










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.31 no.3 La Habana jul.-set. 2014
REVIEW
Current trends and perspectives in veterinary vaccine production
Estado actual sobre la producción de vacunas veterinarias, tendencias y proyecciones
Aníbal Domínguez1, Rafael Polanco1, Gema Cossío2, Yolanda Morejón2, Yanelis Riquenes1
1 Dirección de Investigación y Desarrollo. Grupo Empresarial Labiofam. Avenida Independencia km 16 ½, Boyeros, La Habana, Cuba.
2 Consultoría BioMundi, Dirección de Inteligencia Corporativa, Instituto de Información Científica y Tecnológica, IDICT, Ministerio de Ciencia, Tecnología y Medio Ambiente, Citma. La Habana, Cuba.
ABSTRACT
The dynamism observed in the area of veterinary vaccinology needs to be monitored by the biopharmaceutical industry, thus, to anticipate to the possible changes and take actions that allow it to adapt to the future scenario. On this basis, a study was performed to assess the current status, trends, scientific and technological projections in the production of veterinary vaccines. To this end, a topic search was done on vaccines for poultry, cattle, pigs, rabbits, horses, fish and dogs, in the Scopus database from 2008 to 2012 and further contextualized to early 2014. It was found that veterinary vaccinology is a booming field, with the highest growth in the year 2011 with 18.05 % compared to 2008. As such, the US and China are leading this issue, having a major scientific interest in protein antigens, adjuvants and viral vaccines produced with conventional technologies. The most attended species are birds, pigs and cattle, the first species with 15.21 % of all publications. We conclude that the dominance of the classical vaccine production technologies will continue despite the advances in genetic engineering and biotechnology. Vaccines produced with modern technologies seem to be, at least for now, less affordable for the producer, and therefore impractical. Similarly, the next few years will be very important for the development and registration of new vaccines obtained by DIVA technology and their diagnostic systems.
Keywords: veterinary vaccines, vaccination, production technology, adjuvant.
RESUMEN
El dinamismo observado en el área de la vacunología veterinaria requiere ser monitoreado por la industria biofarmacéutica, para de esta forma anticiparse a los posibles cambios y tomar acciones que le permita adaptarse al escenario futuro. Sobre esa base se realizó un estudio con el objetivo de evaluar el estado actual, tendencias y proyecciones científico-tecnológicas en la producción de vacunas veterinarias. A tal efecto se efectuó una búsqueda temática en la base de datos Scopus sobre vacunas destinadas a aves, bovinos, cerdos, conejos, equinos, peces y perros, limitada al periodo 2008-2012 y se contextualizó hasta principios de 2014. Se comprobó que la vacunología veterinaria es un campo temático en plena expansión, siendo el año 2011 el de mayor crecimiento con un 18.05 % con respecto al 2008. En tal sentido Estados Unidos y China lideran esta temática, existiendo un interés científico mayoritario por los antígenos proteicos, adyuvantes y vacunas virales producidas con tecnologías clásicas. Las especies más atendidas son las aves, porcinos y bovinos, sobresaliendo la primera especie con el 15.21 % de todas las publicaciones. Se concluye que continuara el dominio de las tecnologías clásicas de producción de vacunas a pesar de los adelantos en materia de ingeniería genética y biotecnología. Las vacunas producidas con tecnologías modernas parecen ser, al menos por ahora, poco asequibles algunos productores, y por tanto, poco prácticas. De igual manera los próximos años serán muy importantes para el desarrollo y el registro de nuevas vacunas obtenidas por tecnología DIVA y sus respectivos sistemas de diagnóstico
Palabras clave: vacunas veterinarias, vacunación, tecnología de producción, adyuvantes.
INTRODUCTION
Veterinary vaccines are biological preparations designed from infectious agents and aimed at developing a protective immunological response against them in healthy animals once administered. These formulations resemble the pathogen’s natural infection but do not develop the disease, mounting a protective, effective and long-lasting immune response. From that moment on, the vaccinated organism not only survives to a second contact with the agent, but also breaks the transmission chain. For these reasons, vaccination has been considered one of the most efficacious sanitary measures to prevent, control and eradicate infectious diseases in domestic and wild-life animals, and one of the factors available to face the increasing demand for food supplies and food dependency [1].
Nevertheless, the epidemiological situation cannot be solved only through vaccination, but complementing it with sanitary measures such as: outbreak containment, control of animal transportation and biosafety measures, among others.
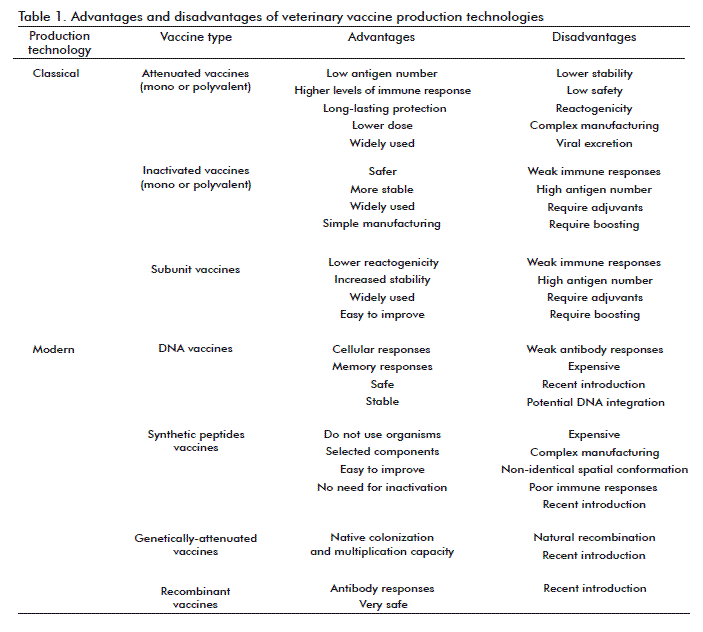
There are several classification criteria for vaccines attending to its administration route, composition, target biological agent, the technology for its production, and others. Since the technological point of view, there are two main types: classical vaccines and modern vaccines (Table 1) [2].
