










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.30 no.3 La Habana jul.-set. 2013
RESEARCH
Identification of genes with altered expression levels in contrasting rice cultivars exposed to salt stress treatments
Identificación de genes con niveles de expresión alterados en cultivos contrastantes de arroz expuestos a estrés salino
Mayra Rodríguez1, María C González2, Elizabeth Cristo2, Osvaldo Oliva1, Merardo Pujol1, Orlando Borrás-Hidalgo1
1 Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 11600, La Habana, Cuba.
2 Instituto Nacional de Ciencias Agrícolas, INCA. San José de las Lajas, Mayabeque, CP 10600, Cuba.
ABSTRACT
Soil salinization causes negative effects on plant productivity and poses an increasingly serious threat to the sustainability of agriculture. Improving salt and drought tolerance is one of the most difficult tasks for cereal breeders. A breeding program using somaclonal variation through biotechnological techniques has been used in rice to improve drought and salinity tolerance for increasing agricultural productivity in affected regions. The gene expression of the genes coding for cyclin-dependent kinase C, calcium-dependent protein kinase 7, protein phosphatase, the 7TM-Mlo protein, as well as the expressing sequence tags (EST) EX452034 and EX451286 was determined in rice salt-tolerant somaclone under salt stress conditions. These results show the role of those genes in the salt tolerance performance in rice.
Keywords: Oryza sativa, salt stress, cyclin-dependent kinase C, calcium-dependent kinase 7, phosphatase, seven transmembrane mlo.
RESUMEN
La salinización del suelo provoca efectos negativos en la productividad de las plantas y constituye una amenaza cada vez más grave para la sostenibilidad de la agricultura. Una de las tareas más difíciles para los mejoradores de cereales es aumentar la tolerancia a la salinidad y a la sequía. Durante el programa de mejoramiento del arroz para obtener variedades tolerantes a la salinidad y a la sequía y aumentar su productividad agrícola en las regiones afectadas, se han usado técnicas biotecnológicas empleando la variación somaclonal. Bajo condiciones de salinidad se determinó la expresión de los genes que codifican para la quinasa C dependiente de ciclina, proteína quinasa-7 dependiente de calcio, proteína fosfatasa, la proteína 7TM-Mlo y las secuencias expresadas EX452034 y EX451286 en un somaclón de arroz. Los resultados revelaron la participación de estos genes del arroz en la tolerancia a la salinidad.
Palabras clave: Oryza sativa, estrés salino, quinasa C dependiente de ciclina, proteína quinasa-7 dependiente de calcio, fosfatasa, proteína de transmembrana 7-Mlo.
INTRODUCTION
Rice (Oryza sativa L.) is one of the most important crops in the world and it provides the main resource of energy for more than half of the world population. Rice is planted on about one tenth of the earth’s arable land and is the single largest source of food energy to half of humanity. Salt contamination of the soil is a particular problem in areas with insufficient rain or poor drainage and currently affects almost 20 % of the cultivated and nearly half of the irrigated land worldwide [1]. Cuba, which has an agricultural area of about 7.08 million hectares, has more than a million affected by salinity, and 1.5 million have potential problems of salinization; 55 % of arable soils are classified as saline in the eastern provinces [2]. Plant responses to salt and water stress have much in common. Salinity reduces the ability of plants to take up water. Increasing salt contents in soil water declines the osmotic potential difference and this quickly causes reductions in growth rate,along with a suite of metabolic changes identical to those caused by water stress [3].
Plant adaptation to environmental stresses is controlled by cascades of molecular networks. They activate stress-responsive mechanisms to reestablish homeostasis and to protect and repair damaged proteins and membranes. In contrast to plant resistance to biotic stresses, which is mostly dependent on monogenic traits, the genetically complex responses to abiotic stresses are multigenic and thus more difficult to control and engineer [4]. It is known that the plant stress response is characteristic of highly complex and often integrated signaling pathways. Many drought-inducible genes are also induced by salt stress, which suggests the existence of similar mechanisms of stress responses. Genes induced during drought-stress conditions are thought to function not only in protecting cells from water deficit by the production of important metabolic proteins but also in the regulation of genes for signal transduction in the drought stress response [5].
