










My SciELO
 Custom services
Custom servicesServices on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBiotecnología Aplicada
On-line version ISSN 1027-2852
Biotecnol Apl vol.31 no.3 La Habana July.-Sept. 2014
REVIEW
Evolutionary, physiological and biotechnological aspects of ferrochelatase and heme in higher plants
Aspectos evolutivos, fisiológicos y biotecnológicos de la ferroquelatasa y hemo en plantas superiores
Patricia Ortega-Rodés1, Bernhard Grimm2, Eduardo Ortega1
1 Laboratorio de Fisiología Vegetal, Departamento de Biología Vegetal, Facultad de Biología, Universidad de La Habana, La Habana, Cuba.
2 Institute of Biology/Plant Physiology, Humboldt University, Philippstr.13, Building 12, 10115 Berlin, Germany.
ABSTRACT
Heme, is a cofactor for essential proteins, it is synthesized after the insertion of Fe2+ into protoporphyrin ring by the ferrochelatase (FC) enzyme. Although FC is well described, there are still major questions about the regulation of the heme biosynthetic pathway, and the localization of the enzyme in plants. This review provides a comprehensive survey of research on function and the metabolism of heme in plants including the unique properties of the plant heme metabolism. We will summarize the knowledge of the expression, reaction mechanism, localization and the phylogeny of FC. Increasing the heme iron content in plant foods is a biotechnological challenge to improve the dietary intake of iron in the population; it is known that heme-contained iron is absorbed 5-10 times higher than non-heme iron. In this review we suggest potential biotechnological modifications of plant heme metabolism for increasing the capacity of crop plants to resist abiotic and biotic stress, thus improving food and fodder plants as source of dietary iron
Keywords: tetrapyrrole, iron, evolution, biotechnological plant improving.
RESUMEN
El grupo hemo, cofactor para proteínas esenciales, es sintetizado a partir de la inserción de Fe2+ en el anillo de protoporfirina por la enzima ferroquelatasa (FC). A pesar de que FC está bien caracterizada, existen aún incógnitas principales acerca de la regulación de la vía biosintética y la localización de la enzima en las plantas. Este artículo provee una revisión detallada acerca de las investigaciones sobre la función y el metabolismo del hemo en las plantas; incluye además información sobre las características únicas del metabolismo del hemo. Resumimos los conocimientos sobre la expresión, mecanismos de reacción, localización y filogenia de la FC. Incrementar el contenido de hierro hemo en los alimentos de origen vegetal es un gran reto biotecnológico para mejorar la ingesta de hierro en la población; el hierro hemo es de 5-10 veces más absorbido que el hierro no hemo. En esta revisión nosotros sugerimos modificaciones biotecnológicas potenciales en el metabolismo del hemo en las plantas para incrementar la capacidad de los cultivos de resistir los estreses biótico y abiótico y mejorar las cualidades nutricionales de las plantas como fuente de hierro para la dieta.
Palabras clave: tetrapirroles, hierro, evolución, mejoramiento biotecnológico de plantas.
INTRODUCTION
During the past decades, a significant increase of reports became evident on studies of successful attempts to apply genetic engineering to multiple plant species, variants and cultivars which highlight the potential to improve their yield, herbicide resistance and stress resistance among others characteristics [1-5].
Transgenic plants, in which the tetrapyrrole biosynthetic pathway was genetically modified are potentially useful for agricultural and horticultural applications [6]. None of these studies with transformants included modified ferrochelatase (FC) expression or attempts to improve heme production.
Here we outline a few principles for the use of transgenic plants with modified heme biosynthesis. Two aspects can be considered for this biotechnological approach to modify plant heme synthesis: 1) plants as heme iron source in food and fodder production 2) heme and its role in stress defense.
Heme is an essential molecule for all living organisms; it is the prosthetic group of several apoproteins, including hemoglobin, cytochromes, which are involved in the electron transport chains of photosynthesis and respiration, peroxidases, catalases, nitrite reductase and nitric oxide synthase and many more.
Heme iron from animal food is relatively soluble and is absorbed by specific receptors. Iron is then released by the action of heme oxygenase (HO) and enters the labile cytosolic iron pool as ferrous iron (Fe2+). Non-heme iron is more heterogeneous and is often present as poorly soluble ferric iron (Fe3+) or as iron tightly bound to chelators [7]. In human, the percentage of absorbed heme-contained iron is 5-10-fold higher than non-heme iron. Although heme represents only 10-15% of dietary iron in meat-eating populations, it may account for nearly one-third of absorbed iron [8].
Heme is synthesized in a universal way and those organisms, which are not able to synthesize heme de novo, depend on heme supply, by dietary intake. Iron deficiency causes an enormous problem in world’s population, with approximately two billion people suffering from anemia [9]. Plant food (especially staples like rice, maize, wheat and beans) tend to be poor sources of dietary iron. Thus, significant interest exists to generate crop varieties with elevated levels of bioavailable iron [10].
To date, transgenic rice lines transformed with the soybean ferritin gene had higher iron and zinc content in the grain even after polishing the seeds [11]. However, increasing heme iron in the total iron content in staple food remains an important challenge.
The importance of engineered food and fodder plants with high heme content is not only because of the need for higher iron content for nutrition, but also because heme content can influence the yield and stress tolerance of crops, as we will discuss.
REGULATION OF HEME SYNTHESIS
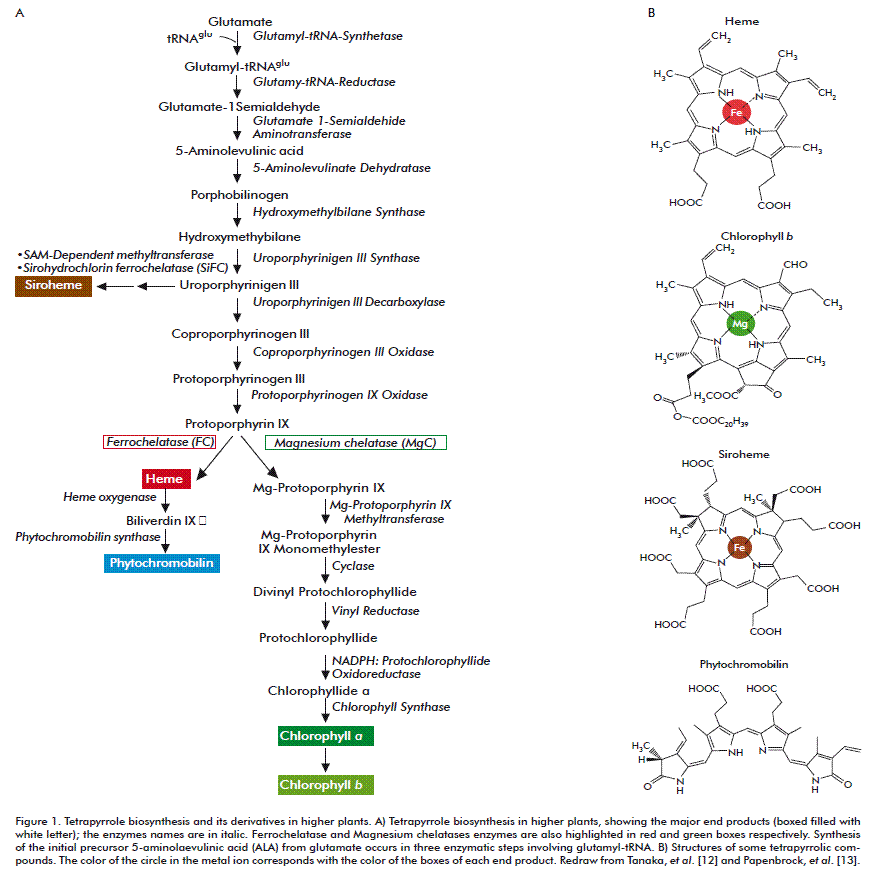
The metabolic pathway of the synthesis of chlorophyll and heme in plants starts from glutamate to form 5-aminolevulinic acid (ALA) (C5 pathway) (Figure 1). Eight molecules of ALA are assembled to form the tetrapyrrole skeleton. At first, the linear tetrapyrrole molecule, hydroxymethylbilane is formed, followed by an isomerization that produces the first cyclic tetrapyrrole, uroporphyrinogen III. Uroporphyrinogen III can continue along one branch to produce siroheme or towards the porphyrin branch in which the molecule is decarboxylated and oxidized to form protoporphyrin IX (ProtoIX). In continuation from ProtoIX, the pathway has two branches, one for the synthesis of chlorophyll and another for the synthesis of heme [12, 13].
