










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.30 no.3 La Habana jul.-set. 2013
RESEARCH
Molecular and biochemical characterization of tomato (Solanum lycopersicum L.) plants cv. Micro-Tom under lead (Pb)-induced stress
Caracterización molecular y bioquímica de plantas de tomate (Solanum lycopersicum L.) cv. Micro-Tom sometidas a estrés inducido por plomo (Pb)
Sandra Pérez1, Ahmed I S Ahmed2, Daniel Cabezas1
1 Departamento de Biología y Sanidad Vegetal, Universidad Agraria de La Habana Fructuoso Rodríguez Pérez. Carretera Tapaste, Km 22 ½, San José de las Lajas, Mayabeque, Cuba.
2 Plant Pathology Unit, Plant Protection Department, Desert Research Center, Cairo, Egypt.
ABSTRACT
Two different Pb concentrations (5 and 10 mg/kg of PbAc2) were used to study the response of Solanum lycopersicum L. cv. Micro-Tom (MT) plants in two vegetative growth phases. Two genes that have been previously reported to be heavy metal inducible genes (superoxide dismutase, SOD, EC 1.15.1.1; and isoflavone reductase, IFR; EC 1.6.4.2) and another one putatively interesting in this field (transcriptionally controlled tumor protein, TCTP) were selected for an expression study using a real time PCR technique. In the first growth phase (germination to flowering) TCTP was repressed at 10 mg/kg and in the control, SOD expression was low at both concentrations (5 and 10 mg/kg of PbAc2) and IFR was higher at 10 mg/kg PbAc2. In the second phase (flowering to fructification) three genes were expressed in both concentrations but in the case of TCTP and SOD the expression was higher at 5 mg/kg of PbAc2. SOD and Glutathione reductase (GR; EC 1.6.4.2) were selected for a biochemical study together with the determination of protein concentration using spectrophotometer. SOD was higher at 10 mg/kg PbAc2 showing significant difference with control, and GR had the same behavior but had significant differences with the other two treatments while total proteins were higher for the control showing significant differences at 10 mg/kg of PbAc2. This research suggests that Pb toxicity leads to the induction of key enzymes of antioxidant defense system in tomato plants.
Keywords: tomato, heavy metals, expression study, molecular mechanism, abiotic stress, lead.
RESUMEN
Se estudió la respuesta de plantas de tomate Solanum lycopersicum L. cv. Micro-Tom (MT) sometidas a estrés inducido por plomo (5 y 10 mg/kg de PbAc2) y en las fases de desarrollo vegetativo germinación-floración y floración-fructificación. Se determinaron los niveles de expresión en estas condiciones de los genes inducidos por metales pesados superóxido dismutasa (SOD; EC 1.15.1.1) e isoflavona reductasa (IFR; EC 1.6.4.2), y otro de potencial interés en este campo (proteína tumoral controlada durante la transcripción-TCTP). En la primera fase, la TCTP fue reprimida en el grupo control y a la máxima concentración empleada de PbAc2, la expresión de la SOD fue baja en las dos concentraciones y la IFR fue mayor a 10 mg/kg. En la segunda fase, los tres genes se expresaron en las dos concentraciones, la TCTP y la SOD a mayores niveles en 5 mg/kg de PbAc2. Se determinó por densidad óptica la actividad específica de SOD y de la glutatión reductasa (GR; EC 1.6.4.2), junto con la concentración de proteínas totales. Ambas enzimas antioxidantes mostraron mayor actividad específica a 10 mg/kg de PbAc2, con diferencias estadísticamente significativas con respecto al tratamiento control, y la GR también mostró diferencias estadísticamente significativas con respecto al tratamiento con 5 mg/kg. La concentración de proteínas totales fue mayor en el grupo control, y estadísticamente significativa con respecto a la del tratamiento con 10 mg/kg. Esta investigación sugiere que la toxicidad por plomo provoca la inducción de enzimas antioxidantes claves del sistema de defensa en las plantas de tomate.