One of the primary aims of ongoing investigations in the area of rice genomics is to understand gene function and regulatory networks. As well as breeding more stress tolerant varieties of crops, it is important to understand the genetic and molecular basis of the stress response. To this end, the expression of drought inducible genes previously identified by cDNA-AFLP [6] was analyzed in rice under salt stress. In the present work, we determined the gene expression of the encoded genes from rice for O. sativa cyclin-dependent kinase C (OsCDKC), calcium-dependent protein kinase 7 (OsCDPK-7), protein phosphatase (OsPP), 7 transmembrane-Mlo (7TM-Mlo) protein, as well as the expressing sequence tags (EST) EX452034 and EX451286 under salt stress conditions in rice genotypes Amistad-82 (A-82) and INCA LP-7 (LP-7) which develop contrasting salt-stress performances. The Cuban rice variety A-82 has good agronomical traits, however, it is highly susceptible to salt and drought stresses. A-82 has been used as progenitor in biotechnological techniques through somaclonal variation to develop tolerant genotypes to salt and/or drought stresses [7]. LP-7 was obtained from A-82 and is highly tolerant to salinity and good resistance to agronomical traits.
Abiotic stress negatively affects plant growth. Often the reduction in plant size can be attributed to a reduction in cell number, although in many cases, cell growth is also affected. Post proliferative cell enlargement is modulated in response to stress by the plant growth hormone gibberellin. Reduction in cell number has been attributed to inhibition of cyclin-dependent kinase (CDK) activity [8]. Plant growth is a result of cell division and subsequent cell expansion. Cell division is mainly restricted to meristems and is regulated both spatially and temporally in response to growth regulators and environmental signals. The basic regulatory mechanism of the cell cycle is conserved in all higher eukaryotes and consists of a specific family of Ser/Thr kinases, the CDKs. Primarily, CDK activation requires binding with a group of regulatory proteins, the cyclins [9]. Cyclin-dependent kinases are a large family of serine/threonine protein kinases with an important role in ensuring cells progression in an orderly fashion over the different stages of cell division [10].
Calcium-dependent protein kinases (CDPKs) are encoded by multigene family and are involved in signal transduction as important sensors of Ca2+ influx in plants in response to stress. The expression levels of CDPKs genes are spatially and temporally controlled throughout development. In addition, a subset of CDPK genes responds to external stimuli [11]. Transient increases in cytosolic Ca2+ are perceived by various Ca binding proteins. In the case of abiotic stress signaling, evidences suggest that CDPKs sensors are major players in coupling this universal inorganic signal to specific protein phosphorylation cascades. It seems that calcium signaling is crucial for salt tolerance in plants [1]. On the other hand protein phosphatases (PP) participate in signal transduction cascades necessary for cell cycling, development, and disease resistance [12].
The Mlo (mildew resistance locus o) proteins belong to a family of seven transmembrane (7TM) domain proteins located in the plasma membrane. Transcripts for at least half of the six Mlo proteins detected by microarray were induced by NaCl-treatment. These are 7TM proteins with features similar to G-protein coupled receptors, and some members of this gene family have been reported to be abiotic stress responsive [13].
Here we report the first study addressing gene expression of the O. sativa proteins Cyclin dependent kinase (OsCDKC), protein phosphatase (OsPP), calcium-dependent protein kinase 7 (OsCDPK7), EST EX452034 and EX451286, and the 7TM-Mlo protein, under salt stress conditions using a somaclonal cultivar (LP-7) as salt-tolerant rice cultivar. The results allowed us to suggest that the phenotypic variation related with salt tolerance observed in the somaclone is caused by genomic changes. After further confirmatory gene functional experiments, not only we could confirm the changes produced by somaclonal variation but also recommend the use of these genes in breeding programs as either molecular markers or breeding tools through genetic modified rice.