The insertion of divalent metal ions into a range of modified tetrapyrroles is catalyzed by enzymes of the chelatase type. Magnesium chelatase (MgC; EC 6.6.1.1) inserts Mg2+ into ProtoIX generating Mg-ProtoIX as a precursor for chlorophyll synthesis; ferrochelatase (FC; EC 4.99.1.1) and sirohydrochlorin ferrochelatase (SiFC; EC 4.99.1.4) insert Fe2+ into ProtoIX and uroporphyrinogen III generating either protoheme (heme b) or, siroheme; cobaltochelatase (CoC; EC 6.6.1.2) insert Co2+ into hydrogenobyrinic acid a,c-diamide leading with some additional reactions to cobalamin.
The distribution of ProtoIX for either heme or chlorophyll synthesis requires a tight pathway control to direct the substrate for FC or MgC into the iron or magnesium branch of tetrapyrrole biosynthesis [14]. FC exists as a monomer or homodimer without cofactor requirement. MgC consists of three different subunits and requires ATP as substrate and for activation. ATP inhibits FC activity [15]. MgC has a Km for deuteroporphyrin of 8 nM, which is three hundred times lower than that of FC (2.4 µM); however, it is not excluded that MgC may compete with FC for the ProtoIX substrate [16, 17].
In plants, the requirement of heme and chlorophyll depends among others on cell type, stage of development, abiotic stress and light-dark cycle. A continuous supply of heme is needed in all cells to supply heme for respiratory cytochromes and other essential heme-proteins. Papenbrock, et al. [18] found in tobacco leaves about 40 nmol heme gFW-1 compared with 1150 nmol chlorophyll g FW-1. This indicates a strong regulatory demand in photosynthetic cells to direct ProtoIX to MgC. Non-photosynthetic tissue such as roots exclusively synthesizes heme; chlorophyll formation ultimately depends on the light in angiosperms [19].
The activity of FC oscillates during a 24 h day-night cycle in nearly a reverse amplitude than the MgC activity [13]. This diurnal oscillation could be a consequence of different supplies of ATP for the activity of each enzyme. In the light, when ATP levels are high in chloroplasts, MgC activity is enhanced and FC activity might be limited.
Another regulator of the tetrapyrrole pathway is GUN4, which interacts with Mg-chelatase and stimulates its activity by facilitating substrate binding and/or product release and possibly the interaction of MgC with chloroplast membranes [20, 21].
It is hypothesized that pathway control of chlorophyll or heme synthesis in plants includes not only the temporal regulation but also a spatial separation of the two enzymes FC and MgC. If the enzymes are located in different subcompartments of chloroplasts, they would not directly compete for the same pool of substrate. There is contradictory evidence concerning the localization of these two enzymes in different parts of chloroplasts. According to Matringe, et al. [16] the activity of FC was associated only with thylakoid membranes of intact pea chloroplasts and not detected in highly purified envelope membrane fractions. However, Roper and Smith [22], localized Arabidopsis FC in the envelope, stroma and thylakoids from peas (Pisum sativum L.). Suzuki, et al. [23] did not detect FC in the stroma, but they showed a precursor FC import to both thylakoid and envelope membranes using isolated pea chloroplast. The localization of the subunits of Mg-chelatase (ChlH, ChlD, ChlI, ChlM) is particularly difficult to indicate precisely. Spectral counting reveals that all four subunits are found in the stroma, but that ChlH is also found in the envelope, whereas ChlD is also present in thylakoids [24]. ChlH associates with the envelope membrane at high (5 mM) Mg2+ concentration, whereas it is detected in the stroma at lower (1 mM) Mg2+ concentration [25]. All these results indicate that localization of both enzymes is still not entirely elucidated and methodology for spatial characterization has to be improved. If both enzymes can be detected in the same compartment, the proposed idea about spatial separation of FC and MgC is obsolete. Then, other mechanisms are necessary to control substrate channeling in the tetrapyrrole biosynthesis pathway.
A possible explanation for the diverse ProtoIX accumulation in the FC and MgC antisense lines could be the complex channeling mechanism of ProtoIX to the two chelatases. FC antisense lines with lower FC activity accumulate ProtoIX [26], but the MgC antisense lines [18] did not accumulate ProtoIX. Once the substrate is dedicated to heme synthesis, it seems to be inaccessible for the MgC branch, even if both enzymes are localized in the same organelle and in the same fraction.
Up to now we and other research groups have analyzed the regulatory aspects of heme and chlorophyll synthesis in the chloroplast only. However, in contrast to chlorophyll confined to chloroplast, heme is widely distributed in all cellular compartments. Therefore, it is challenging to clarify, how heme is shuffled into these subcellular compartments and whether heme synthesis takes place also in plant mitochondria.
SUBCELLULAR LOCALIZATION OF FERROCHELATASE IN PLANT TISSUES
Ferrochelatase is a nuclear-encoded protein and is translocated to the subcellular site where it accomplishes its functions. Proteins, like ferrochelatase, that are localized in organelles require a transit peptide, presequence, signal peptide or any recognition motif to be precisely targeted to the designated cellular compartment [27, 28].
Most reports on plant heme synthesis address the plastid-localized pathway. Only a few publications suggest that the last two steps of heme synthesis occur parallel in plant plastids and mitochondria [29-31]. To date it is still not entirely proven whether plant mitochondria have their own heme synthesis. The localization of the two isoforms (FC I and FC II, only described for plants) of ferrochelatase is an open question because of contradictory findings. All authors agree on the localization of FC I and FC II within the plastids, but debate about the presence of FC I in mitochondria.
Import experiments of Arabidopsis FCs into isolated pea chloroplasts and mitochondria showed that the FC II gene encodes a precursor, which is imported solely into chloroplast but not into mitochondria. In contrast, FC I could be targeted to chloroplast and mitochondria in organelle uptake experiments [31]. Similar results were obtained with Cucumis sativus FCs using an in vitro import technique of isolated pea organelles and it was demonstrated that FC II (CsFeC2) was predominantly localized in thylakoid membranes as an intrinsic protein, but a minor portion was also detected in envelope membranes; FC I (CsFeC1) was imported into mitochondria and processed to a protein essentially identical to that imported by chloroplasts [23]. In contrast, Lister, et al. [32] found that none of Arabidopsis thaliana ferrochelatase isoforms were imported into Arabidopsis mitochondria.
Apparently, the body of evidence is more unequivocal for protoporphyrinogen oxidase [33], the preceding enzyme of FC. There are clear evidence for the translocation and activity of PPO in plastids as well as in mitochondria [34]. PPO catalyzes the oxidation from protoporphyrinogen IX to ProtoIX and was reported to exist in tobacco in the isoforms I and II, which are located in chloroplasts and mitochondria, respectively [35]. Spinacia oleracea PPO II has been reported to be dual-targeted into mitochondria and plastids [36] and perform the oxidation reaction of protoporphyrinogen IX in both organelles. In plastid, the ProtoIX is channeled into MgC or FC branch. The successive use of ProtoIX produced by PPO II by FC in mitochondria is still in question due to the non-conclusive data about the presence of FC inside this organelle. The potential ability of FC to perform dual targeting would allow completing the two-step heme synthesis in mitochondria. Alternatively, an additional function of PPOII in mitochondria has to be taken into account.