Palabras clave: tomate, metales pesados, estudio de expresión, mecanismo molecular, estrés abiótico, plomo.
INTRODUCTION
The Solanaceae family includes several species of agronomic importance, such as tomato (Solanum lycopersicum L.), potato (Solanum tuberosum L.), and tobacco (Nicotiana tabacum L.). Tomato is one of the most important vegetable crops worldwide, with a total production of around 141 million tons on a cultivated area of around 5 million hectares. This crop represents also one of the major products of the food industry worldwide [1]. Additionally, tomato is an important plant model to numerous studies of genetics [2], biochemistry [3], morphology and anatomy [4], mutagenesis [5], and others.
Particularly, tomato has been used in numerous studies as a hyper accumulator plant to decontaminate soils with high metal concentration [6, 7], as one of the approximately 400 plant species from at least 45 plant families that have been reported to hyperaccumulate metals [8]. The high concentration of metal in soil is a severe detrimental factor among those causing abiotic stresses which affect crop production [9, 10]. One of the most common heavy metal contaminants in the environment is lead (Pb), widely spread by activities such as mining, smelting, and dumping of municipal sewage wastes which have polluted extensive areas throughout the world [11]. Pb is not an essential element in the metabolic processes in plants or animals, and it can accumulate to levels enough to become toxic or lethal to living organisms [10].
Studies carried out in plants have shown that Pb is mainly absorbed in soil, affecting mineral nutrient absorption, growth and metabolic processes such as photosynthesis, respiration, and cell division in germinating seeds of various plant species [12]. Excessive stress by heavy metals, including Pb, causes oxidative damage, but some reactive oxygen species (ROS) can participate in signal transduction pathways [13].
To get more knowledge of the effect of Pb in vegetable crops, as one of the first links in the food chain, the expression profiles of three genes involved in Pb-stress in tomato (using the model cultivar Micro-Tom [14]) were analyzed: superoxide dismutase (SOD), the transcriptionally-controlled tumor protein (TCTP) and the isoflavone reductase (IFR).
MATERIALS AND METHODS
Plant material
The Micro-Tom cultivar of tomato (S. lycopersicum) was kindly provided by Dr A Levy (Weizmann Institute of Science, Rehovot, Israel). Seeds were sown in boxes containing a mixture of commercial pot mix (Basaplant®, São Paulo, Brazil) and vermiculite (1:1 v/v), and supplemented with 1 g of NPK (Nitrogen:Phosphorus:Potassium, 10:10:10 v/v) and 4 g of lime per liter of mixture. After the first true leaves appeared, seedlings were transplanted to 1-L pots filled with sand. One control and two different Pb treatments (5 and 10 mg of PbAc2/kg of soil, respectively) with three replicates each were established. The Pb solutions were applied to tomato plants twice a week. Leaves were collected in tomato flowering and fructification, and stored at -80 ºC for further analysis.
Molecular procedures
Total RNA isolation
Total RNA was extracted using the InviTrap® Plant RNA Mini Kit. The RNA concentration was determined using a Nano-Drop Spectrophotometer prior a complementary DNA (cDNA) experiment.
cDNA first strand synthesis
The first strand of cDNA was synthesized using a SuperScriptTM first strand synthesis system (Invitrogen Life Technologies, USA). A polymerase chain reaction (PCR) of three sequential steps of one cycle each was performed at 42 ºC for 5 min, 50 ºC for 50 min and another at 70 ºC for 15 min, respectively.
Analysis of the expression levels by real time-PCR
The differential expression of genes coding for proteins widely involved in stress response (SOD and IFR) and a metal metabolism enzyme (TCTP) was analyzed using real time-PCR (RT-PCR). The primers were designed using Primer Express 2.00 (Applied Biosystems software), based on sequences retrieved from the National Center of Biotechnology Information (NCBI) database. In the case of TCTP gene, it was selected from a cDNA tobacco library [15], its role starting to be reported as relevant in metal metabolism.