MATERIALS AND METHODS
Salt stress treatment in a greenhouse conditions
Seeds from A-82 and LP-7 were germinated in a greenhouse on floating trays with a mixture of floor-organic matter in a relationship 3:1. Trays were immersed in big rectangle basins containing half-strength Hoagland nutrient solution under outdoor conditions: natural illumination and average day/night temperature of 31 ºC/26 ºC. Nutrient solution was changed every 7 d. After 4 weeks of growth, rice plants were subjected to salt stress by adding NaCl to 150 mM final concentration. Samples of leaves of 5 seedlings were pooled and collected at 0, 7 and 10 days after treatment onset and then frozen in liquid nitrogen and stored at -70 C for further RNA extraction.
Quantitative real-time polymerase chain reaction
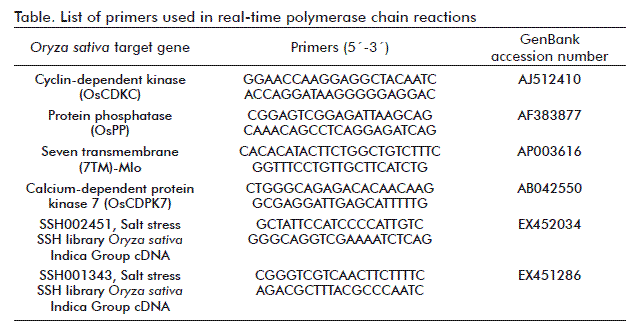
Total RNA (10 mg) was extracted using a commercially available kit using the RNeasy kit (Qiagen, Germany). The integrity of the RNA was checked on agarose gels. The cDNA was synthesized using an oligo(dT) primer and the Super-Script III reverse transcriptase kit (Invitrogen, USA) according to the manufacturer’s instructions. Quantitative real-time polymerase chain reaction (qRT-PCR) was conducted using a Rotor-Gene 3000 PCR machine (Corbett Life Science, Australia) with the QuantiTect SYBR Green PCR kit (Qiagen, Germany). Rice homologies of nucleotide sequences from a selected group of genes were BLAST searched in the NCBI database. Specific primers for OsCDKC, OsCDPK7, OsPP, 7TM-mlo, as well as the EST EX452034 and EX451286 were designed for qRT-PCR with the Primer3 software, version 2000 (Table) [14]. The PCR mixture (Roche Diagnostics, Switzerland) consisted of 0.75 U Tth DNA polymerase, Tth DNA polymerase buffer, 4 mM of MgCl2, 0.4 µM of each primer (one primer set used per assay), 0.2 mM each of dATP, dCTP, dGTP, and dTTP, SYBR Green I diluted 1:10 000 and 4 µL sample. The total volume was 20 µL. Each amplification started with a denaturation step of 15 min at 95 ºC, followed by 40 cycles of 15 s denaturation at 94 ºC, 30 s annealing at 56 ºC and elongation for 30 s at 72 ºC. Each measurement had 3 replicates of the cDNA samples. Actin was used as endogenous gene of reference (Table) forward primer: 5´-GACCCAGATCATGTTTGAGACC-3´; reverse primer: 5´-ATCACCAGAGTCCAACACAATACC-3´). The C value of each gene was normalized with the value of actin to obtain the value of ΔC. Values of ΔΔC relative to control was used to calculate relative gene expression following the procedure described in the relative quantification using the comparative C method in the User Bulletin #2, ABI Prism 7700 Sequence Detection System (Applied Biosystems, Inc., USA).
RESULTS AND DISCUSSION
Salt stress treatment under greenhouse conditions showed the remarkable differences in the behavior of the salt-susceptible variety A-82 and the salt-tolerant somaclone LP-7 under 150 mM NaCl solution. It is well supported by literature that plant growth is greatly affected by environmental abiotic stresses, such as drought and high salinity. The somaclone LP-7 showed high tolerance to salinity in greenhouses condition (not shown).