Many efforts have been undertaken to prove detection of FC inside mitochondria but the experimental evidences of localization remained debatable. Using transgenic lines expressing fluorescent-tagged FC I Woodson, et al. [37] did not show experiments that FC I is localized in mitochondria, but FC I was exclusively imported into plastids. Other authors have also failed with their approaches to show mitochondrial localization of FC I when plants were transformed with genes encoding chimeric FC I-GFP. A reason for the outcome of these experiments could possibly be a different expression level of FC inside plastid and mitochondria. Singh, et al. [38] found about 110 fold more total ferrochelatase activity in chloroplast than in the mitochondria of pea plants. It must also be noted that a bona fide evidence for mitochondria-targeted FC I requires the isolation of pure and intact mitochondria and a specific anti-FC-antibody. Only isolated mitochondria could enable measurements of the low FC activity in mitochondria. Moreover, any chloroplast contamination has to be excluded. It is proposed that FC I overexpressing lines could contribute to the elucidation of these challenging questions, when an excessive amount of FC may allow a prediction which compartment benefits from the increasing heme-synthesizing capacity. In addition, experiments with FCs isoforms from Oriza sativa plants show that the FC II has a typical transit sequence (70 amino acids) for plastid targeting, but isoform I has no apparent transit sequence [39].
Emanuelsson, et al. [40] developed the TargetP software that is able to discriminate between proteins which are destined for the translocation into the mitochondrion, the chloroplast, the secretory pathway, and other localizations using N-terminal sequence information with a success rate of 85 % for plants. Table 1 shows the scores for the mitochondrial/plastidal localization of some ferrochelatase proteins, especially calculated for this review, using different available software: TargetP [40]; MitoProtII [41]; iPSORT [42]; TargetLoc [43].
We use as positive controls for mitochondrial prediction FC proteins of human (Homo sapiens), yeast (Saccharomyces cerevisiae) and red algae (Cyanidioschyzon merolae) that have been previous demonstrated to be localized in mitochondria [44-46]. Using TargetP and TargetLoc, the translocalization of the FC of some plants (Arabidopsis sp., Nicotiana tabacum, Oryza sativa, Hordeum vulgaris, Cucumis sativus, Zea mays and Solanum tuberosum) was predicted to be mainly targeted to chloroplasts, but to lesser extent to mitochondrion or secretory pathway, due to the fact that the sequences contain a predicted chloroplast transit peptide. The very low values obtained for mitochondrial targeting mean a low probability that the protein is targeted into mitochondria. In the case of HvFC1, OsFc1 and ZmFC1the values were similar for both organelles using TargetP; however using TargetLoc, mitochondria localization obtained a higher score.
For some proteins, MitoProt II and iPSORT confirmed the prediction of chloroplast targeting. Using MitoProt II, the isoform FC2 of Arabidopsis lyrata (AlFC2) and Hordeum vulgaris (HvFC2) was predicted to be targeted to mitochondria with high probability values (0.9203 and 0.7025 respectively). Using the iPSORT software, NtFc1, OsFc1, HvFC2 and OsFc2 were predicted to be target to mitochondria. These results convincingly predict mitochondria localization for some FC proteins that need to be demonstrated with other experimental methods. The results obtained and shown in table 1 indicate that among higher plants, Hordeum vulgaris, Oryza sativa, and Zea mays seem to be promising species for experimental proofs of ferrochelatase translocation to both organelles.
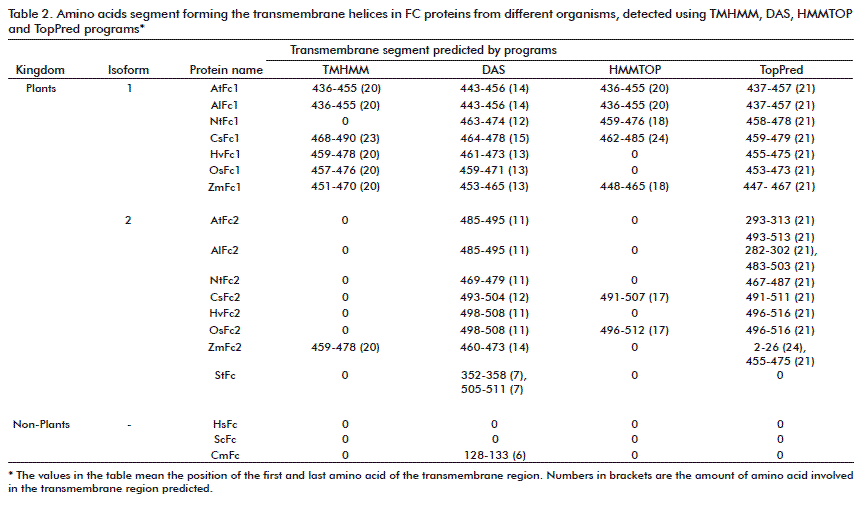
Mitochondrial outer membrane proteins are synthesized without a cleavable presequence, but contain a peptide motif responsible for mitochondrial targeting and membrane integration within the molecule: a transmembrane segment and N- or C-terminal flanking segments [47]. Except for the B. subtilis enzyme, which is a soluble protein [48], ferrochelatase has been reported to be a membrane-associated protein in chloroplasts (located in the inner envelope membrane and the thylakoid membrane) and in mitochondria (inner membrane) of Eukaryotes, or to be associated with cytoplasmic membrane in prokaryotes [46, 49]. The use of different software programs available in the ExPAsy Web (TMHMM [50], DAS [51], HMMTOP [52], TopPred [53]) reveals in some cases the presence of transmembrane domains (TMD) in C-terminal parts of the protein with maximum lengths of 24 amino acids (Table 2). The TMD is predicted for all 15 plant FCs tested when using the DAS and TopPred software; however when using the TMHMM and HMMTOP software programs, the TMD appears more commonly in the FC I isoform and only rarely in the FC II isoform.
FC II isoform from plants as well as cyanobacterial and algal FC possess a carboxyl-terminal Chl a/b-binding (CAB) domain which forms a putative transmembrane segment containing a conserved Chl-binding motif [23, 54]. However, FC enzyme truncated lacking the C-terminal domain in Synechocystis mutant is located almost exclusively in the membrane fraction [55].
Taken all together we can assume that the plant FC is not an integral membrane protein, but it is assumed to be associated with membranes. This is in agreement with previous findings, when FC has been purified from membrane fractions and its activity was enhanced with fatty acids [46]. The association of FC with the membrane could be realized through its own transmembrane motif (as it was predicted for some FC proteins using different software programs) or by non-ionic interaction between FC and the membrane or by interaction with other membrane proteins. In bovine heart mitochondria, FC is associated with Complex I, indicating that ferrous ion is produced by NADH oxidation in Complex I and is then utilized for heme synthesis by FC [56]. The association of the enzyme with the membrane might be a requirement for heme synthesis due to the low solubility of the substrate (ProtoIX) and the product of the reaction (heme) in aqueous solutions at neutral pH [57].
The localization of PPO in plant mitochondria (as well as in plastids) [35, 36] and a possible association with a putative plant mitochondrial FC would facilitate substrate (ProtoIX) channeling. Thus, the accumulation of ProtoIX, potentially toxic, is avoided and the production of heme for the mitochondrial heme proteins like respiratory cytochromes occurs inside the mitochondria. Nevertheless, up to now, we are aware on the basis of current data that we only speculate about the fate of mitochondrial heme synthesis; more efforts should be made to unravel the subcellular localization of heme synthesis in plants.
On the other hand if there is no FC in the mitochondria, a heme export from chloroplast and import into mitochondria is necessary to ensure the assembly of heme-binding proteins assembly. Studies with developing chloroplasts from greening cucumber cotyledons have shown that these chloroplasts are capable of heme efflux with a steady-state rate between 0.12 and 0.45 pmol heme∙minute-1mg plastid proteins-1 [58]. Direct evidence of heme import into plant mitochondria is still awaited; the relevant studies have not been published.