The ‘One health’ concept has made difficult to establish differences between human and veterinary vaccines, as certainly are. Similarities arise from the shared properties of the immune systems in both types of organisms: highly specific immune responses and immunological memory. This convergence favors that most of the knowledge and current results on human vaccines obtained in animal models could be subsequently applied in veterinary vaccines manufacturing [3].
The cost-benefit balance both for the farmer and the industry arises as the paramount factor determining the manufacture and practical use of veterinary vaccines compared to vaccines designed for human use. It is so remarkable that makes irrelevant considerations regarding the etiological agent to be controlled or the target animal species to be protected. Therefore, it determines in the clinical practice that vaccines with low production costs not only decrease zoosanitary complications, and productivity and cost limitations, but also, and significantly, producers can afford it [4].
All these properties have fostered the search for new knowledge on animal immunology, microbiology, vaccine production technologies, together with the development of new animal models for human diseases and the recent vertiginous increase on veterinary vaccinology. However, such dynamism requires to be monitored by the veterinary vaccines industry. In this work some of the trends observed in the scientific literature for vaccine research, development and production worldwide in the last five years are provided.
GLOBAL FOCUS ON CURRENT DEVELOPMENTS
The most recent scientific information flow has to be properly covered to identify the probable behavior of the scientific field in short term (5-10 years). For that purpose, a topic search was made in the Scopus database on vaccines about poultry, cattle, swine, rabbit, horse, fish and dogs, from 2008 to 2012. The title, abstract and keywords bibliographic fields were searched for the most relevant technological- and infectious agent-specific terms. The search strategy included the most common denominative terms for each species: poultry (bird, avian, pigeon, poultry), cattle (cattle, calf, bovine, cow), pig (pig, hog, swine, porcine), horse (horse, equine), fish (fish), dogs (dog, canine) and rabbits (rabbit).
A detailed analysis of the articles retrieved on vaccine production for these species, vaccine technologies and specific pathogens supported the view on the novelty of the information and the vertiginous expansion of the veterinary vaccine field of research. In this sense, there was a steady increase in the articles published from 2008 to 2012, representing an 18.05 % of increase in 2011 compared to 2008 for all the species, but in rabbits, which decreased in 2010 to its lowest point (47.30 %).
It could be inferred from the data that there was a disproportionate research attention worldwide to the species under study (Figure 1). Approximately 34 % of the articles indexed in Scopus versed on poultry, pigs and cattle vaccines, species comprising economically relevant animals, with 15.21 % of the articles on poultry.

This behavior was closely influenced by the morbimortality of some diseases during the period, and by the relative relevance of some of these species for human consumption, particularly meat and its derivatives (Figure 2). In this regard, there were also some significant emergent zoonotic diseases of potential human transmission. Poultry was an example of that scenario, where both factors converged, providing the highest numbers of vaccine studies, particularly in broiler chickens, laying hens, turkeys and ducks.

Remarkably, poultry accounted for up to 28 % of the worldwide meat supply for consumption, as compared to swine and cattle (34 % and 20 %, respectively). Derivatives from these animal species has to be also considered, such as whole milk (78 %) and eggs (14 %).
These figures may explain the fact that the leading countries of the meat and egg production and export industry were the ones providing the largest scientific production as determined. The main contribution for the seven species under study was made by researchers from US and China universities, and articles on poultry, swine and cattle vaccines predominated. Particularly in fish vaccines, China got ahead of US, and both together with South Korea and India dominated this area. This could be explained by the role of fish for traditional diet in these countries and also its economic relevance.
Similarly, the production on viral vaccines almost doubled that on bacterial vaccines, except for aquaculture. This last could be explained by the fact that most bacteria pathogenic for fish can be isolated both in salt and fresh water and display a wide host range, including man. That is the case of Edwardsiella tarda, Streptococcus iniae, Aeromonas hydrophila, Lactococcus garvieae, Vibrio harveyi and V. anguillarum. Those bacteria are being targeted by monovalent and bivalent vaccines under research or development, and mostly but not exclusively by classical vaccine production technologies [5-8].
The disproportionate scientific production on viral versus bacterial veterinary vaccines can also be related to the availability of therapeutic alternatives affordable to producers, and to the numerous efforts on optimizing the use of antibacterial chemotherapy. Undoubtedly, another factor influencing the higher production on viral veterinary vaccines in this period was the numerous highly- or mildly-pathogenic isolates of the influenza virus emerging since 2003 [9]. The fast geographic spread of this virus isolates and their easiness to cross the interspecies barrier provoked complex epidemiological situations in many countries in Asia, Europe, Africa and the American continent. That situation consequently impacted with a 29 % increase in research and 15 % in publications on veterinary vaccines against influenza. Noteworthy, the articles on influenza related to humans, flying mammals, animals in captivity, wildlife birds (aquatic, fresh water, marine, marshes, swamps, predatory, migratory or from urban areas), were excluded from the current analysis.
The abovementioned increase resulted from several factors, such as: the large number of susceptible economically relevant species, the genetic instability of the infectious agent (mainly genetic mutation and recombination), and the vaccination strategies for disease control. Major topics comprised the use of antigens from several subtypes to generate classic vaccines mostly, vaccine testing in target animals, and vaccine adjuvant formulations [10].
