










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.32 no.2 La Habana abr.-jun. 2015
RESEARCH
Lactic acid production by Lactobacillus plantarum L10 in a cell-retention continuous culture under different cultivation strategies
Producción de ácido láctico por Lactobacillus plantarum L10 en cultivo continuo con retención de células bajo diferentes estrategias de cultivo
Waldir D Estela-Escalante1,2, Karthik Kumaran AP3, Sourav Dutta3, Angélica Rodríguez-Best1
1 Facultad de Química e Ingeniería Química. Pabellón B. Universidad Nacional Mayor de San Marcos. Av. Universitaria s/n. Lima 1, Lima, Perú.
2 Institute of Chemical Technology Prague. Department of Fermentation Chemistry and Bioengineering. Faculty of Food and Biochemical Technology. Prague 6, Dejvice, Czech Republic.
3 Faculty of Engineering and Technology, SRM University, Kattankulathur, Chennai 603203, India.
ABSTRACT
The aim of this study was to evaluate the production of lactic acid in two different cell-retention continuous systems using Lactobacillus plantarum L10. This microorganism was cultured in synthetic medium under different cultivation strategies, temperatures (30 and 37 °C), medium composition and glucose concentration in the feeding medium (20, 40 and 80 g/L), at different dilution rates. Lactic acid’s yields and productivity were improved differently in both systems operating at different cultivation conditions. In one system name, 92.75 ± 0.18 % high yield values were obtained at low cultivation temperature, low dilution rates (0.21 h-1) and low glucose concentration in the feeding medium. In the other, a high productivity of 14.42 ± 0.08 g/L•h was attained, correlating to high dilution rates (0.48 h-1), cultivation temperature and glucose concentration in the feeding medium, respectively. Production of the L(+)-isomer of lactic acid was influenced by cultivation temperature while yeast extract concentration had a negligible effect. The results of this study can be used to optimize the production of L(+)-lactic acid in Lactobacillus plantarum cultures.
Keywords: continuous cultivation, cell-recycle system, Lactobacillus plantarum, L(+)-lactic acid.
RESUMEN
En este estudio se evaluó la producción de ácido láctico en dos sistemas de cultivo continuo con retención de células, mediante el cultivo de Lactobacillus plantarum L10 en medio sintético, con diferentes estrategias de cultivo. Esto incluyó diferentes temperaturas (30 y 37 °C), composición y concentración de glucosa en el medio de alimentación (20, 40 y 80 g/L), y tasas de dilución. La potenciación de los rendimientos y la productividad de ácido láctico en ambos sistemas se asociaron a distintas condiciones de cultivo y parámetros de operación. Un sistema mostró altos rendimientos del 92.75 ± 0.18 % a baja temperatura de cultivo, con bajas tasas de dilución (0.21 h-1) y baja concentración de glucosa en el medio de alimentación. El otro mostró una alta productividad de 14.42 ± 0.08 g/L•h que fue correlacionada con una mayor tasa de dilución (0.48 h-1), mayor temperatura de cultivo y mayor concentración de glucosa en el medio de alimentación. La producción del isómero L(+) de ácido láctico estuvo influenciada por la temperatura de cultivo, mientras que el efecto de la concentración de extracto de levadura fue insignificante. Los resultados de este estudio permiten optimizar la producción de L(+)-ácido láctico en cultivos de Lactobacillus plantarum.
Palabras clave: cultivo continuo, sistema de recirculación de células, Lactobacillus plantarum, L(+)-ácido láctico.
INTRODUCTION
Microbial production of lactic acid, particularly its L(+)-isomer, is of great interest from the industrial point of view for the synthesis of polymers with special chemical, physical, and biodegradable properties [1-3]. The production of the L(+) isomer depends mainly on the bacterial strain, growth temperature, pH, acid tolerance and substrate specificity [3-6].