RT-PCR was performed using a Platinum® SYBR® Green qPCR Super MIX-UDG (Invitrogen), with 48-wells plates and the standard cycling program. Amplification comprised one cycle at 95 ºC for 15 min, 40 cycles at 95 ºC for 15 min, 60 ºC for 60 min, followed by one cycle at 95 ºC for 15 min and 60 ºC for 60 min. Data was analyzed by comparative quantification CT (ΔΔCT) with the StepOne software.
Biochemical procedures
Enzyme extraction and assays
The following steps were carried out at 4 ºC otherwise stated. Tomato (S. lycopersicum) samples were homogenized (2:1 buffer volume:fresh weight) in a mortar with pestle with 100 mM potassium phosphate buffer (pH 7.5) containing 1 mM ethylenediamine tetraacetic acid (EDTA), 3 mM DL-dithiothreitol and 5 % (w/v) insoluble PVPP. The homogenate was centrifuged at 10 000 × g for 30 min and the supernatant was kept stored in separate aliquots at -80 ºC, prior to glutathion reductase (GR) and SOD analyses. Total GR activity was determined in a spectrophotometer as described by Azevedo [16]. The rate of reduction oxidized glutathione (GSSG) was followed by monitoring the increase in absorbance at 412 nm for 2 min [9]. GR activity is expressed as µmol/min per milligram of protein. SOD activity was measured by optical density at 560 nm [17], based on the inhibition of the photochemical reduction of nitroblue tetrazolium (NBT). One unit of enzyme activity was defined as the amount of SOD required for 50 % inhibition of NBT reduction.
Determination of protein concentration
Protein concentration was determined by the Bradford’s method [18] using bovine albumin serum (BSA).
Statistics
Data variability and validity of the results were confirmed by applying the Duncan’s test at a confidence interval of 95 %.
RESULTS AND DISCUSSION
Growth reduction was observed in tomato plants of cv Micro-Tom under Pb-stress (Figure 1). Chlorosis and necrotic lesions appeared after 35 days of growth, indicating altered mineral nutrient absorption and photosynthesis.
Tomato plants under Pb-stress for 35 days at the highest concentration of 10 mg/kg showed lost almost all the leaves, with reduction in growth length. Similar results have been reported by studies in Brassica juncea L. [19], Zea mays [20], S. lycopersicum [21], Phaseolus vulgaris and Lens culinaris [22], also reporting delayed development, low quality harvests and decreased yields [23].
The response to heavy metal stress involves a complicated signal transduction network that is activated by sensing the heavy metal, followed by the synthesis of stress-related proteins and signaling molecules, which ultimately activate transcription of specific metal-responsive genes to counteract the stress [24]. The relevant signal transduction pathways include hormones, ROS signaling, and the activation of some stress-related genes. Different signaling pathways may be used to respond to different heavy metals [25].
Stress at cellular level results in excessive production of ROS, causing lipid peroxidation and damage to several enzymes. On the other hand, cells mechanism can modulate stress responses by antioxidant enzymes such as SOD, GR, Catalase (CAT) and Ascorbate peroxidase (Asc) [26].
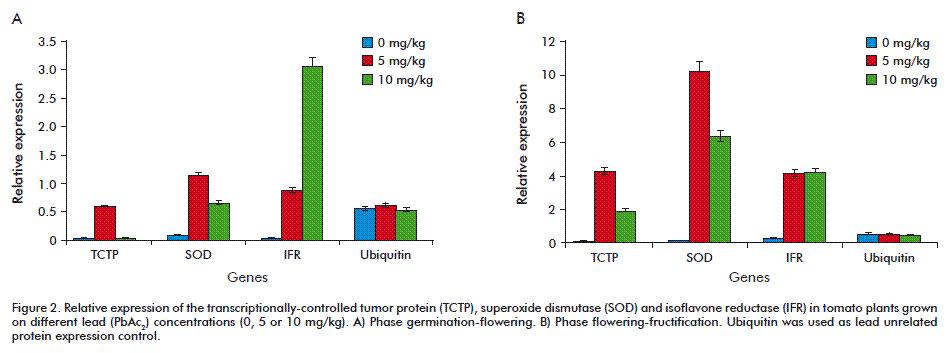
SOD, TCTP and IFR genes were studied to know their behavior at molecular and biochemical level in tomato plants cv. Micro-Tom under Pb stress. The three genes (SOD, TCTP and IFR) showed different expression profiles under the tested concentrations and depending on the collection phase (Figure 2). Ubiquitin was used as control gene due to its basal stable expression, as commonly used in different crops, with the ubiquitin-conjugating enzyme and elongation factor-1 regarded as the most stable based on their transcriptional profiles in Oryza sativa and Pennisetum ciliare [27, 28].