Either bacterial or viral in origin, the infecto-contagious diseases targeted for the development of preventive, effective and safe protection cause colossal losses and increase restrictions to the international trade of animal meat, eggs and other derivatives as leather.
Those agents have in common:
- 1. There are no treatments available and, when an outbreak occurs, it is difficult to contain and costly its eradication (sacrifice);
- 2. They decrease productivity, animal growth and daily weight gain in susceptible animals;
- 3. Cause reproductive failures;
- 4. Induce high mortality;
- 5. Increase production costs, due to the high cost of resources for their screening, treatment and control;
- 6. Strict international restrictions are imposed to the trade of meat and derivatives immediately upon diagnosis; and
- 7. They affect several countries and species at a time, involving domestic, captive and wild species, even man.
Co-occurrence analyses of specific topic terms allowed to confirm that the field of veterinary vaccines is composed of complex and highly interconnected knowledge clusters (Figure 3). Protein, virus and adjuvants were the key terms identified as central topics.
Their centrality, together with their interconnections, indicated that much of the original results published in the period 2008-2012 were mostly but not exclusively on obtaining new protein and peptidic viral antigens by different methods, the characterization of the generated immune responses in target species and animal models (poultry, swine and bovine), and particularly viral vaccine strains and new adjuvants.
In this last, there was increased interest to find new and better adjuvants, cheaper, more stable and safe, and able to stimulate both the humoral and cellular branches of the immune response, to reduce the administered antigen dose and the number of boosts required to develop a protective response. Another incentive fostering the research in this area is the ability of these compounds to be included as part of vaccine formulations to be delivered by mucosal route, particularly against diseases transmitted by this route.
Montanide adjuvants are among those being actively investigated during the period, in poultry, pigs and bovines [11]. Nevertheless, its equivalence or superiority compared to alum hydroxide gels still remains to be demonstrated in clinical practice, together with its approval for commercial use.
All these suggest that, first, classical technologies for vaccine production are well established for veterinary vaccines research and development. Second, that there are huge efforts ongoing to improve vaccine formulations elaborated with classical technologies, to adapt them to the emergent epidemiological situations and to favor their compliance with the ultimate goal of any vaccination against an infectious disease: the long lasting resistance against infection. And third, the entanglement of a coordinated international framework to keep vaccination as the short- and long-term best choice for the control of infectious diseases.
In the same direction, a new technology was developed combining benefits from classic and modern vaccine technologies: DIVA (differentiating infected from vaccinated animals). There are few products available based on this technology, but they have demonstrated to solve a remarkable issue of classical vaccines: the induction through vaccination of a distinctive immune response different from that produced by the wild strain in the target animal. DIVA also provides as additional advantage the plasticity to build a vaccine by any of three possible strategies:
1. Negative marker or deletion vaccine: based on the absence of specific antigen epitopes present in the wild type virus.
2. Extrinsic positive marker: by including in the vaccine an immunodominant epitope different from that of the infectious agent.
3. Intrinsic positive marker: by forming an epitope from the wild type virus that induces different antibody patterns.
A particular advantage of DIVA technology resides in the diagnostic test included with the vaccine, of high specificity and sensitivity, and able to differentiate natural from vaccine antibody responses in sera, plasma, mucosal secretions and milk samples. This eliminates the urgent need of sacrificing large animal populations suspected to be infected, provides an effective way for fast outbreak detection by massive screening, and shortens the time required to declare the outbreak as controlled and demonstrate the lack of infection [12]. All these reasons make of DIVA technology a major unprecedented achievement in veterinary vaccinology of profound ethical, environmental and social implications for the animal well-being and global food safety.
Other differentiating aspects of this technology in respect to classical and modern vaccine technologies at the time of this work were: the absence of studies using it to prevent bacterial diseases or targeting diseases in poultry, the easiness of using the vaccine and its diagnostic test in countries where the target disease is non-endemic, and the large pharmaceutical companies worldwide as the leaders in research and development of this type of vaccines in detriment of research institutes and universities (Table 2). Among the most significant producers are: Merck Animal Health (USA), Bayer Schering AG (Germany), Pfizer Animal Health (USA), Boehringer Ingelheim Animal Health GmbH (Germany) and Merial Limited (USA). This differentiating aspect could be speeding up the introduction of DIVA products in the clinical practice, both the vaccines and the diagnostic tests. There is one exception, with the National Institute for Agricultural Technology (INTA) in Argentina, a government institution which owns the patent of the HVB-1gE labeled vaccine against the bovine herpesvirus type 1, the etiological agent of the infectious bovine rhinotracheitis, the infectious pustular vulvovaginitis and the infectious pustular balanoposthitis, all of them affecting bovine at any age, either wild or domestic.

POULTRY VIRAL VACCINES
The broiler chicken was the model receiving the major research focus, followed by laying hens and turkeys. Similarly, most publications were about Marek´s, Newcastle and Gumboro diseases and the avian infectious bronchitis. Vaccines against these diseases were produced by using classical technologies, which have been essential for control programs and to prevent outbreaks worldwide. The vaccine formulations were developed mostly to be orally administered, by injection or in ovo. This last vaccination modality has several advantages, including the development of an early immune response and supports the automation of the vaccination campaign, decreasing the human error and also the need for labor force [13-17].