HEME SIGNALING IN PLANTS
Because the majority of proteins found in mitochondria and plastid are encoded by nuclear genes, transcribed in the nucleus, translated in the cytoplasm, and then imported into mitochondria or plastid [59, 60] the signaling between mitochondria/plastids and the nucleus is required to maintain organellar biological functions [61, 62].
Signaling between organelles and the nucleus is bidirectional by an anterograde and retrograde control. A number of mechanisms have been evolved to provide a direct control of organellar gene expression by nuclear genes [63]. Anterograde signaling originates in the nucleus and can be induced by environmental (e.g. light) and developmental cues. In contrast, retrograde signaling originates within the organelles and feeds back to alter the expression of nuclear genes that encode organelle-localized proteins [14].
Mense and Zhang [64], Nott, et al. [65] have suggested that tetrapyrrole molecules, such as Mg porphyrins and heme mediate plastid-to-nucleus or mitochondria-to-nucleus retrograde signaling to coordinate nuclear gene expression in plants and animals, respectively. Tetrapyrrole signaling could be a mechanism for coordinating the cell cycles as well as fine-tuning the transcriptional control in various processes [66]. However, some other results argue against the function of tetrapyrrole as signaling molecule, especially Mg-ProtoIX and other chlorophyll biosynthetic intermediates [67, 68].
Some signal molecules are necessary to control photosynthesis-associated nuclear gene (PhANGs) expression. Transgenic Arabidopsis plants over-expressing plastid FC I show increased PhANGs expression. A specific heme pool produced by FC I was proposed to be involved in plastid-to-nucleus communication when heme is being exported from healthy chloroplasts [37].
As we have seen earlier, heme is part of many important molecules for life; its role as signal and its participation in regulatory mechanisms has an outstanding relevance.
As a product of the mitochondria, heme is ideally suited to coordinate the expression of mitochondrial and nuclear genes involved in the biogenesis of the mitochondrial respiratory complexes [69]. However up to now there are not conclusive evidences about the mitochondrial origin of heme found in plants.
HEME SYNTHESIS IN TRANSGENIC PLANTS
As sessile organisms, plants are generally always affected by external environmental signals and have to react on these changes by modified gene expression to induce protective mechanisms. For the application of gene transformation to modify the response of plants to stresses it is important to identify the “useful genes” responsible for better stress tolerance [70].
Plants possess energy-associated central network responsible for the survival under stress. This network is more complex in plants than in animals, using primarily photosynthesis as the energy donor under optimal growth conditions, or glycolysis, the tricarboxylic acid (TCA) cycle, and amino acid catabolism upon exposure to stress [71]. Genes encoding most of the enzymes in chlorophyll and heme biosynthesis appeared to be grouped in the same cluster, indicating that a tightly coordinated stress-induced regulation of these multiple genes is required for efficient reduction in the levels of chlorophyll and heme upon exposure to stress. Such a mechanism may apparently protect plants from the accumulation of toxic reactive oxygen species derived from unused tetrapyrroles. Because of the chemical properties of tetrapyrroles as singlet-oxygen generators, it is tempting to speculate that plants use tetrapyrrole molecules for the singlet-oxygen generation, which may contribute to ROS-triggered defense or protective response.
It is reasonable that in response to stress the heme content will be increased by de novo heme synthesis for the completion of cytochrome c inside mitochondria or the additional demand for heme-containing peroxidases. Pasqualini, et al. [72] demonstrated that during the programmed cell death induced by ozone, an early decrease of cytochrome c content occurred by its release from the mitochondria. Additionally, the induction of FC I gene and the increase in FC activity under different stress conditions occur, the presumably by supplying heme for heme-proteins involved in the defense response [38, 73].
It is proposed that plants with modified capacity to synthesize more heme to avoid the feedback control on ALA synthesis capacity would be able to respond to stress. This type of plants would be a useful tool to exploit growth under adverse environment limiting plant crops growths.
To obtain a transgenic plant with elevated production of heme, it might be necessary to modify more than one enzymatic step. For heme synthesis, FC needs ProtoIX and Fe2+. The ProtoIX synthesis was discussed previously. The supply of Fe2+ to FC depends on different factors: the plant iron supply and metabolism; the redox state in the cells; the activity of ferric reductase oxidase enzyme (FRO), the iron transport through the organelle membranes and the iron sequestration by chaperone molecules.
Iron availability is often low in soils. Non-grass plants have adopted to the so-called strategy I, in which Fe(II) transport is coupled to a Fe(III)-chelate reduction step. Grasses have developed strategy II with phytosiderophores (PS) as relevant components, chelating Fe(III) [74]. Fe(III)-PS complexes are then taken up into the root by a specific transporter. In maize, a transporter-Fe(III)-PS complex is encoded by the Yellow Stripe 1 (YS1) gene [75]. Arabidopsis Yellow Stripe 1-Like (AtYSL1) provides iron during seed filling in Arabidopsis [76]. It could be an important step to modify plants used to produce food seeds; the grains could be heme-iron rich.
Distribution of Fe to the various plant organs involves long-distance transport through the sap [76]. Once iron has entered the plant, both nicotianamine and citrate serve as iron chelators. Increasing the expression of nicotianamine synthase (NAS) increased the level of bioavailable iron (3.0-fold) in mature rice seeds and enabled mice after being fed with the transgenic rice seeds to recover rapidly from anemia [77].
There is very little free Fe present in the cytosol [78]. It is usually assumed that Fe may pass freely across the outer membrane of chloroplast and mitochondria via porins. Permeases in chloroplasts (PIC1) were proposed to transport iron into the organelle [79]. It is unknown whether Fe(II) or Fe(III) is transported by PIC1 and whether a Fe(III) chelate reductase is required.
Studies in Arabidopsis describe a member of the ferric reductase oxidase (FRO) which reduces Fe3+ to Fe2+; FROs may function at the organellar membranes. FRO7 plays an essential role in iron delivery to chloroplasts [80]; FRO3 and FRO8 localized in the mitochondria [81] contribute to mitochondrial iron homeostasis.
Ferrous iron is then translocated across the inner membrane by plastid or mitochondria iron transporter. Experiments using vesicles with isolated inner envelopes from pea chloroplast indicate that Fe2+ transport can take place bidirectionally by the same transporter [82]. Mitochondrial iron transporters are conserved small proteins that belong to the mitochondrial carrier family localized in the mitochondrial inner membrane and are involved in solute transport into the mitochondrial matrix [10]. The first mitochondria iron transporter (MIT) in plants was described in rice by Bashir, et al. [83]. Rice mit knockdown mutants exhibit a low growth phenotype, reduced chlorophyll concentration, poor seed yield and reduced mitochondrial iron concentration. Total iron concentration in this mit knockdown plants is elevated, indicating that iron is apparently mis-localized [83]. The excess of cytosolic iron may be directed toward vacuoles by the vacuolar iron transporter 1 (VIT1). The MIT plays an important role in seed development and its expression level is positively regulated by iron availability [10].
Once the Fe2+ is inside the mitochondrial matrix, it is received by an iron chaperone, frataxin (FH). FH distributes this Fe to the Fe-S cluster assembly proteins and, possibly, to the heme biosynthetic machinery [10] if the second heme synthesis inside the mitochondria is also considered.
A. thaliana frataxin-deficient lines (AtFH) exhibit a decrease in total heme content in leaves and flowers and alterations in several transcripts from the enzymes involved in heme biosynthesis; the genetically modified AtFC1 and AtFC2 expression increased about 1.5-2 fold in leaves. Moreover these mutant plants also suffer from a deficiency of catalase activity, indicating that AtFH, apart from its role in protecting bioavailable iron within mitochondria and the assembly of Fe-S groups, also plays a role in the production of heme groups and the activity of heme-proteins in plants [84].