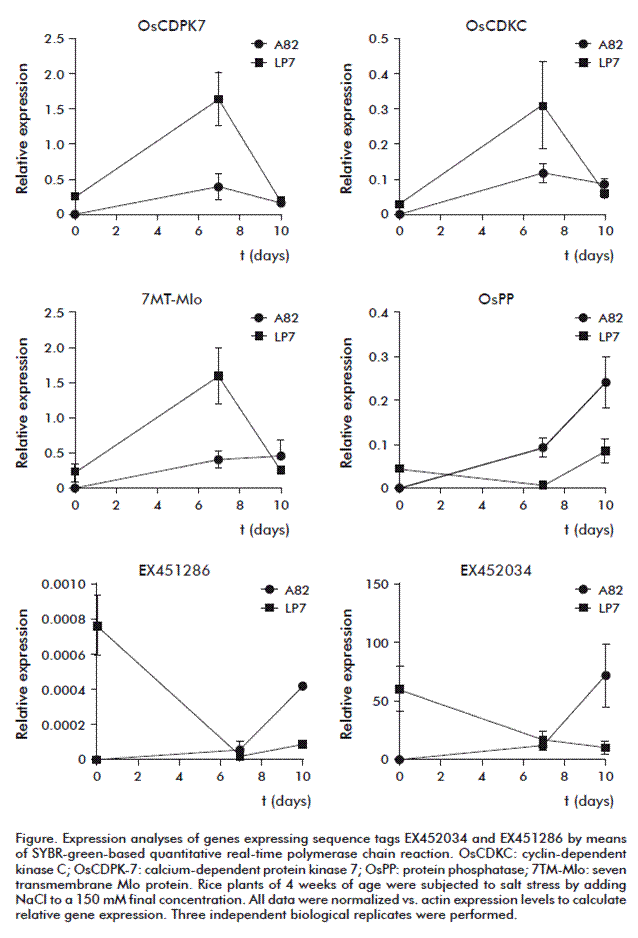
To analyze the expression of some drought stress-up regulated genes under salinity stress conditions we used the above mentioned genotypes because of their contrasting salt-stress behavior. We identified the corresponding genes in rice by searching each TDF against all the sequences in the non-redundant databases using the BlastN algorithms (National Center for Biotechnology Information, USA). The expression of the genes OsCDPK7, OsCDKC, OsPP, 7TM-mlo, EST EX452034 and EST EX451286 were quantified by qRT-PCR (Figure).
OsCDPK7, OsCDKC and 7TM-mlo genes from the cultivar LP-7 increased their relative expression until day seven after salt stress, followed by a silencing of the gene expression at 10 days, when the expression values dropped down to almost baseline. In the variety A-82, the increase of expression at day seven was lower than in somaclone LP-7, followed by a decrease after 10 days except for the 7TM-mlo gene expression, which had a little increase. These results suggested that OsCDKC, 7TM-mlo and OsCDPK7 genes are involved in the tolerance response to salinity in the salt-tolerant rice somaclone LP-7.
The first detectable response to sodium stress is a rise in the cytosolic-free calcium concentration. This calcium signal serves as a second messenger that turns on the machinery for sodium export and potassium/sodium discrimination. Calmodulins (CaM) and CDPKs belong to the major families of calcium-binding proteins. There is an interaction between the CaM and Mlo proteins. Rice Mlo proteins contain a CaM-binding domain in a region of the carboxy-terminal cytoplasmic tail that is relatively conserved in sequence among the Mlo family members. Mlo proteins have a broad involvement in cell death protection and in responses to biotic and abiotic stresses [15]. The 7TM-Mlo protein was induced both in a drought-tolerant rice variety during osmotic stress [6] as well as under salinity conditions in a salt-tolerant rice somaclone. Our results support the hypothesis that induction of 7TM-Mlo proteins increases their interaction with CaM activating the signaling cascade in which calcium is a messenger molecule.