Otherwise, the weaknesses of these vaccine formulations vary according to the model. Marek’s disease vaccines are produced by cell culture technologies, are expensive and require careful handling and administration procedures, in spite of been efficacious (95 % of protection) even in the presence of maternal antibodies, safe, stable and inducing long lasting immunity. Another example, the Newcastle disease vaccines produced by the live virus technology, could interfere with the action of other vaccines against respiratory diseases, particularly those against the infectious bronchitis. When these two vaccines are jointly administered, they compete for the same receptors at the trachea, therefore, affecting the development of immunity. This is very hard to solve in broiler chickens, due to their shorter lifespan [14]. In the case of Gumboro disease, current vaccine formulations based on the strain 2512 effectively protected the animals from clinical symptoms, but they cause bursal damage due to viral replication in the bursa of Fabricius [15, 16].
The abomentioned problems of classical vaccine technologies, together with their interferences in diagnosis and the new developments in molecular biology techniques, fostered new lines of research. Some of them comprised the genomics and proteomics of emergent strains of variable degree of virulence, and new vaccination strategies. There were also attempts to generate DNA vaccines, vector-based and genetically attenuated vaccines, and vaccines encapsulated in nanoparticles, but still having a long way to go until its approval by regulatory authorities [13, 14, 16-21]. Nevertheless, none of the new options has achieved the protection conferred by conventional vaccines against the Marek’s disease. By the contrary, recombinant vaccines (for example, against Newcastle and Gumboro diseases) seem to have overcome the disadvantages of classical vaccines and have neither caused collateral damage nor interfered with other vaccines or maternal antibodies [19, 20].
PORCINE VIRAL VACCINES
Regarding porcine vaccines, major efforts worldwide were intended to control the porcine reproductive and respiratory syndrome, circoviruses and the classical swine fever, with classical vaccine technologies as the most prominent for commercial vaccine production. This type of vaccines has been recognized as a key factor for the success over a decade of the control programs in swine industry worldwide [22-25].
Major inconveniences of classical vaccines against swine viral diseases are technology-related. A particular case comprises inactivated and live-attenuated vaccines available against the porcine reproductive and respiratory syndrome virus. Inactivated vaccines seem to be inadequate in pregnant animals and boars, in the last due to the potential elimination of the vaccine virus through semen. Live-attenuated viruses can persist after vaccination and could be transmitted to unvaccinated animals, causing future outbreaks, as documented. Moreover, as the case of live-attenuated vaccines against the classical swine fever, sick animals are impossible to be serologically distinguished from the vaccinated animals, the vaccine generates a teratogenic effect and the vaccine viral strain also continue to circulate among the vaccinated swine population [26].
Such limitations have being exacerbated in recent years with the intensification of the swine industry, with important changes in the virulence, clinical manifestations and pathogenesis of many infectious agents. This complex situation has been promoted by the pathogen’s genetic variations, modifications in the microbial ecology of production farms, the simultaneous occurrence of infections and the outcome of atypical and unnoticed variants of the diseases [26]. In this complex context there are ongoing efforts to develop molecular techniques that would result in new generations of safer and more effective prophylactic vaccines, able to be used in emergent vaccination campaigns.
This complex scenario has been confronted by a set of strategies, particularly against the porcine reproductive and respiratory syndrome, with promising preclinical satisfactory results (immunogenicity and immunological protection) using a DNA vaccine that expresses the gp3 and gp5 glycoproteins of the virus genotype I [27]. Regarding the control of classical swine fever in susceptible populations, several candidates have been developed during the last 15 years against it. They include: recombinant deletion chimeric vaccines, DNA vaccines, subunit vaccines and vector-based, among others. Many of them did not comply with the requirements of regulatory agencies upon its evaluation, in spite of inducing potent immune responses. Most of the efforts continue targeting the E2 glycoprotein as the main immunogen of this virus and a critical component of DIVA vaccines [28-30].
Circovirus vaccines has been fueled-up by the successful experimental results using subunit vaccines based on serotype 2 and chimeric vaccines formulated from the 1-2a serotype, being intramuscularly and orally administered [31, 32]. In the case of the porcine reproductive and respiratory syndrome, promising results were obtained by using recombinant vaccines, DNA vaccines and subunit vaccines produced in vegetables with the additional advantage of been administered by mucosal route [33, 34]. In spite of such advances against this disease, the quest for a better vaccine is still ongoing, as required to decrease the infection by the PCV2 virus to undetectable levels once after interrupting vaccination.
The Aujeszky’s disease vaccine is worth to be mentioned, due to the increasing number of DIVA vaccines available in the market against this disease and because of being among the first vaccines of this kind approved to be used in Europe. This could have resulted from the virus capacity to infect several mammalian species, including economically relevant ones (swine, bovine, rabbits), domestic animals (dogs, cats) and wild species (wild boar, foxes, bisons). Most of these vaccines were developed from the NIA-3 or Bartha-K61 strains, both of them having the glycoprotein E gene deleted, supporting the generation of a live-attenuated viral vaccine and protecting at the same time against the clinical symptoms of the disease. This last considerably reduces the levels of excreted virus, making the vaccine safer and differentiating vaccinated from infected animals, all these aspects unachieved by live vaccines [35, 36].
BOVINE VIRAL VACCINES
The first depicting element of vaccine research in this species is the high number of studies on anti-tick vaccines. This exception comes from the role that hematophagous parasite has as vector to transmit diseases such as babesiosis and anaplasmosis, in addition to the direct weakening of the animals and its detrimental effects on production and causing host animal death. Other factors influencing this behavior were the high costs of chemical acaricides, their increased toxicity for both the animal and the environment and problems during storage. The scientific interest was mainly focused on finding and characterizing antigenic proteins or cryptic antigens of the digestive tract of these insects, which could be able to induce a protective, safe and lost lasting immune response in vaccinated animals [37-39].