In the case of TCTP, it showed low expression at 5 mg/kg of PbAc2, which was repressed at 10 mg/kg of PbAc2 during the germination and flowering phase (first phase), while showing low expression at both concentrations during flowering-fructification (second phase) (Figure 2). This could be related to a downregulation on TCTP expression by Pb [29-31]. It is also in agreement with previous results [32] showing that the developmental phase influences TCTP expression levels, together with the cell/tissue type and the stress conditions the plants are subjected to [33, 34]. This is irrespective of the ubiquitous distribution of TCTP in all the eukaryotic organisms, in more than 500 tissues and cell types, and the numerous experimental settings and biological systems tested. It has been established that TCTP levels are highly regulated in response to a wide range of extracellular signals and cellular conditions. Typically, growth signals [35] have been reported to rapidly induce TCTP synthesis. That’s why TCTP induction may be repressed at reduced plant growth (Figure 1C and D).
There are some contradictory reports as those of Schmidt et al. [36] who found that some heavy metals in plants up-regulate TCTP under various cells stress conditions, and by Gnanasekar and Ramaswamy [37] showing that TCTP plays some antioxidant functions.
In most of the cases up-regulation of cell growth-related genes is presumed to be involved in metal stress signaling response [38], but in our study TCTP in relation with heavy metal induced stress is not as highly expressed as it would be expected, and it is down-regulated, showing a relationship with plant growth. There is limited information about plant TCTPs so this study is one of the first studies involving TCTPs in plants related to Pb induced stress.
Adaptation of plants to several types of stresses depends upon a complex cellular sign system where ROS, salicylates, cellulose, chitin oligomers can intervene [39, 40]. These species (ROS) can react with cellular components (lipids, proteins, nucleic acids) and cause lipid peroxidation, membrane damage and inactivation of enzymes. Plants have evolved a complex array of mechanisms to maintain low ROS level and avoid the detrimental effects of excessively high ROS concentrations. This antioxidant network includes numerous soluble (ascorbate, glutathione) and membrane compounds (tocopherol) as well as enzymes involved in ROS scavenging (SOD, CAT, Asc) [41].
SOD expression was lower in the first phase for both Pb treatments and higher in the second phase, specifically at 5 mg/kg of PbAc2. A possible explanation for this is the increased production of superoxide radicals due to the rise in ROS production by Pb toxicity. The higher expression of SOD at 5 mg/kg of PbAc2 in the second phase, as for other induced enzymes, may be caused by the possible temporal expression of SOD, mainly transitional. However, this response also may be a consequence of SOD-related transcriptional activity genes. So, in our study, SOD expression was higher at the lower heavy metal concentration, which is in agree with results of Jomová and Morovič [42] who found that lower concentrations of Pb (100, 200 mg/L) in Lupinus luteus L. induced a strong increase of SOD activity, but further elevation on its concentration (300, 400, 500 mg/kg) had the inverse effect. A lower expression at 10 mg/kg of PbAc2 may suggest that higher concentrations could damage plants, which is suggested by the fact that the growth of plants at this concentration was affected. Similar results were obtained in Arabidopsis [43], where the expression of four genes encoding superoxidase enzymes were induced in response to Pb treatment, suggesting that Pb, like other heavy metals ions, activate specific responses to the plant antioxidant defense system. The expression of SOD at both Pb concentrations used in this research indicates that it may be involved in the antioxidative process under Pb induced stresses, since SOD is considered to be a crucial component in biological defense against oxidative stress [44].