According to these results, the salt-tolerant somaclone showed an induction of OsCDPK7 after salt stress treatment, suggesting a function in the salt stress signaling pathways. CDPKs have a characteristic structure in which an N-terminal serine/threonine protein kinase domain is fused to a carboxyterminal CaM-like domain containing EF-hand calcium-binding sites. Therefore, CDPKs do not depend on the interaction with exogenous CaM but can be activated directly by Ca2+ binding certain CDPK signaling pathways [16].
Different members of the CDK have diverse roles in cell cycle progression and in regulating gene expression [17]. A CDKC type from rice (CDKC1) was proposed to be involved in developmental events as well as in the plant salt stress response mechanism through an ABA-signaling pathway. Elevated NaCl also triggered increased expression of CDKC1 in cell suspensions. This suggested that CDKC1 expression might be a primary response to salt stress and could participate in the transcriptional regulation of salt-induced genes for the induction of defense and/or tolerance in a saline environment [18]. Although the molecular interactions that link the cell cycle machinery to perception of stress are not fully understood, recent studies indicated the involvement of CDKs in the plant stress response machinery. CDKs are core cell cycle regulators but their activity has been implicated in additional diverse cellular processes [8].
The OsPP gene showed a gradually increase of expression until day 10 under salinity conditions in the susceptible variety. However, a little increase of the expression from 7 to 10 day was observed in the somaclone LP-7 but this remained lower when compared to the variety A-82. Studies using phosphatase inhibitors have indicated a role for phosphatases in stress signaling, where the activation and inactivation of kinases is regulated by phosphorylation and dephosphorylation events, respectively [12].
The expression under salinity conditions of the studied EST (EX452034 and EX451286) was similar. We found in the somaclone LP-7 a decreased expression of both genes from a higher initial point to almost cero after 10 days, showing silencing of these genes. Otherwise, an increase of its expression during 10 days of salt treatment was observed in the variety A-82, showing an induction after 7 days of salt stress. It has been reported that EST EX452034 and EX451286 were differentially expressed under osmotic and salt stress in rice [19]. This behavior corresponded with the variety A-82; however, in the tolerant genotype LP-7 a down regulation of both ESTs was observed.
Noteworthy, expression profiling has become an important tool to investigate how an organism responds to environmental changes. Plants have the ability to alter their gene expression patterns in response to environmental changes such as temperature, water availability or the presence of deleterious levels of ions. Sometimes these transcriptional changes are successful adaptations leading to tolerance while at other times the plant ultimately fails to adapt to the new environment and is considered to be sensitive to that condition [20]. In this sense, expression profiling can define both tolerant and sensitive responses. These profiles of plant response to environmental extreme conditions are expected to lead to identify regulators that will be useful in biotechnological approaches to improve stress tolerance as well as to new tools for studying regulatory genetic circuitry in plants under different types of stress.
All together, the genomic differences between susceptible- and tolerant-rice genotypes were evidenced, by the differential expression of the genes coding for OsCDKC, OsCDPK7, OsPP, 7TM-Mlo protein, as well as the ESTs EX452034 and EX451286 under salt stress conditions in rice genotypes with contrasting salt-stress performance. These results show the role of OsCDKC, OsCDPK7 and 7TM-mlo genes in the salt tolerance performance in rice and allow us to suggest their function in the primary response to salt stress as targets of stress responsive signaling cascades. Therefore, we recommend the use of these genes in breeding programs as either molecular markers or for breeding through genetically-modified rice. The functions of stress-inducible genes not only to understand the molecular mechanisms of stress tolerance and the responses of higher plants, but also to improve the stress tolerance of crops by gene manipulation must be also considered.
REFERENCES
1. Xiong L, Schumaker KS, Zhu JK. Cell signaling during cold, drought, and salt stress. Plant Cell. 2002;14 Suppl:S165-83.