Because iron reactivity with organic ligands is very high, a good control for Fe content that includes accounting for movement and compartmentalization inside the cell is necessary. The ferritin protein stores iron in a non-toxic form and releases it when needed for metabolic functions [85]; thus it serves to prevent oxidative damage [86, 87]. Ferritins exhibit ferroxidase activity oxidizing Fe2+ and can accommodate up to 4500 Fe3+ atoms per ferritin molecule [88]. The molecular mechanism underlying the release of iron from ferritins in plants is not described. Plant ferritins are localized mainly in plastids but they can also be targeted to mitochondria [89].
Some authors have found increased Fe content when the expression of ferritin is increased in rice endosperm [11, 90]. In Arabidopsis it has been demonstrated that ferritins do not constitute the major seed iron pool, and that the absence of ferritins does not have an impact on germination or on proper development of the young plant. Loss of ferritins in vegetative and reproductive organs does result in sensitivity to excess of iron, as shown by reduced growth and large defects in flower development [86].
The ferritin content increases in Lemna minor growing in water highly contaminated with iron [91]. This small floating monocotyledoneous macrophyte is able to grow and remove iron from iron rich neutral mine drainage and has a potential use in iron phytoremediation. The high rate of iron removal, suggests that the species could be particularly interesting for the decontamination of mine discharges that are continuously produced, contaminating streams and the rivers [92]. Modified plants with high capacity to produce ferritin, could improve the Fe storage in vacuoles and thus avoid cytotoxicity making the Fe unavailable, thus preventing oxidative stress, while allowing normal development and plant growth. Plants which had been transformed with the high capacity to remove iron from contaminating sources would have a potential field of application.
Other genes, encoding either iron transporter or iron translocation proteins are important for iron homeostasis. Over-expression of rice Fe transporters OsIRT1 resulted in a slight rise in Fe concentration in seeds [93]. Rice Fe-nicotianamine transporter (OsYSL2) is important for Fe translocation, especially in the shoots and endosperm. When OsYSL2 expression was driven by the sucrose transporter promoter, the Fe concentration in the polished transformant was up to 4.4-fold higher compared to the polished wild type seeds [94].
Transgenic approaches, like manipulation of the ferritin and the iron transporter content as well as translocation of metals have been used to elevate Fe accumulations in cereal grains. Any of these biotechnological modifications deal with a heme catabolism and anabolism. A goal to improve plant iron content for animal and human feeding implies that plants enhance their iron absorption and transport and that this iron is ultimately inserted into the Proto IX ring, which is absorbed at higher percentages than non-heme iron, at least in mammals. This process certainly needs a finely fine-tuned control for which in turn is necessary to understand the tetrapyrrole pathway in plants very well.
HEME AND FERROCHELATASE IN THE EVOLUTIONARY PROCESS
In animals and fungi, heme synthesis is separated in enzymatic steps located in the cytosol and in mitochondria, but in plants more recent papers highlight that heme synthesis most likely takes place only in chloroplasts. A potential mitochondrial heme synthesis is still under discussion. The idea of a possible mitochondria localization of plant FC is analyzed here taking into account the evolutionary origin of plastids and mitochondria and the structural similarity of FC proteins of photosynthetic and non-photosynthetic organisms.
The endosymbiotic theory suggests the derivation of plastids and mitochondria of eukaryotic cells from bacterial endosymbionts [95]. Mitochondria and plastids are proposed to derive from descendants of α-proteobacterium- and cyanobacterium-like progenitors, respectively. The heme biosynthesis in photosynthetic eukaryotes depends solely upon nucleus-encoded proteins of mixed origin (cyanobacterial and non-cyanobacterial) [63, 96]. This mix origin could explain a possible heme pathway in plant mitochondria derived from α-proteobacterium origin.
Some indications for the existence of multiple heme biosynthetic pathways can be deduced from analysis of the tetrapyrrole biosynthetic pathway in Euglena gracilis. In this organism, ALA is synthesized in two different routes: from glutamate like in higher plants, and via condensation of glycine and succinyl-CoA, as it is found in animals. Euglena evolved from an organism which is derived from a secondary endosymbiotic origin. Weeden [97] recognized that the endosymbiont introduced novel pathways (for amino acid, heme, and starch syntheses) into the host via endosymbiosis.
Engulfment of a cyanobacterial ancestor and the subsequent restructuring to semiautonomous organelles within the eukaryotic host initially led to the formation of four lineages with primary plastids: the chlorophytes, embryophytes, rhodophytes and glaucophytes [98]. After primary symbiosis, the common ancestor of plants may have retained FC in both mitochondria and plastids, at least during the first phase. However, according to recent findings, in plants, FC has only been found in plastids, but not in mitochondria. In contrast, in the unicellular red algae Cyanidioschyzon merolae, FC has been detected only in the mitochondria and not in plastids. Plastids in red algae and in green plants differ significantly from each other (e.g. in structure and in the light harvesting mechanism) and allow reliably to distinguish between the red and green plastid lineage [99]. It is suggested that the different properties of these plastids would influence the capacity to recognize the FC precursor.
During evolution, the FC gene from the progenitors of the two organelles might be lost, either the mitochondrial FC from green plants or the plastid FC from red algae [45]. But not all descendents may have had the same fate. So, it might be still sensible to continuously examine FC activity in plant mitochondria.
Table 3 shows all divisions of organisms belonging to the Viridiplantae harboring FC sequences, which are published in NCBI data base. The classification was performed according to Judd, et al. [100]. FC accessions have not been published in NCBI data base for all genera, classes or divisions, yet.
To date, one hundred and thirty five FC protein sequences of Viridiplantae have been registered on NCBI. Only a few of them have been published and characterized. Seed plants are the group with the highest number (102) of sequences reported in 24 genera, followed by green algae with 17 sequences in 8 genera, and Bryophytes and Lycopodiophyta with 8 sequences each. Among the seed plants, the Zea and Oryza genera have the highest number of reported sequences with 25 and 17, respectively, but some of them are identical.
Especially for this review, a phylogenetic tree of FC proteins was constructed using amino acid sequences from photosynthetic Eukaryotes, cyanobacteria, α- and γ-proteobacteria, fungi, animals, apicomplexa and Archaea. Among the complete FC sequences presented in NCBI for Eukaryotic photosynthetic organism, we used 23 representatives of red algae, green algae and land plants for preparing the phylogenetic tree. The sequences were downloaded from GenBank [101]; accession numbers are given in the figure caption (Figure 2). Sequences were aligned using the ClustalX program [102]. Phylogenetic and molecular evolutionary analyses were conducted with the help of the MEGA software version 5 [103]. The phylogenetic tree was constructed for this review using the maximum likelihood (ML) method, WAG model and discrete Gamma (G) distribution in five rate categories assuming that certain peptide motifs and amino acid residues are evolutionarily invariable (I).
Only one isoform of FC has been reported in animals, fungi, bacteria and algae, but some plants have two FC isoforms (I and II). An open question regarding the evolution of heme synthesis is how plants evolved to possess two isoforms of FC. Here we address and analyze two possible evolutionary reasons for the existence of two FC isoforms in plants: 1) Both isoforms were acquired from the different organelles (plastids or mitochondria dependent from the bacterial endosymbiont) during the endosymbiotic event or 2) as a result of the gene duplication of the FC gene acquired from the cyanobacterial ancestor.
Among the photosynthetic organisms it is possible to distinguish in the phylogenetic tree, a common ancestor of FCs in cyanobacteria and the plant clade with a bootstrap value of 100. Both isoforms (I and II) of FC in plants are grouped separately from other organisms with high bootstrap value (98).
According to the phylogenetic tree, FC of non-photosynthetic organisms (No-Ph-FC) diverged from photosynthetic organisms (Ph-FC). This divergence could be related to a functional specialization of FC in photosynthetic and non-photosynthetic organisms. If the evolutionary origin of FC I were derived from mitochondria of the non-photosynthetic organism, FC I would appear closer to the groups of non-photosynthetic organism in the tree.