Regarding the host system, lactic acid-producing bacteria can be further divided as homofermentative and heterofermentative microbes, the former having the greatest significance due to economic reasons [1, 2, 7]. Under laboratory and production scale conditions, homofermentative species normally yield lactic acid levels higher than 90 % their theoretical yields, by using glucose as substrate [3]. Growth of lactic acid-producing bacteria is optimal at a temperature and pH ranges of 10-45 °C and 3.5-9.6, respectively, but it also depends on the bacterial strain [8-10]. Nutritional requirements are also relevant, with a high demand for sources of complex nitrogen, amino acids, vita-mins (especially complex B), growth factors and trace elements [9]. In fact, this facilitates the use of cheap and undefined sources rich in these nutrients, in spite of synthetic media which is advantageous but more expensive [11].
The culture systems based on fed-batch and continuous cultivation are economically feasible for lactic acid production due to their high productivity, as compared with batch processes. Many studies cultivating free cell suspensions in continuous stirred-tank reactors (CSTRs) reported the increase in lactic acid productivity, the system used in combination with a cell-recycle system which incorporated a membrane separation unit to separate the product from cells [12-14]. Other CSTRs systems showed productivities ranging 8.9-13 g/L•h by using synthetic media, these values considerably higher than those obtained in batch processing (3-5 g/L•h). Additionally, the lactic acid yields were reported for both types of processes in the range 0.70-0.90 g with glucose as carbon source [12-16]. Other carbon sources such as whey permeate, lignocellulose-derived materials, cassava starch, among others, are cheaper but disadvantageous as compared with glucose, due to their low productivity values in the range 3-5 g/L•h [9, 11, 15, 17]. Hence, continuous cultivation with a cell-recycle system could be effective to improve the lactic acid yields and productivity. However, the control of the dilution rate is paramount since it can affect the cell physiology, biomass concentration and ultrafiltration membrane performance, its overall effect being detrimental for the productivity of the system [18].
Therefore, we report in this work the effect of cultivation conditions such as medium composition, temperature and dilution rates on the lactic acid yields and productivity, by comparing two different arrangements of a cell-retention continuous cell culture system for Lactobacillus plantarum L10 cultivation.
MATERIALS AND METHODS
Microorganism and its preservation
All the experiments were carried out using Lactobacillus plantarum L10 strain supplied by the Culture Collection of Dairy, Institute of Chemical Technology, Prague, Czech Republic. The strain was stored at –80 °C in a deep freezing chamber. Preservation was done in vials containing 2/3 volume of bacterial suspension in MRS medium (Oxoid, England) and 1/3 volumes of glycerol as cryoprotector.
Culture media and inoculum propagation
All reagents used for culture media preparation were ACS grade, and all media were adjusted to pH 5.8. Continuous cultivations were started in batch mode using a synthetic medium (80 g/L glucose, 13.3 g/L yeast extract, 5.3 g/L KH2PO4, 13.3 g/L CH3COONa, 5.3 g/L ammonium citrate and 0.5 g/L MgSO4•7H2O). The chemical composition of the feeding fresh media for continuous cultivations is showed in table 1. Three concentrations (w/v) of glucose as carbon source were used: 20, 40 and 80 g/L. Inoculum propagation was performed in static 100-mL cultures of sterile MRS medium in 500-mL Erlenmeyer flasks, either at 30 ºC for 18 h or 37 ºC for 14 h. The bioreactor containing 2 L culture medium was directly inoculated with 10 % (v/v) inoculum under aseptic conditions. Media sterilization was carried out in an autoclave at 0.1 Mpa. Small volume liquids were sterilized for 20 min and the larger ones (including the bioreactor containing the cultivation medium) and the accessories (nozzle, tubes, funnels, etc.) for 30 min.
Continuous cultivation experiments
Continuous cultivation experiments were carried out in a 2.5 L laboratory bioreactor (B. Braun Biotech International, Germany) containing 2 L of culture medium (as batch mode). Before autoclaving the culture medium, two drops of anti-foaming agent Breox (International Speciality Chemicals) was added, without any further addition of anti-foaming agents during cultivation. The bioreactor was connected to a micro-DCU-300 control and regulation unit. Bioreactor assembly included a mechanical stirrer (2 × 6 paddles), a stirring regulator MCU-200 and a module of peristaltic pumps (two for pH regulation and one for alternative use, either for antifoam or medium supply). pH value was controlled “on line“. Data collection was done by means of the software command, which was installed in a computer connected to the regulation control unit.