2. González LM, Ramírez R. Los suelos salinos y su utilización en la producción agrícola. Alimentaria. 2002;339:103-7.
3. Munns R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002;25(2):239-50.
4. Wang W, Vinocur B, Altman A. Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta. 2003;218(1):1-14.
5. Shinozaki K, Yamaguchi-Shinozaki K, Seki M. Regulatory network of gene expression in the drought and cold stress responses. Curr Opin Plant Biol. 2003;6(5):410-7.
6. Rodríguez M, Canales E, Borroto CJ, Carmona E, López J, Pujol M, et al. Identification of genes induced upon water-deficit stress in a drought-tolerant rice cultivar. J Plant Physiol. 2006;163(5):577-84.
7. González MC, Pérez N, Cristo E, Rodríguez M, Borrás O. Development of salinity tolerant-rice varieties using biotechnological nuclear techniques. In: Shu QY, editor. Induced plant mutations in the genomics era. Rome: Food and Agriculture Organization of the United Nations (FAO). 2009; p. 138-40.
8. Kitsios G, Doonan JH. Cyclin dependent protein kinases and stress responses in plants. Plant Signal Behav. 2011;6(2):204-9.
9. Stals H, Inze D. When plant cells decide to divide. Trends Plant Sci. 2001;6(8):359-64.
10. John PC, Mews M, Moore R. Cyclin/Cdk complexes: their involvement in cell cycle progression and mitotic division. Protoplasma. 2001;216(3-4):119-42.
11. Serrano R, Gaxiola R, Rios G, Forment J, Vicente O, Ros R. Salt stress proteins identified by a functional approach in yeast. Monatshefte fur Chemie. 2003;134(11):1445-64.
12. Takahashi K, Isobe M, Muto S. An increase in cytosolic calcium ion concentration precedes hypo-osmotic shock-induced activation of protein kinases in tobacco suspension culture cells. FEBS Lett. 1997;401(2-3):202-6.
13. Jiang Y, Deyholos MK. Comprehensive transcriptional profiling of NaCl-stressed Arabidopsis roots reveals novel classes of responsive genes. BMC Plant Biol. 2006;6:25.
14. Rozen S, Skaletsky HJ. Primer3 on the WWW for general users and for biologist programmers. Methods Mol Biol. 2000;132:365-86.
15. Kim MC, Panstruga R, Elliot C, Muller J, Devoto A, Yoon HW, et al. Calmodulin interacts with Mlo protein to regulate defense against mildew in barley. Nature. 2002;416(6879):447-50.
16. Cheng SH, Willmann MR, Chen HC, Sheen J. Calcium signalling through protein kinases. The Arabidopsis calcium dependent protein kinase gene family. Plant Physiol. 2002;129(2):469-85.
17. West G, Inze D, Beemster GT. Cell cycle modulation in the response of the primary root of Arabidopsis to salt stress. Plant Physiol. 2004;135(2):1050-8.
18. Huang YW, Tsay WS, Chen CC, Lin CW, Huang HJ. Increased expression of the rice C-type cyclin-dependent protein kinase gene, Orysa;CDKC;1, in response to salt stress. Plant Physiol Biochem. 2008;46(1):71-81.
19. EX449804. SSH00339 Osmotic stress SSH library Oryza sativa Japonica Group cDNA, mRNA sequence [Internet]. Winden: MetaLife AG. c2013 [cited 2013 Jan 18]. Available from: http://www.metalife.com/Genbank/157650170.
20. Hazen SP, Wu Y, Kreps JA. Gene expression profiling of plant responses to abiotic stress. Funct Integr Genomics. 2003;3(3):105-11.
Received in January, 2013.
Accepted in March, 2013.
Mayra Rodríguez. Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 11600, La Habana, Cuba. E-mail: mayra.rodriguez@cigb.edu.cu.


{kind=link}
{kind=link}