But, the plant FC I is phylogenetically more related to the plant FC II than to FC localized in mitochondria from non-photosynthetic organisms, indicating that probably both plant FC isoforms are derived from independent gene duplication. As was discussed before about the possible localization of FC I in mitochondria, it could be due to a secondary specialization associated to this organelle and is not explained by a gene derived from the mitochondrial ancestor. Thus, depicted from the phylogenetic tree, even if FC I is located in mitochondria, the FC I gene is descendent from the cyanobacterial ancestor and its gene product has an acquired function.
CONCLUDING REMARKS
Improving mineral nutrition through plant biotechnology may be a sustainable strategy to combat deficiencies in human populations [104]. Improving plant iron content became a major biotechnological challenge identified by the World Health Organization [105]. Enhancing the heme iron in fodder plants may be a good strategy for iron fortification. There are many potential biotechnological avenues to modify tetrapyrrole biosynthesis in photosynthetic organisms; these new transgenic plants could also contribute to elevated stress tolerance of plants or intracellular accumulation of dietary iron for human and animals.
Higher plants, sessile organisms, are highly affected by the environment and have few possibilities to escape from abiotic and biotic stresses. Plants instead have evolved a fine-tuned regulation of metabolism and a complex protection strategy. Two gene copies of FC with its differential gene expression seem to be part of this protective concept. As a consequence two pools of heme play different roles including the regulatory role as a signal molecule for communication between nucleus and organelles. We do not exclude the FC I as is also translocated to plant mitochondria, although the more recent reports highlight its localization in plastids. Three plant species (Hordeum vulgaris, Oryza sativa and Zea mays) are good candidates to search for FC I protein translocation to mitochondria.
ACKNOWLEDGEMENTS
Patricia Ortega Rodés and Eduardo Ortega Delgado express their gratitude to Jennifer Hosek, Mike Zirkel and Virginia Waddick for helping with the English language.
REFERENCES
1. De Greef W, Delon R, De Block M, Leemans J, Botterman J. Evaluation of herbicide resistance in transgenic crops under field conditions. Nature Biotechnol. 1989;7:61-4.
2. Reguera M, Peleg Z, Abdel-Tawab YM, Tumimbang EB, Delatorre CA, Blumwald E. Stress-induced cytokinin synthesis increases drought tolerance through the coordinated regulation of carbon and nitrogen assimilation in rice. Plant Physiol. 2013;163(4):1609-22.
3. Penna S. Building stress tolerance through over-producing trehalose in transgenic plants. Trends Plant Sci. 2003;8(8):355-7.
4. Garg AK, Kim JK, Owens TG, Ranwala AP, Choi YD, Kochian LV, et al. Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses. Proc Natl Acad Sci USA. 2002;99(25):15898-903.
5. Lermontova I, Grimm B. Overexpression of plastidic protoporphyrinogen IX oxidase leads to resistance to the diphenyl-ether herbicide acifluorfen. Plant Physiol. 2000;122(1):75-84.
6. Tanaka R, Tanaka A. Tetrapyrrole biosynthesis in higher plants. Annu Rev Plant Biol. 2007;58:321-46.
7. Murgia I, Arosio P, Tarantino D, Soave C. Biofortification for combating 'hidden hunger' for iron. Trends Plant Sci. 2012;17(1):47-55.
8. Cook JD. Adaptation in iron metabolism. Am J Clin Nutr. 1990;51:301-8.
9. Wu AC, Lesperance L, Bernstein H. Screening for iron deficiency. Pediatr Rev. 2002;23(5):171-8.
10. Jain A, Connolly EL. Mitochondrial iron transport and homeostasis in plants. Frontiers Plant Sci. 2013;4:348.
11. Vasconcelos M, Datta K, Oliva N, Khalekuzzaman M, Torrizo L, Krishnan S, et al. Enhanced iron and zinc accumulation in transgenic rice with the ferritin gene. Plant Sci. 2003;164(3):371-8.
12. Tanaka R, Kobayashi K, Masuda T. Tetrapyrrole metabolism in Arabidopsis thaliana. Arabidopsis Book. 2011;9:e0145.
13. Papenbrock J, Mock HP, Kruse E, Grimm B. Expression studies in tetrapyrrole biosynthesis: inverse maxima of magnesium chelatase and ferrochelatase activity during cyclic photoperiods. Planta. 1999;208(2):264-73.
14. Papenbrock J, Grimm B. Regulatory network of tetrapyrrole biosynthesis--studies of intracellular signalling involved in metabolic and developmental control of plastids. Planta. 2001;213(5):667-81.
15. Cornah JE, Roper JM, Pal Singh D, Smith AG. Measurement of ferrochelatase activity using a novel assay suggests that plastids are the major site of haem biosynthesis in both photosynthetic and non-photosynthetic cells of pea (Pisum sativum L.). Biochem J. 2002;362(Pt 2):423-32.
16. Matringe M, Camadro JM, Joyard J, Douce R. Localization of ferrochelatase activity within mature pea chloroplasts. J Biol Chem. 1994;269(21):15010-5.
17. Guo R, Luo ML, Weistein JD. Magnesium-chelatase from developing Pea leaves. Characterization of a Soluble Extract from Chloroplasts and Resolution into Three Required Protein Fractions. Plant Physiol. 1998;116(2):605-15.
18. Papenbrock J, Pfundel E, Mock HP, Grimm B. Decreased and increased expression of the subunit CHL I diminishes Mg chelatase activity and reduces chlorophyll synthesis in transgenic tobacco plants. Plant J. 2000;22(2):155-64.
19. Mohanty S, Grimm B, Tripathy BC. Light and dark modulation of chlorophyll biosynthetic genes in response to temperature. Planta. 2006;224(3):692-9.
20. Adhikari ND, Orler R, Chory J, Froehlich JE, Larkin RM. Porphyrins promote the association of GENOMES UNCOUPLED 4 and a Mg-chelatase subunit with chloroplast membranes. J Biol Chem. 2009;284(37):24783-96.
21. Larkin RM, Alonso JM, Ecker JR, Chory J. GUN4, a regulator of chlorophyll synthesis and intracellular signaling. Science. 2003;299(5608):902-6.
22. Roper JM, Smith AG. Molecular localisation of ferrochelatase in higher plant chloroplasts. Eur J Biochem. 1997;246(1):32-7.
23. Suzuki T, Masuda T, Singh DP, Tan FC, Tsuchiya T, Shimada H, et al. Two types of ferrochelatase in photosynthetic and nonphotosynthetic tissues of cucumber: their difference in phylogeny, gene expression, and localization. J Biol Chem. 2002;227:4731-7.
24. Joyard J, Ferro M, Masselon C, Seigneurin-Berny D, Salvi D, Garin J, et al. Chloroplast proteomics and the compartmentation of plastidial isoprenoid biosynthetic pathways. Mol Plant. 2009;2(6):1154-80.
25. Nakayama M, Masuda T, Bando T, Yamagata H, Ohta H, Takamiya K. Cloning and expression of the soybean chlH gene encoding a subunit of Mg-chelatase and localization of the Mg2+ concentration-dependent ChlH protein within the chloroplast. Plant Cell Physiol. 1998;39(3):275-84.
26. Papenbrock J, Mishra S, Mock HP, Kruse E, Schmidt EK, Petersmann A, et al. Impaired expression of the plastidic ferrochelatase by antisense RNA synthesis leads to a necrotic phenotype of transformed tobacco plants. Plant J. 2001;28(1):41-50.
27. Schatz G, Dobberstein B. Common principles of protein translocation across membranes. Science. 1996;271(5255):1519-26.
28. Rusch SL, Kendall DA. Protein transport via amino-terminal targeting sequences: common themes in diverse systems. Mol Membr Biol. 1995;12(4):295-307.
29. Smith AG, Santana MA, Wallace-Cook AD, Roper JM, Labbe-Bois R. Isolation of a cDNA encoding chloroplast ferrochelatase from Arabidopsis thaliana by functional complementation of a yeast mutant. J Biol Chem. 1994;269(18):13405-13.