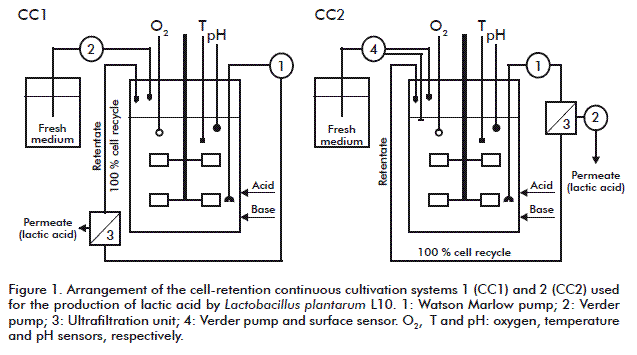
Cell-retention continuous cultivation systems
Two different arrangements of cell-retention continuous cultivation systems (CC) were used (Figure 1, CC1 and CC2, respectively). In each case, experiments started as batch cultivations (BC1 and BC2). Air was supplied only during the first 2 h of batch cultivation to promote cell growth (417 mL/min air flow rate). The stirring speed was 500 min-1. Cultivation temperatures were 30 °C for CC1 and 37 °C for CC2. Fresh feeding medium was pumped into the continuous cultivation system CC1 (Figure 1) with a peristaltic pump (Verder, USA). For a constant dilution rate, the outlet flow rate of the permeate had to equal the inlet flow rate of feeding fresh medium.
In the second arrangement, CC2 (Figure 1), the process was not managed by controlling the flow rate of the feeding fresh medium, but rather the permeate outflow rate. The permeate was pumped using a pump (Verner, USA) placed after the ultrafiltration unit at a defined flow rate. The bioreactor was installed internally with a surface probe, which was connected to a peristaltic pump.
The cell-recycle process in both systems was done by using peristaltic pumps (Watson Marlow, United Kingdom), silicone pipes and an ultrafiltration module developed and adapted from Melzoch et al. [19]. Medium was pumped to the ultrafiltration unit and the retained cells were flowed back into the bioreactor.
Analytical methods
Sampling
Samples for dry biomass determinations were taken by using a “T” device placed in front of the ultrafiltration unit. Those for determining the glucose and lactic acid concentrations were taken from the permeate (free cell fermented medium) leaving the filtration unit.
Dry biomass determination
Dry biomass was determined gravimetrically by using membrane filters (Millipore, type HA, pore size 0.45 μm). Five-milliliters sample volumes were prefiltered under reduced pressure in dried and weighed membrane filters, then, washed two times with 5 mL demineralized water and dried in an oven at 105 °C until constant weight was achieved. The filters were cooled down for 10 min in a desiccator and weighed. Dry biomass weight was calculated and expressed as g/L.
Glucose and lactic acid determination
Glucose and lactic acid concentration were determined by high-performance liquid chromatography (HPLC-laboratory instruments, Prague, Czech Republic). For separation of compounds, a column Ostion LGKS 0800 in H+ cycle was used. As mobile phase, a degassed 0.005 mol/L H2SO4 solution was used at a flow rate of 1 mL min-1. The column was tempered at 85 °C. Samples (20 μL) were injected through a three-way dispensing valve Rheodyne. The column output was detected by a refractive index detector RIDK 101 connected to a computer, and data were analyzed with the Data Apex program. Glucose and lactic acid concentrations were determined by using the external standard method.