Regarding the most studied viruses, there was a significant attention worldwide to the foot and mouth disease virus, the bovine viral diarrhea and the bluetongue disease virus. There were recurrent statements on the effectiveness of inactivated and live-attenuated vaccines in the different national control programs enforced for the long-term containment of these diseases [40-45]. Nevertheless, current vaccine formulations bear some weaknesses, such as the coexistence of several viral strains of the foot and mouth disease virus (serotypes O, A, C, SAT 1, 2 and 3, Asia 1; comprising more than 60 strains) devoid of immune response crossreactivity between them. This comes from the fact that most of those strains comprise live-attenuated viruses which use is forbidden in the countries where they are used, and there are technical limitations to differentially detect infected from immunized animals. From 2008 to 2013, significant improvements were achieved, particularly on the knowledge of viral structure and the development of subunit, DNA and DIVA vaccines, based on SP and NSP proteins [46-50]. Once again, DIVA was the most promising development in spite of some inconsistencies.
The bovine diarrhea virus was also investigated, mainly due to the lack of a standard vaccine but with several commercial preparations available based on classical vaccine technologies but with certain limitations. Besides the technically-associated limitations, the live virus vaccine induces immunosuppression and is not recommended to be administered to pregnant cattle to avoid persistent infections, particularly in fetuses [51]. The use of non-structural compounds as part of inactivated vaccines seems to be promising to solve such inconveniences [52].
The live-attenuated vaccines against bluetongue disease virus inconveniently cause embryo death and reabsorption, fetal death and congenital malformations in the offspring. Their use was rejected in many countries because they contain several serotypes that could revert the vaccine viral strain phenotype to a virulent form and facilitate its subsequent vector-borne transmission. That’s why the inactivated vaccines against this virus usually contain few serotypes, something that limits their protective spectrum [53]. Such difficulties have fostered the research on safer and more efficacious vaccines, able to induce crossreactive immunity and having differential diagnostic capacity. Several prototypes were developed, including recombinant vaccines, with potential wide-range crossreactivity among serotypes, but remaining to be validated in the field [54].
A key point comprises reemergent diseases affecting these species and of growing interest for the veterinary vaccinology community, as the bovine infectious rhinotraqueitis, caused by the bovine herpesvirus type 1. In this sense, improved attenuated vaccines were tested, as well as mono or polyvalent inactivated vaccines and its administration by different routes [55]. Commercially available vaccines against this virus based on conventional technologies are widely used in different variants: live-modified or inactivated virus vaccines, either mono or polyvalent. Its use depends on the capacity of the given vaccine to induce immunogenicity, its efficacy to reduce clinical manifestations of the disease and, consequently, the economic losses, even with an incomplete protection from infection. Additionally, live-attenuated vaccines could induce abortion, and the lifespan persistence of the vaccine strain in vaccinated animals could lead to potential reactivation of latent virus after corticosteroid immunosuppression and, ultimately, to infection.
Trying to solve those limitations, new vaccine preparations were investigated using new antigens such as glycoproteins D and E, combined with its delivery by different routes, one of them a recombinant vaccine by mucosal route [56], DNA vaccines [57] and DIVA.
Particularly on DIVA, it is worth to mention that it has become one of the most resourceful techniques for veterinary disease vaccination and eradication programs, with its associated serological tests. In fact, there are several products using this technology against the infectious bovine rhinotracheitis virus type 1 in the market, some of them adjuvanted with aluminum hydroxide. The most widely accepted strategy to generate DIVA vaccines against this disease worldwide includes the deletion of the glycoprotein E gene, differentiating the naturally infected animals which develop antibodies against this protein from the vaccinated ones. The deletion also favors the generation of live-attenuated vaccines that could be applied either to economically relevant or wild animals. Similarly, the potential advantage for reexcretion through nasal secretions of the vaccine strain after boosting provides it the additional advantage of establishing herd immunity [58].
Results demonstrated a reduced viral excretion after boosting and the early induction of immunity after vaccination by the intranasal route. Nevertheless, recent studies evidenced that the nasal coinfection with a gE-negative vaccine strain and a wild-type strain could generate a recombinant viral progeny of lower virulence in vitro compared to wild-type strains, but higher than that of the parental strains in vivo. In this sense, it was confirmed that recombination events could restore the natural virulence to the gE negative vaccine strains and limit its application under field conditions or in areas of widespread circulation of wild-type strains [59].
The Rift Valley fever disease is another example of the effectiveness of DIVA technology. This vector-borne disease similarly affects ruminants and humans, its causative agent being a class A, non-prioritized agent, and was classified as reemergent in Africa and the Middle East [59]. Current inactivated and live-attenuated vaccines obtained from the MP-12 strains and clone 13 inconveniently require up to three boosts, induce abortions, congenital malformations, hepatotoxicity and necrosis, in spite of been immunogenic and protective in vaccinated animals. Such inconveniences have forced the research on new vaccine candidates that could overcome those limitations or to provide alternatives [60, 61].
Once again, DIVA seems to be the most advanced and promising alternative, by the deletion of genes coding for NSm and NSs proteins. The vaccine induced differentiating immunity in rats and was shown to be innocuous in pregnant animals, what seems to comply with the expected requirements. Nevertheless, such advances do not represent the single solution to overcome limitations of classical vaccine technologies. Intense work is ongoing to develop new formulations using water/oil emulsions as adjuvants, vectored vaccines using Newcastle disease virus and poxviruses bearing the Gc and Gn genes coding for structural glycoproteins, and recombinant and subunit vaccines [62-65].
CONCLUSIONS
The detailed analysis of the articles published from 2008 to 2012 indexed in Scopus provides an overview of a renovated optimism of veterinary vaccines produced by classical technologies, in spite of improvements in the field of genetic engineering and biotechnology. The sanitary success achieved by classical technologies vaccines guarantee their predominance in the market in the near future. Such predictions are supported by their proved effectiveness and safety, low costs for research and development, better cost-benefits ratios and the availability of the production infrastructure required to face future technological needs for adjuvants and vaccine antigens.