30. Chow KS, Singh DP, Roper JM, Smith AG. A single precursor protein for ferrochelatase-I from Arabidopsis is imported in vitro into both chloroplasts and mitochondria. J Biol Chem. 1997;272(44):27565-71.
31. Chow KS, Singh DP, Walker AR, Smith AG. Two different genes encode ferrochelatase in Arabidopsis: mapping, expression and subcellular targeting of the precursor proteins. Plant J. 1998;15(4):531-41.
32. Lister R, Chew O, Rudhe C, Lee MN, Whelan J. Arabidopsis thaliana ferrochelatase-I and -II are not imported into Arabidopsis mitochondria. FEBS Lett. 2001;506(3):291-5.
33. Dimitrijevic L, Puppo A, Trinchant JC, Rigaud J. Ferrochelatase activities and heme contents in purified mitochondria from soybean roots and root nodules. Plant Sci. 1989;134(5):642-4.
34. Jacobs JM, Jacobs NJ. Oxidation of protoporphyrinogen to protoporphyrin, a step in chlorophyll and haem biosynthesis Purification and partial characterization of the enzyme from barley organelles. Biochemistry J. 1987;244(1):219-24.
35. Lermontova I, Kruse E, Mock HP, Grimm B. Cloning and characterization of a plastidal and a mitochondrial isoform of tobacco protoporphyrinogen IX oxidase. Proc Natl Acad Sci USA. 1997;94(16):8895-900.
36. Watanabe N, Che FS, Iwano M, Takayama S, Yoshida S, Isogai A. Dual targeting of spinach protoporphyrinogen oxidase II to mitochondria and chloroplasts by alternative use of two in-frame initiation codons. J Biol Chem. 2001;276(23):20474-81.
37. Woodson JD, Perez-Ruiz JM, Chory J. Heme synthesis by plastid ferrochelatase I regulates nuclear gene expression in plants. Curr Biol. 2011;21(10):897-903.
38. Singh DP, Cornah JE, Hadingham S, Smith AG. Expression analysis of the two ferrochelatase genes in Arabidopsis in different tissues and under stress conditions reveals their different roles in haem biosynthesis. Plant Mol Biol. 2002;50(4-5):773-88.
39. Kang K, Lee K, Park S, Lee S, Kim YS, Back K. Overexpression of rice ferrochelatase I and II leads to increased susceptibility to oxyfluorfen herbicide in transgenic rice. J Plant Biol. 2010;53(4):291-6.
40. Emanuelsson O, Nielsen H, Brunak S, von Heijne G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J Mol Biol. 2000;300(4):1005-16.
41. Claros MG, Vincens P. Computational method to predict mitochondrially imported proteins and their targeting sequences. Eur J Biochem. 1996;241(3):779-86.
42. Bannai H, Tamada Y, Maruyama O, Nakai K, Miyano S. Extensive feature detection of N-terminal protein sorting signals. Bioinformatics. 2002;18(2):298-305.
43. Hoglund A, Donnes P, Blum T, Adolph HW, Kohlbacher O. MultiLoc: prediction of protein subcellular localization using N-terminal targeting sequences, sequence motifs and amino acid composition. Bioinformatics. 2006;22(10):1158-65.
44. Whitcombe DM, Carter NP, Albertson DG, Smith SJ, Rhodes DA, Cox TM. Assignment of the human ferrochelatase gene (FECH) and a locus for protoporphyria to chromosome 18q22. Genomics. 1991;11(4):1152-4.
45. Watanabe S, Hanaoka M, Ohba Y, Ono T, Ohnuma M, Yoshikawa H, et al. Mitochondrial localization of ferrochelatase in a red alga Cyanidioschyzon merolae. Plant Cell Physiol. 2013;54(8):1289-95.
46. Camadro JM, Labbe P. Purification and properties of ferrochelatase from the yeast Saccharomyces cerevisiae. Evidence for a precursor form of the protein. J Biol Chem. 1988;263(24):11675-82.
47. Horie C, Suzuki H, Sakaguchi M, Mihara K. Targeting and assembly of mitochondrial tail-anchored protein Tom5 to the TOM complex depend on a signal distinct from that of tail-anchored proteins dispersed in the membrane. J Biol Chem. 2003;278(42):41462-71.
48. Hansson M, Hederstedt L. Purification and characterisation of a water-soluble ferrochelatase from Bacillus subtilis. Eur J Biochem. 1994;220(1):201-8.
49. Ferreira GC, Franco R, Lloyd SG, Moura I, Moura JJ, Huynh BH. Structure and function of ferrochelatase. J Bioenerg Biomembr. 1995;27(2):221-9.
50. Krogh A, Larsson B, von Heijne G, Sonnhammer EL. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J Mol Biol. 2001;305(3):567-80.
51. Cserzo M, Eisenhaber F, Eisenhaber B, Simon I. TM or not TM: transmembrane protein prediction with low false positive rate using DAS-TMfilter. Bioinformatics. 2004;20(1):136-7.
52. Tusnady GE, Simon I. The HMMTOP transmembrane topology prediction server. Bioinformatics. 2001;17(9):849-50.
53. Claros MG, von Heijne G. TopPred II: an improved software for membrane protein structure predictions. Comput Appl Biosci. 1994;10(6):685-6
54. Sobotka R, Tichy M, Wilde A, Hunter CN. Functional assignments for the carboxyl-terminal domains of the ferrochelatase from Synechocystis PCC 6803: the CAB domain plays a regulatory role, and region II is essential for catalysis. Plant Physiol. 2011;155(4):1735-47.
55. Sobotka R, McLean S, Zuberova M, Hunter CN, Tichy M. The C-terminal extension of ferrochelatase is critical for enzyme activity and for functioning of the tetrapyrrole pathway in Synechocystis strain PCC 6803. J Bacteriol. 2008;190(6):2086-95.
56. Taketani S, Tanaka-Yoshioka A, Masaki R, Tashiro Y, Tokunaga R. Association of ferrochelatase with Complex I in bovine heart mitochondria. Biochim Biophys Acta. 1986;883(2):277-83.
57. Al-Karadaghi S, Hansson M, Nikonov S, Jonsson B, Hederstedt L. Crystal structure of ferrochelatase: the terminal enzyme in heme biosynthesis. Structure. 1997;5(11):1501-10.
58. Thomas J, Weinstein JD. Measurement of heme efflux and heme content in isolated developing chloroplasts. Plant Physiol. 1990;94(3):1414-23.
59. Daley DO, Whelan J. Why genes persist in organelle genomes. Genome Biol. 2005;6(5):110.
60. Sato S, Nakamura Y, Kaneko T, Asamizu E, Tabata S. Complete structure of the chloroplast genome of Arabidopsis thaliana. DNA Res. 1999;6(5):283-90.
61. Jung HS, Chory J. Signaling between chloroplasts and the nucleus: can a systems biology approach bring clarity to a complex and highly regulated pathway? Plant Physiology. 2010;152(2):453-9.
62. Millar AH, Whelan J, Soole KL, Day DA. Organization and regulation of mitochondrial respiration in plants. Annual Rev Plant Biol. 2011;62:79-104.
63. Kleine T, Maier UG, Leister D. DNA transfer from organelles to the nucleus: the idiosyncratic genetics of endosymbiosis. Annual Rev Plant Biol. 2009;60:115-38.
64. Mense SM, Zhang L. Heme: a versatile signaling molecule controlling the activities of diverse regulators ranging from transcription factors to MAP kinases. Cell Res. 2006;16(8):681-92.
65. Nott A, Jung HS, Koussevitzky S, Chory J. Plastid-to-nucleus retrograde signaling. Annual Rev Plant Biol. 2006;57:739-59.