The induction of IFR was higher at 10 mg/kg of PbAc2 in the first phase, and, in the second phase (flowering and fructification), its expression was similar for both concentrations tested. A higher expression of IFR at 10 mg/kg of PbAc2 at germination-flowering phase is probably due to the adaptation of plants to a new environment (sand) with an application of a toxic metal on this phase. At this point tomato plant may produce lignin which functions and distribution suggest that lignins are a group of the earliest forms of defense mechanism and they are an important aromatic secondary metabolites produced in the phenylpropanoid pathway [45]. With respect to IFR expression it was higher at 10 mg/kg of PbAc2 in the first phase and in the second was similar at both tested concentrations. Similar results with cadmium in Linum usitatissimum L. showed that higher concentration (50 and 100 mM) of this heavy metal induced IFR expression [46].
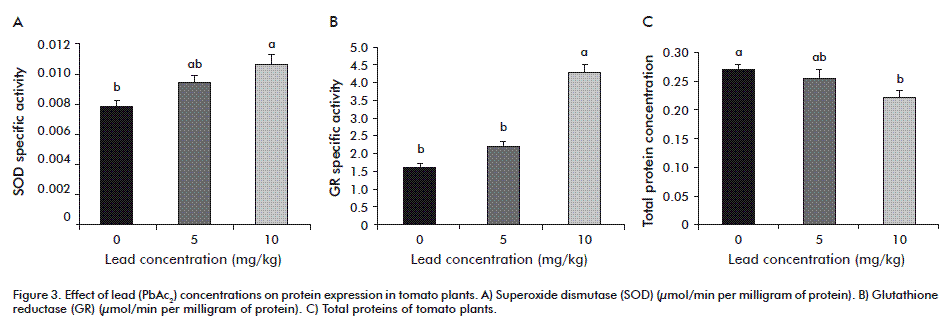
The biochemical analysis showed differences between SOD (µmol/min per milligram of protein FW), GR (µmol/min per milligram of protein FW) and total proteins (Figure 3).
During the experimental period, the content of SOD was higher at 10 mg/kg of PbAc2 with significant differences with respect to the control and 5 mg/kg of PbAc2. This is also corroborated by the fact that plant growth at this concentration was really affected. Plants exposed to Pb stress also show rapid and temporary drops in growth rate and activate antioxidant defense system by producing ROS, which alters gene expression and enzyme activity patterns of SOD.
Similar results were found in tomato plants under cadmium induced stress [47] and, as the results obtained in this research, suggested that SOD activity might play a crucial role in the response of tomato plants to metallic stress [48]. Accumulation of SOD, indicating the oxidative stress, is related to the maintenance of the overall defense system. The activation of SOD could be useful to reduce O2 accumulation, decrease of hydrogen peroxide and alleviate Pb stress [49].
GR content was higher also at 10 mg/kg of PbAc2, with significant differences with respect to the other two treatments. This enzyme is part of the defenses system against oxidative stress. GRs are indispensable components of ascorbate-glutathione pathway, required to scavenge H2O2 produced mainly in chloroplasts and other cell organelles and to maintain the redox state of the cell [50]. Glutathione reductase catalyzes the NADPH-dependent GSSG to reduced GSH that is involved in the redox regulation of the cell cycle and has often been considered to play an important role in defense of plants and other organisms against oxidative stress [51].
This results show increased GR activity in Pb treated tomato plants, which suggests possible involvement of GR in regenerating GSH under Pb toxicity conditions to increase GSH/GSSG ratio and the total glutathione pool. Similar to our results, induction in GR activity has been reported in leaves of tomato plants under cadmium induced stress [47] and in rice seedling under Pb induced stress [52]. Total proteins were higher in the absence of PbAc2 (control), which means there could be degraded with the other two concentrations used.
Production of ROS takes place in cell under normal conditions, however adverse environmental conditions that interrupt cellular homeostasis could produce oxidative damage to proteins, DNA and to the lipids [53]. This could explain why total proteins decreased at higher concentrations of Pb.