Moreover, the opportunities provided by recombinant DNA techniques to generate new vaccines seems to be either less affordable or practical than required by producers. This is motivated by the high vaccination costs and the raise in standards to comply with regulatory requirements for sanitary registration, due to their recent introduction.
At the same time, DIVA technology and its related diagnostic tests will face significant challenges from development to registration. It is advantageous compared to other vaccine production technologies not only by its ability to safely and effectively control and prevent high incidence diseases, but also to avoid unnecessary sacrifice of uninfected animals and to control the reemergence of diseases declared as eradicated.
ACKNOWLEDGEMENTS
The authors thank to María de los Reyes Bermúdez, from the Agriculture Virtual Library of FAO Representation in Cuba, and to Omaida Maria Medina Alamo, specialist from the Direction of Research and Development, Labiofam Enterpreneurial Group, for their contributions to this work. Also to Ernesto Galbán Rodríguez, from the Elfos Scientiae publisher, for the critical review of the manuscript.
REFERENCES
1. Pastoret PP. Role of vaccination in animal health. Bull Acad Natl Med. 2012;196(3):589-90, 619-20.
2. López M, Mallorquín P, Pardo R, Vega M. Vacunas de nueva generación. Informe de vigilancia tecnológica. Madrid: Fundación Española para el Desarrollo de la Investigación en Genómica y Proteómica/Fundación General de la Universidad Autónoma de Madrid; 2004.
3. Williamson ED, Duchars MG, Kohberger R. Predictive models and correlates of protection for testing biodefence vaccines. Expert Rev Vaccines. 2010;9(5):527-37.
4. Lee NH, Lee JA, Park SY, Song CS, Choi IS, Lee JB. A review of vaccine development and research for industry animals in Korea. Clin Exp Vaccine Res. 2012;1(1):18-34.
5. Hu YH, Dang W, Deng T, Sun L. Edwardsiella tarda DnaK: expression, activity, and the basis for the construction of a bivalent live vaccine against E. tarda and Streptococcus iniae. Fish Shellfish Immunol. 2012;32(4):616-20.
6. Bastardo A, Ravelo C, Castro N, Calheiros J, Romalde JL. Effectiveness of bivalent vaccines against Aeromonas hydrophila and Lactococcus garvieae infections in rainbow trout Oncorhynchus mykiss (Walbaum). Fish Shellfish Immunol. 2012;32(5):756-61.
7. Hu YH, Deng T, Sun BG, Sun L. Development and efficacy of an attenuated Vibrio harveyi vaccine candidate with cross protectivity against Vibrio alginolyticus. Fish Shellfish Immunol. 2012;32(6):1155-61.
8. Hamod MA, Nithin MS, Shukur YN, Karunasagar I. Outer membrane protein K as a subunit vaccine against V. anguillarum. Aquaculture. 2012;354-355:107-10.
9. Li Y, Shi J, Zhong G, Deng G, Tian G, Ge J, et al. Continued evolution of H5N1 influenza viruses in wild birds, domestic poultry, and humans in China from 2004 to 2009. J Virol. 2010;84(17):8389-97.
10. Luo DY, Xue CY, Cao YC. Research progress and prospect of universal influenza vaccine. Bing Du Xue Bao. 2013;29(6):646-50.
11. Parker R, Deville S, Dupuis L, Bertrand F, Aucouturier J. Adjuvant formulation for veterinary vaccines: Montanide™ Gel safety profile. Procedia Vaccinol. 2009;1(1):140-7.
12. von Rosen T, Rangelova D, Nielsen J, Rasmussen TB, Uttenthal A. DIVA vaccine properties of the live chimeric pestivirus strain CP7_E2gif. Vet Microbiol. 2014;170(3-4):224-31.
13. Biggs PM, Nair V. The long view: 40 years of Marek's disease research and Avian Pathology. Avian Pathol. 2012;41(1):3-9.
14. Alexander DJ, Aldous EW, Fuller CM. The long view: a selective review of 40 years of Newcastle disease research. Avian Pathol. 2012;41(4):329-35.
15. León N, Icochea E, Gonzalez R, Perales R. Nivel de protección de una vacuna intermedia contra la enfermedad de Gumboro en aves de postura. Rev Investig Vet Perú. 2012;23(4):477-83.
16. Mahgoub HA, Bailey M, Kaiser P. An overview of infectious bursal disease. Arch Virol. 2012;157(11):2047-57.
17. Cook JK, Jackwood M, Jones RC. The long view: 40 years of infectious bronchitis research. Avian Pathol. 2012;41(3):239-50.
18. Zhang X, Wu Y, Huang Y, Liu X. Protection conferred by a recombinant Marek's disease virus that expresses the spike protein from infectious bronchitis virus in specific pathogen-free chicken. Virol J. 2012;9:85-91.
19. Zhao K, Chen G, Shi XM, Gao TT, Li W, Zhao Y, et al. Preparation and efficacy of a live newcastle disease virus vaccine encapsulated in chitosan nanoparticles. PLoS One. 2012;7(12):e53314.
20. Perozo F, Villegas P, Mavárez Y, Fernández R, Cruz J. Eficacia de un herpesvirus de pavo recombinante expresando la proteína vp2 del virus de la enfermedad de Gumboro ante un desafío experimental. Rev Cient 2010;20(2):132-7.
21. Britton P, Armesto M, Cavanagh D, Keep S. Modification of the avian coronavirus infectious bronchitis virus for vaccine development. Bioeng Bugs. 2012;3(2):114-9.