66. Kobayashi Y, Kanesaki Y, Tanaka A, Kuroiwa H, Kuroiwa T, Tanaka K. Tetrapyrrole signal as a cell-cycle coordinator from organelle to nuclear DNA replication in plant cells. Proc Natl Acad Sci USA. 2009;106(3):803-7.
67. Mochizuki N, Tanaka R, Tanaka A, Masuda T, Nagatani A. The steady-state level of Mg-protoporphyrin IX is not a determinant of plastid-to-nucleus signaling in Arabidopsis. Proc Natl Acad Sci USA. 2008;105(39):15184-9.
68. Moulin M, McCormac AC, Terry MJ, Smith AG. Tetrapyrrole profiling in Arabidopsis seedlings reveals that retrograde plastid nuclear signaling is not due to Mg-protoporphyrin IX accumulation. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(39):15178-83.
69. Padmanaban G, Venkateswar V, Rangarajan PN. Haem as a multifunctional regulator. Trends Biochem Sci. 1989;14(12):492-6.
70. Grover A, Pareek A, Singla SL, Minhas D, Katiyar S, Ghawana S, et al. Engineering crops for tolerance against abiotic stress through gene manipulation. Current Science. 1998;75(7):689-96.
71. Avin-Wittenberg T, Tzin V, Angelovici R, Less H, Galili G. Deciphering energy-associated gene networks operating in the response of Arabidopsis plants to stress and nutritional cues. Plant J. 2012;70(6):954-66.
72. Pasqualini S, Piccioni C, Reale L, Ederli L, Della Torre G, Ferranti F. Ozone-induced cell death in tobacco cultivar Bel W3 plants. The role of programmed cell death in lesion formation. Plant Physiol. 2003;133(3):1122-34.
73. Nagai S, Koide M, Takahashi S, Kikuta A, Aono M, Sasaki-Sekimoto Y, et al. Induction of isoforms of tetrapyrrole biosynthetic enzymes, AtHEMA2 and AtFC1, under stress conditions and their physiological functions in Arabidopsis. Plant Physiol. 2007;144(2):1039-51.
74. Marschner H. Mineral nutrition of higher plants. 2nd ed. London: Academic Press; 1995.
75. Curie C, Panaviene Z, Loulergue C, Dellaporta SL, Briat JF, Walker EL. Maize yellow stripe1 encodes a membrane protein directly involved in Fe(III) uptake. Nature. 2001;409(6818):346-9.
76. Le Jean M, Schikora A, Mari S, Briat JF, Curie C. A loss-of-function mutation in AtYSL1 reveals its role in iron and nicotianamine seed loading. Plant J. 2005;44(5):769-82.
77. Lee S, Kim YS, Jeon US, Kim YK, Schjoerring JK, An G. Activation of Rice nicotianamine synthase 2 (OsNAS2) enhances iron availability for biofortification. Mol Cells. 2012;33(3):269-75.
78. Hider RC, Kong X. Iron speciation in the cytosol: an overview. Dalton Trans. 2013;42(9):3220-9.
79. Duy D, Wanner G, Meda AR, von Wiren N, Soll J, Philippar K. PIC1, an ancient permease in Arabidopsis chloroplasts, mediates iron transport. Plant Cell. 2007;19(3):986-1006.
80. Jeong J, Cohu C, Kerkeb L, Pilon M, Connolly EL, Guerinot ML. Chloroplast Fe(III) chelate reductase activity is essential for seedling viability under iron limiting conditions. Proc Natl Acad Sci USA. 2008;105(30):10619-24.
81. Jeong J, Connolly EL. Iron uptake mechanisms in plants: Functions of the FRO family of ferric reductases. Plant Sci. 2009;176(6):709-14.
82. Shingles R, North M, McCarty RE. Ferrous ion transport across chloroplast inner envelope membranes. Plant Physiol. 2002;128(3):1022-30.
83. Bashir K, Ishimaru Y, Shimo H, Nagasaka S, Fujimoto M, Takanashi H, et al. The rice mitochondrial iron transporter is essential for plant growth. Nature Commu. 2011;2:322.
84. Maliandi MV, Busi MV, Turowski VR, Leaden L, Araya A, Gomez-Casati DF. The mitochondrial protein frataxin is essential for heme biosynthesis in plants. FEBS J. 2011;278(3):470-81.
85. Briat JF. Roles of ferritin in plants. J Plant Nutr. 1996;19(8-9):1331-42.
86. Ravet K, Touraine B, Boucherez J, Briat JF, Gaymard F, Cellier F. Ferritins control interaction between iron homeostasis and oxidative stress in Arabidopsis. Plant J. 2009;57:400-12.
87. Arosio P, Ingrassia R, Cavadini P. Ferritins: a family of molecules for iron storage, antioxidation and more. Biochim Biophys Acta. 2009;1790(7):589-99.
88. Harrison PM, Arosio P. The ferritins: molecular properties, iron storage function and cellular regulation. Biochim Biophys Acta. 1996;1275(3):161-203.
89. Tarantino D, Casagrande F, Soave C, Murgia I. Knocking out of the mitochondrial AtFer4 ferritin does not alter response of Arabidopsis plants to abiotic stresses. J Plant Physiol. 2010;167(6):453-60.
90. Goto F, Yoshihara T, Shigemoto N, Toki S, Takaiwa F. Iron fortification of rice seed by the soybean ferritin gene. Nature Biotechnol. 1999;17(3):282-6.
91. Kheir W, Ismail G, El-Nour FA, Tawfik T, Hammad D. Assessment of the efficiency of duckweed (Lemna gibba) in waste water treatment. International J Agric Biol. 2007;9(5):681-7.
92. Teixeira S, Vieira MN, Espinha Marques J, Pereira R. Bioremediation of an iron-rich mine effluent by Lemna minor. Int J Phytoremediation. 2014;16(7-12):1228-40.
93. Lee S, An G. Over-expression of OsIRT1 leads to increased iron and zinc accumulations in rice. Plant Cell Environ. 2009;32(4):408-16.
94. Ishimaru Y, Masuda H, Bashir K, Inoue H, Tsukamoto T, Takahashi M, et al. Rice metal-nicotianamine transporter, OsYSL2, is required for the long-distance transport of iron and manganese. Plant J. 2010;62(3):379-90.
95. Margulis L. Symbiosis and evolution. Sci Am. 1971;225:48-57.
96. Obornik M, Green BR. Mosaic origin of the heme biosynthesis pathway in photosynthetic eukaryotes. Mol Biol Evol. 2005;22(12):2343-53.
97. Weeden NF. Genetic and biochemical implications of the endosymbiotic origin of the chloroplast. J Mol Evol. 1981;17(3):133-9.
98. Gould SB, Waller RF, McFadden GI. Plastid evolution. Annual Rev Plant Biol. 2008;59:491-517.
99. Keeling PJ. Diversity and evolutionary history of plastids and their host. American Journal of Botany. 2004;91(10):1481-93.
100. Judd WS, Campbell CS, Kellogg EA, Stevens PF, Donoghue MJ. Plant systematics. A phylogenetic approach. 3rd ed. Sunderland, Massachusetts: Sinauer Associates; 2008.
101. Benson DA, Karsch-Mizrachi I, Lipman DJ, Ostell J, Sayers EW. GenBank. Nucleic Acids Res. 2011;39(Database issue):D32-7.
102. Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997;25(24):4876-82.
103. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28(10):2731-9.
104. Zimmermann MB, Hurrell RF. Improving iron, zinc and vitamin A nutrition through plant biotechnology. Curr Opin Biotechnol. 2002;13(2):142-5.
105. Briat JF, Curie C, Gaymard F. Iron utilization and metabolism in plants. Curr Opin Plant Biol. 2007;10:276-82.
Received in April, 2014.
Accepted in July, 2014.
Eduardo Ortega. Laboratorio de Fisiología Vegetal, Departamento de Biología Vegetal, Facultad de Biología, Universidad de La Habana, La Habana, Cuba. E-mail: eortega@fq.uh.cu.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}