An important role in plants adaptation and surviving under stress conditions is played by antioxidants enzymes in oxidative stress tolerance [50].
In summary, the present research suggests that Pb toxicity in situ leads to production of lipid peroxides and induces some of the key enzymes of antioxidant defense system in tomato plants. Induction in the activities of antioxidative enzymes is a general strategy adopted by plants to overcome oxidative stress due to the imposition of environmental stresses.
ACKNOWLEDGEMENTS
We are grateful to The Academy of Sciences for Developing Word (TWAS) and CNPq for providing the opportunity to do this research at Sao Paulo University/Agricultural School Luiz de Queiros, Brazil.
REFERENCES
1. FAOSTAT. The statistics division of the FAO. Roma: FAO. c2012 [cited 2013 Jan 15]. Available from: http://faostat.fao.org.
2. Butelli E, Titta L, Giorgio M, Mock HP, Matros A, Peterek S, et al. Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nat Biotechnol. 2008;26(11):1301-08.
3. Gratão PL, Monteiro CC, Carvalho RF, Tezotto T, Piotto FA, Peres LEP, et al. Biochemical dissection of diageotropica and Never ripe tomato mutants to Cd-stressful conditions. Plant Physiol Biochem. 2012;56:79-96.
4. Gratão PL, Monteiro CC, Rossi ML, Martinelli AP, Peres LEP, Medici LO, et al. Differential ultrastructural changes in tomato hormonal mutants exposed to cadmium. Environ Exp Bot. 2009;67(2):387-94.
5. Pino-Nunes LE, Figueira de O AV, Tulmann-Neto A, Zsögön A, Piotto FA, Silva JA, et al. Induced mutagenesis and natural genetic variation in tomato 'Micro-Tom'. Acta Hort (ISHS). 2009;821:63-72.
6. Kumar N, Soni H, Kumar RN, Bhatt I. Hyperaccumulation and mobility of heavy metals in vegetable crops in India. J Agr Environ. 2009;10:29-38.
7. He LY, Chen ZJ, Ren GD, Zhang YF, Qian M, Sheng XF. Increased cadmium and lead uptake of a cadmium hyperaccumulator tomato by cadmium-resistant bacteria. Ecotoxicol Environ Saf. 2009;72(5):1343-8.
8. Al-Qurainy F. Toxicity of heavy metals and their molecular detection on Phaseolus vulgaris (L.). Aust J Basic Appl Sci. 2009;3(3):3025-35.
9. Gomes-Junior RA, Moldes CA, Delite FS, Pompeu GB, Gratão PL, Mazzafera P, et al. Antioxidant metabolism of coffee cell suspension cultures in response to cadmium. Chemosphere. 2006;65(8):1330-7.
10. Chen F, Wang F, Wu F, Mao W, Zhang G, Zhou M. Modulation of exogenous glutathione in antioxidant defense system against Cd stress in the two barley genotypes differing in Cd tolerance. Plant Physiol Biochem. 2010;48(8):663-72.
11. Cui YS, Wang QR, Dong YT, Li HF, Christie P. Enhanced uptake of soil Pb and Zn by Indian mustard and winter wheat following combined soil application of elemental sulphur and EDTA. Plant Soil. 2004;261(1-2):181-8.
12. Sharma P, Dubey RS. Lead toxicity in plants. Braz J Plant Physiol. 2005;17(1):35-52.
13. Mittler R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002;7(9):405-10.
14. Meissner R, Jacobson Y, Melamed S, Levyatuv S, Shalev G, Ashri A, et al. A new model system for tomato genetics. Plant J. 1997;12(6):1465-72.
15. Pérez SP, Cabezas DM, Haitao D. Large-scale identification of ESTs from Nicotiana tabacum L. by normalized cDNA library sequencing. Beitrage zur Tabakforschung Int. 2006;22(2):114-24.
