serotypes 2 and 9 in experimentally infected growers. Vet Microbiol. 2008;128(3-4):423-30.
16. Del’Arco AE, Santos JL, Bevilacquia PD, Faria JE, Guimaraes WV. Swine infection by Streptococcus suis: a retrospective study. Arq Bras Med Vet Zootec. 2008;60(4):878-83.
17. Wisselink HJ, Smith HE, Stockhofe-Zurwieden N, Peperkamp K, Vecht U. Distribution of capsular types and production of muramidase-released protein (MRP) and extracellular factor (EF) of Streptococcus suis strains isolated from diseased pigs in seven European countries. Vet Microbiol. 2000;74(3):237-48.
18. Muñiz M, Valdivia MP, Heredia J, López O, Arias T, Mederos CM, et al. Gestión tecnológica en la producción porcina cubana. In: VI Encuentro regional sobre nutrición y producción de especies monogástricas. La Habana: Asociación Cubana de produc-ción animal; 2001. Available from: http://www.sian.info.ve/porcinos/publicaciones/viencuent/marisol.htm.
19. Berthelot-Herault F, Morvan H, Keribin AM, Gottschalk M, Kobisch M. Production of muraminidase-released protein (MRP), extracellular factor (EF) and suilysin by field isolates of Streptococcus suis capsular types 2, 1/2, 9, 7 and 3 isolated from swine in France. Vet Res. 2000;31(5):473-9.
20. Luther DA, Almeida RA, Oliver SP. Elucidation of the DNA sequence of Streptococcus uberis adhesion molecule gene (sua) and detection of sua in strains of Streptococcus uberis isolated from geographically diverse locations. Vet Microbiol. 2008;128(3-4):304-12.
21. Staats JJ, Plattner BL, Stewart GC, Changappa MM. Presence of the Streptococcus suis suilysin gene and expression of MRP and EF correlates with high virulence in Streptococcus suis type 2 isolates. Vet Microbiol. 1999;70(3-4):201-11.
22. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28(10):2731-9.
23. Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4(4):406-25.
24. Kimura M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol. 1980; 16(2):111-20.
25. Felsenstein J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution. 1985; 39:783-91.
26. MacInnes JI, Desrosiers R. Agents of the “suis-ide diseases” of swine: Actinobacillus suis, Haemophilus parasuis, and Streptococcus suis. Can J Vet Res. 1999; 63(2):83-9.
27. Messier S, Lacouture S, Gottschalk M. Distribution of Streptococcus suis capsular types from 2001 to 2007. Can Vet J. 2008; 49(5):461-2.
28. Dee SA, Corey MM. The survival of Streptococcus suis on farm and veterinary equipment. Swine Health Prod. 1993: 1(1):17-20.
29. Luque I, Blume V, Borge C, Vela AI, Perea JA, Marquez JM, et al. Genetic analysis of Streptococcus suis isolates recovered from diseased and healthy carrier pigs at different stages of production on a pig farm. Vet J. 2010;186(3):396-8.
30. Su Y, Yao W, Perez-Gutierrez ON, Smidt H, Zhu WY. Changes in abundance of Lactobacillus spp. and Streptococcus suis in the stomach, jejunum and ileum of piglets after weaning. FEMS Microbiol Ecol. 2008;66(3):546-55.
31. Princivalli MS, Palmieri C, Magi G, Vignaroli C, Manzin A, Camporese A, et al. Genetic diversity of Streptococcus suis clinical isolates from pigs and humans in Italy (2003-2007). Euro Surveill. 2009;14(33): pii: 19310. Available from: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=19310.
32. Wei Z, Li R, Zhang A, He H, Hua Y, Xia J, et al. Characterization of Streptococcus suis isolates from the diseased pigs in China between 2003 and 2007. Vet Microbiol. 2009;137(1-2):196-201.
33. Tarradas C, Borge C, Arenas A, Maldonado A, Astorga R, Miranda A, et al. Suilysin production by Streptococcus suis strains isolated from diseased and healthy carrier pigs in Spain. Vet Rec. 2001; 148(6):183-4.
34. Smith HE, Vecht U, Wisselink HJ, Stockhofe-Zurwieden N, Biermann Y, Smits MA. Mutants of Streptococcus suis types 1 and 2 impaired in expression of muramidase-released protein and extracellular protein induce disease in newborn germfree pigs. Infect Immun. 1996;64(10):4409-12.
35. Fittipaldi N, Fuller TE, Teel JF, Wilson TL, Wolfram TJ, Lowery DE, et al. Serotype distribution and production of muramidase-released protein, extracellular factor and suilysin by field strains of Streptococcus suis isolated in the United States. Vet Microbiol. 2009;139(3-4):310-7.
36. Fittipaldi N, Xu J, Lacouture S, Tharavichitkul P, Osaki M, Sekizaki T, et al. Lineage and virulence of Streptococcus suis serotype 2 isolates from North America. Emerg Infect Dis. 2011;17(12):2239-44.
37. De Greeff A, Buys H, Verhaar R, Van Alphen L, Smith HE. Distribution of environmentally regulated genes of Streptococcus suis serotype 2 among S. suis serotypes and other organisms. J Clin Microbiol. 2002;40(9):3261-8.
Received in March, 2012.
Accepted in June, 2012.
Ivette Espinosa. División de Biología Molecular, Centro Nacional de Sanidad Agropecuaria, Censa. AP 10, San José de las Lajas, Mayabeque, Cuba. E-mail: espinosa@censa.edu.cu.

{kind=link}
{kind=link}
{kind=link}
{kind=link}