22. Zhai SL, Chen SN, Xu ZH, Tang MH, Wang FG, Li XJ, et al. Porcine circovirus type 2 in China: an update on and insights to its prevalence and control. Virol J. 2014;11:88.
23. Yang K, Li W, Niu H, Yan W, Liu X, Wang Y, et al. Efficacy of single dose of an inactivated porcine circovirus type 2 (PCV2) whole-virus vaccine with oil adjuvant in piglets. Acta Vet Scand. 2012;54:67.
24. Lee JA, Kwon B, Osorio FA, Pattnaik AK, Lee NH, Lee SW, et al. Protective humoral immune response induced by an inactivated porcine reproductive and respiratory syndrome virus expressing the hypo-glycosylated glycoprotein 5. Vaccine. 2014;32(29):3617-22.
25. Dortmans JC, Loeffen WL, Weerdmeester K, van der Poel WH, de Bruin MG. Efficacy of intradermally administrated E2 subunit vaccines in reducing horizontal transmission of classical swine fever virus. Vaccine. 2008;26(9):1235-42.
26. Blome S, Gabriel C, Beer M. Possibilities and limitations in veterinary vaccine development using the example of classical swine fever. Berl Munch Tierarztl Wochenschr. 2013;126(11-12):481-90.
27. Ren JQ, Sun WC, Lu HJ, Wen SB, Jing J, Yan FL, et al. Construction and immunogenicity of a DNA vaccine coexpressing GP3 and GP5 of genotype-I porcine reproductive and respiratory syndrome virus. BMC Vet Res. 2014;10(1):128.
28. Wang C, Sun Y, Qiu H. Progress in new-type vaccines against classical swine fever. Sheng Wu Gong Cheng Xue Bao. 2013;29(7):880-90.
29. Beer M, Reimann I, Hoffmann B, Depner K. Novel marker vaccines against classical swine fever. Vaccine. 2007;25(30):5665-70.
30. Tignon M, Kulcsar G, Haegeman A, Barna T, Fabian K, Levai R, et al. Classical swine fever: comparison of oronasal immunisation with CP7E2alf marker and C-strain vaccines in domestic pigs. Vet Microbiol. 2010;142(1-2):59-68.
31. Opriessnig T, Gomes-Neto JC, Hemann M, Shen HG, Beach NM, Huang Y, et al. An experimental live chimeric porcine circovirus 1-2a vaccine decreases porcine circovirus 2b viremia when administered intramuscularly or orally in a porcine circovirus 2b and porcine reproductive and respiratory syndrome virus dual-challenge model. Microbiol Immunol. 2011;55(12):863-73.
32. Feng H, Blanco G, Segales J, Sibila M. Can Porcine circovirus type 2 (PCV2) infection be eradicated by mass vaccination? Vet Microbiol. 2014;172(1-2):92-9.
33. Huang YW, Meng XJ. Novel strategies and approaches to develop the next generation of vaccines against porcine reproductive and respiratory syndrome virus (PRRSV). Virus Res. 2010;154(1-2):141-9.
34. Renukaradhya GJ, Dwivedi V, Manickam C, Binjawadagi B, Benfield D. Mucosal vaccines to prevent porcine reproductive and respiratory syndrome: a new perspective. Anim Health Res Rev. 2012;13(1):21-37.
35. Wang CH, Yuan J, Qin HY, Luo Y, Cong X, Li Y, et al. A novel gE-deleted pseudorabies virus (PRV) provides rapid and complete protection from lethal challenge with the PRV variant emerging in Bartha-K61-vaccinated swine population in China. Vaccine. 2014;32(27):3379-85.
36. Kong H, Zhang K, Liu Y, Shang Y, Wu B, Liu X. Attenuated live vaccine (Bartha-K16) caused pseudorabies (Aujeszky's disease) in sheep. Vet Res Commun. 2013;37(4):329-32.
37. Almazan C, Moreno-Cantu O, Moreno-Cid JA, Galindo RC, Canales M, Villar M, et al. Control of tick infestations in cattle vaccinated with bacterial membranes containing surface-exposed tick protective antigens. Vaccine. 2012;30(2):265-72.
38. Carreon D, de la Lastra JM, Almazan C, Canales M, Ruiz-Fons F, Boadella M, et al. Vaccination with BM86, subolesin and akirin protective antigens for the control of tick infestations in white tailed deer and red deer. Vaccine. 2012;30(2):273-9.
39. Guerrero FD, Miller RJ, Perez de Leon AA. Cattle tick vaccines: many candidate antigens, but will a commercially viable product emerge? Int J Parasitol. 2012;42(5):421-7.
40. Nagendrakumar SB, Srinivasan VA, Madhanmohan M, Yuvaraj S, Parida S, Di Nardo A, et al. Evaluation of cross-protection between O1 Manisa and O1 Campos in cattle vaccinated with foot-and-mouth disease virus vaccine incorporating different payloads of inactivated O1 Manisa antigen. Vaccine. 2011;29(10):1906-12.
41. Gonzalez AM, Arnaiz I, Yus E, Eiras C, Sanjuan M, Dieguez FJ. Evaluation of long-term antibody responses to two inactivated bovine viral diarrhoea virus (BVDV) vaccines. Vet J. 2014;199(3):424-8.
42. Charleston B. Eradicating bovine viral diarrhoea virus. Vet Rec. 2013;172(25):659-60.
43. Zientara S, Sanchez-Vizcaino JM. Control of bluetongue in Europe. Vet Microbiol. 2013;165(1-2):33-7.
44. Hasler B, Howe KS, Di Labio E, Schwermer H, Stark KD. Economic evaluation of the surveillance and intervention programme for bluetongue virus serotype 8 in Switzerland. Prev Vet Med. 2012;103(2-3):93-111.