16. Azevedo R, Alas R, Smith R, Lea P. Response of antioxidant enzymes to transfer from elevated carbon dioxide to air and ozone fumigation, in the leaves and roots of wild-type and a catalase-deficient mutant of barley. Physiol Plant. 1998;104(2):280-92.
17. Beyer WF, Fridovich I. Assaying for superoxide dismutase activity: some large consequences of minor changes in conditions. Anal Biochem. 1987;161(2):559-66.
18. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem. 1976;72:248-54.
19. John R, Ahmad P, Gadgil K, Sharma S. Heavy metal toxicity: Effect on plant growth, biochemical parameters and metal accumulation by Brassica juncea L. Int J Plant Prod. 2009;3(3):65-76.
20. Çimrin KM, Turan M, Kapur B. Effect of elemental sulphur on heavy metals solubility and remediation by plants in calcareous soils. Fresenius Environ Bull. 2007;16(9a):1113-20.
21. Akinci IE, Akinci S, Yilmaz K. Response of tomato (Solanum lycopersicum L.) to lead toxicity: Growth, element uptake, chlorophyll and water content. Afr J Agri Res. 2010;5(6):416-23.
22. Haider S, Kanwal S, Uddin F, Azmat R. Phytotoxicity of Pb II. Changes in chlorophyll absorption spectrum due to toxic metal Pb stress on Phaseolus mungo and Lens culinaris. Pakistan J Biol Sci. 2006;9(11):2062-8.
23. Begum A. Cultivation of tomato plants (Lycopersicum esculentum) using Municipal Solid Waste Compost. Int Soc Environ Bot. 2010;16(2):850-63.
24. Maksymiec W. Signaling responses in plants to heavy metal stress. Acta Physiol Plant. 2007;29(3):177-87.
25. DalCorso G, Farinati S, Furini A. Regulatory networks of cadmium stress in plants. Plant Signal Behavior. 2010;5(6):663-7.
26. Gratão PL, Monteiro CC, Peres LEP, Azevedo RA. The Isolation of Antioxidant Enzymes from Mature Tomato (cv. Micro-Tom). Plant Hort Sci. 2008;43:1608-10.
27. Jain M, Nijhawan A, Tyagi AK, Khurana JP. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem Biophys Res Commun. 2006;345(2):646-51.
28. Singh M, Burson B, Finlayson S. Isolation of candidate genes for apomictic development in buffelgrass (Pennisetum ciliare). Plant Mol Biol. 2007;64(6):673-82.
29. Azcón-Bieto J, Talón M. Fundamentos de Fisiología Vegetal. 2nd ed. Madrid: McGraw-Hill Interamericana; 2008.
30. Kawaguchi E, Gyokusen K, Saito A. Appearance of partially wilted shoots in a Pinus thunbergii inoculated with Bursaphelenchus xylophilus and its influence on the growth of the tree. Bull Kyushu Univ For. 1999;80:41-9.
31. Berkowitz O, Jost R, Pollmann S, Masle J. Characterization of TCTP, the Translationally Controlled Tumor Protein, from Arabidopsis thaliana. Plant Cell. 2008;20(12):3430-47.
32. Rao KV, Chen L, Gnanasekar M, Ramaswamy K. Cloning and characterization of a calcium-binding, histamine-releasing protein from Schistosoma mansoni. J Biol Chem. 2002;277(34):31207-13.
33. Thiele H, Berger M, Skalweit A, Thiele BJ. Expression of the gene and processed pseudogenes encoding the human and rabbit translationally controlled tumour protein (TCTP). Eur J Biochem. 2000;267(17):5473-81.
34. Bommer UA, Thiele BJ. The translationally controlled tumour protein (TCTP). Int J Biochem Cell Biol. 2004;36(3):379-85.
35. Bommer, UA, Heng C, Perrin A, Dash P, Lobov S, Elia A, et al. Roles of the transnationally controlled tumor protein (TCTP) and the double stranded RNA-dependent protein kinase, PKR, in cellular stress response. Oncogene. 2010;29(5):763-73.
36. Schmidt I, Fahling M, Nafz B, Skalweit A, Thiele BJ. Induction of translationally controlled tumor protein (TCTP) by transcriptional and post-transcriptional mechanisms. FEBS J. 2007;274(20):5416-24.
37. Gnanasekar M, Ramaswamy K. Translationally controlled tumor protein of Brugia malayi fuctions as an antioxidant protein. Parasitol Res. 2007;101(6):1533-40.
38. Ermolayev V, Weschke W, Manteuffel R. Comparison of Al induced gene expression in sensitive and tolerant soybean cultivars. J Exp Bot. 2003;54(393):2745-56.
39. Buchel AS, Brederode FT, Bol JF, Linthorst HJ. Mutation of GT-1 binding sites in the PR-1a promoter influences the level of inducible gene expression in vivo. Plant Mol Biol. 1999;40(3):387-96.
40. Knoester M, Pieterse CM, Bol JF, Van Loon LC. Systemic resistance in Arabidopsis induced by rhizobacteria requires ethylene-dependent signaling at the site of application. Mol Plant Microbe Int. 1999;12(8):720-7.
41. Ruci?iska-Sobkowiak R. Oxidative stress in plants exposed to heavy metals. Postepy Biochem. 2010;56(2):191-200.
42. Jomová K, Morovic M. Effect of heavy metal treatment on molecular changes in root tips of Lupinus luteus L. Czech J Food Sci. 2009;27(Special issue):386-9.
43. Liu T, Liu S, Guan H, Ma L, Chen Zh, Gu H, et al. Transcriptional profiling of Arabidopsis seedlings in response to heavy metal lead (Pb). Environ Exp Bot. 2009;67:377-86.
44. Abu-Romman S, Shatnawi M. Isolation and expression analysis of chloroplastic copper/zinc superoxide dismutase gene in barley. S Afr J Bot. 2010;77(2):328-34.
45. França SC, Roberto PG, Marins MA, Puga RD, Rodriguez A, Pereira JO. Biosynthesis of secondary metabolites in sugarcane. Genet Mol Biol. 2001;24(1-4):243-50.
46. Hradilová J, Rehulka P, Rehulková H, Vrbová M, Griga M, Brzobohatý B. Comparative analysis of proteomic changes in contrasting flax cultivars upon cadmium exposure. Electrophoresis. 2010;31(2):421-31.
47. Ammar WB, Nouairi I, Zarrouk M, Ghorbel MH, Jemal F. Antioxidative response to cadmium in roots and leaves of tomato plants. Biol Plantarum. 2008;52(4):727-31.
48. Alsher RG, Erturk N, Heath LS. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J Exp Bot. 2002;53(372):1331-41.
49. Wang C, Wang X, Tian Y, Xue Y, Xu X, Sui Y, et al. Oxidative stress and potential biomarkers in tomato seedlings subjected to soil lead contamination. Ecotoxicol Environ Saf. 2008;71(3):685-91.
50. Malecka A, Jarmuszkiewicz W, Tomaszewska B. Antioxidative defense to lead stress in subcellular compartments of pea root cells. Acta Biochim Polon. 2001;48(3):687-98.
51. Sanchez-Fernandez R, Fricker M, Corben LB, White NS, Sheard N, Leaver CJ, et al. Cell proliferation and hair tip growth in the Arabidopsis root are under mechanistically different forms of redox control. Proc Natl Acad Sci USA. 1997;94(6):2745-50.
52. Verma Sh, Dubey R. Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Sci. 2003;164(4):645-55.
53. Pitzschke A, Fornazi C, Hirt H. Reactive oxygen species signalling in plants. Antioxid Redox Signal. 2006;8(9-10):1757-64.
Received in March, 2013.
Accepted in May, 2013.
Sandra Pérez. Departamento de Biología y Sanidad Vegetal, Universidad Agraria de La Habana Fructuoso Rodríguez Pérez. Carretera Tapaste, Km 22 ½, San José de las Lajas, Mayabeque, Cuba. E-mail: sandra05@isch.edu.cu.


{kind=link}
{kind=link}
{kind=link}