45. Batten CA, Edwards L, Oura CA. Evaluation of the humoral immune responses in adult cattle and sheep, 4 and 2.5 years post-vaccination with a bluetongue serotype 8 inactivated vaccine. Vaccine. 2013;31(37):3783-5.
46. Jamal SM, Belsham GJ. Foot-and-mouth disease: past, present and future. Vet Res. 2013;44:116.
47. Smith MT, Bennett AM, Grubman MJ, Bundy BC. Foot-and-mouth disease: technical and political challenges to eradication. Vaccine. 2014;32(31):3902-8.
48. Yang B, Yang F, Wang SH, Zhang Y, Cao WJ, Yin H, et al. Advances in reverse genetics-based vaccines of foot and mouth disease. Bing Du Xue Bao. 2014;30(2):213-20.
49. Fu Y, Cao Y, Sun P, Bao H, Bai X, Li P, et al. Development of a dot immunoblot method for differentiation of animals infected with foot-and-mouth disease virus from vaccinated animals using non-structural proteins expressed prokaryotically. J Virol Methods. 2011;171(1):234-40.
50. Uddowla S, Hollister J, Pacheco JM, Rodriguez LL, Rieder E. A safe foot-and-mouth disease vaccine platform with two negative markers for differentiating infected from vaccinated animals. J Virol. 2012;86(21):11675-85.
51. Palomares RA, Marley SM, Givens MD, Gallardo RA, Brock KV. Bovine viral diarrhea virus fetal persistent infection after immunization with a contaminated modified-live virus vaccine. Theriogenology. 2013;79(8):1184-95.
52. Alvarez M, Donate J, Makoschey B. Antibody responses against non-structural protein 3 of bovine viral diarrhoea virus in milk and serum samples from animals immunised with an inactivated vaccine. Vet J. 2012;191(3):371-6.
53. Oura CA, Edwards L, Batten CA. Evaluation of the humoral immune response in adult dairy cattle three years after vaccination with a bluetongue serotype 8 inactivated vaccine. Vaccine. 2012;30(2):112-5.
54. Calvo-Pinilla E, Castillo-Olivares J, Jabbar T, Ortego J, de la Poza F, Marin-Lopez A. Recombinant vaccines against bluetongue virus. Virus Res. 2014;182:78-86.
55. Biswas S, Bandyopadhyay S, Dimri U, Patra PH. Bovine herpesvirus-1 (BHV-1) - a re-emerging concern in livestock: a revisit to its biology, epidemiology, diagnosis, and prophylaxis. Vet Q. 2013;33(2):68-81.
56. Kumar P, Ayalew LE, Godson DL, Gaba A, Babiuk LA, Tikoo SK. Mucosal immunization of calves with recombinant bovine adenovirus-3 coexpressing truncated form of bovine herpesvirus-1 gD and bovine IL-6. Vaccine. 2014;32(26):3300-6.
57. Mackenzie-Dyck S, Kovacs-Nolan J, Snider M, Babiuk LA, van Drunen Littel-van den Hurk S. Inclusion of the bovine neutrophil Beta-defensin 3 with glycoprotein D of bovine herpesvirus 1 in a DNA vaccine modulates immune responses of mice and cattle. Clin Vaccine Immunol. 2014;21(4):463-77.
58. El-Kholy AA, Rady DI, Abdou ER, Elseafy MM, Abdelrahman KA, Soliman H. Construction, characterization and immunogenicity of a glycoprotein E negative bovine herpesvirus-1.1 Egyptian strain "Abu-Hammad". J Virol Methods. 2013;194(1-2):74-81.
59. Kortekaas J. One Health approach to Rift Valley fever vaccine development. Antiviral Res. 2014;106:24-32.
60. Wilson WC, Bawa B, Drolet BS, Lehiy C, Faburay B, Jasperson DC, et al. Evaluation of lamb and calf responses to Rift Valley fever MP-12 vaccination. Vet Microbiol. 2014;172(1-2):44-50.
61. Indran SV, Ikegami T. Novel approaches to develop Rift Valley fever vaccines. Front Cell Infect Microbiol. 2012;2:131.
62. Bird BH, Maartens LH, Campbell S, Erasmus BJ, Erickson BR, Dodd KA, et al. Rift Valley fever virus vaccine lacking the NSs and NSm genes is safe, nonteratogenic, and confers protection from viremia, pyrexia, and abortion following challenge in adult and pregnant sheep. J Virol. 2011;85(24):12901-9.
63. Lihoradova O, Ikegami T. Countermeasure development for Rift Valley fever: deletion, modification or targeting of major virulence factor. Future Virol. 2014;9(1):27-39.
64. Kortekaas J, Antonis AF, Kant J, Vloet RP, Vogel A, Oreshkova N, et al. Efficacy of three candidate Rift Valley fever vaccines in sheep. Vaccine. 2012;30(23):3423-9.
65. Weingartl HM, Nfon CK, Zhang S, Marszal P, Wilson WC, Morrill JC, et al. Efficacy of a recombinant Rift Valley fever virus MP-12 with NSm deletion as a vaccine candidate in sheep. Vaccine. 2014;32(20):2345-9.
Received in April, 2014.
Accepted in July, 2014.
Aníbal Domínguez. Dirección de Investigación y Desarrollo. Grupo Empresarial Labiofam. Avenida Independencia km 16 ½, Boyeros, La Habana, Cuba. E-mail: anibal.dominguez@labiofam.co.cu , anibaldominguezodio1@gmail.com.


{kind=link}
{kind=link}