Enzyme assay of lactic acid isomers
Determination of lactic acid isomers was based on the oxidation of D-lactic acid (L-lactic acid) to pyruvate in the presence of D- and L-lactate dehydrogenase using Nicotinamide adenine dinucleotide oxidized (NAD+). The amount of reduced nicotinamidadenindinucleotide (NADH) formed corresponded to the amount of D-lactate and L-lactate. The increase of NADH concentration was determined by measuring the absorbance at 340 nm. The amounts of D- and L-lactic acid in the cuvette must have to be in the range of 0.3 to 20 μg. Absorbance was measured on the spectrophotometer (Shimadzu, Japan).
The substrate concentration (S) during continuous cultivation (C), and the productivity (Pp) and yield (Yp/s) of lactic acid in pseudosteady state of the continuous system were calculated by the following formula:

Where: Si and So were the substrate concentrations in the fresh medium inlet or the permeate outlet, respectively (g/L).

Where:
D: Dilution rate (h-1).
PU: Lactic acid concentration in pseudosteady state (g/L).

Where SU: Substrate concentration in pseudosteady state (g/L).
RESULTS AND DISCUSSION
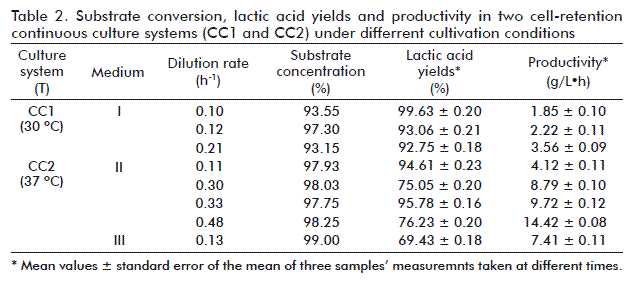
The arrangement of both cell-recycle continuous cultivation systems are shown in figure 1. In continuous cultivation system CC1, fresh medium with glucose concentration of 20 g/L (medium I) was used in the feeding. In this case, the increase of dilution rate did not cause the increase of substrate concentration in the bioreactor and substrate levels were constant at very low values. In the CC2 system, medium II (glucose concentration of 40 g/L) was used and substituted by medium III after a 69-h cultivation. The parameters and results of cultivation experiments with both CC systems are summarized in table 2. Working at low dilution rates in both CC1 and CC2 systems, high lactic acid yields can be obtained regardless the glucose concentration in the feeding medium, but affecting productivity values. Additionally, working at similar dilution rates (0.10 h-1 for CC1 and 0.11 h-1 for CC2); the temperature, medium composition and the arrangement of the cultivation system exert an important effect on the yield and productivity values of lactic acid. From the economical point of view, a low cost process with balanced yields and productivity is preferred.
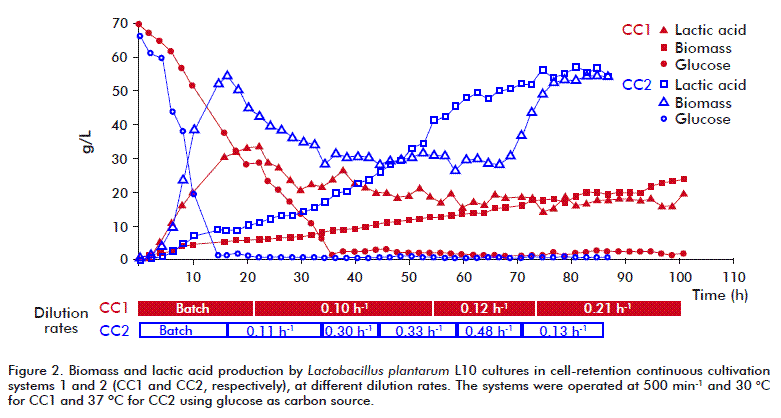
In the CC1 system (Figure 2), the fresh medium I feeding began after 22 h, and after 16 h in CC2. In this experiment, initial glucose and biomass concentrations in the medium after batch cultivation were approximately 10 g/L and 5 g/L, respectively. Very low residual glucose concentrations were detected in the medium by working at a dilution rate of 0.1 h-1 (pseudosteady condition), this parameter increased until 0.21 h-1. The dry biomass weight was 23.06 g/L at the end of the CC1 process after 101 h.
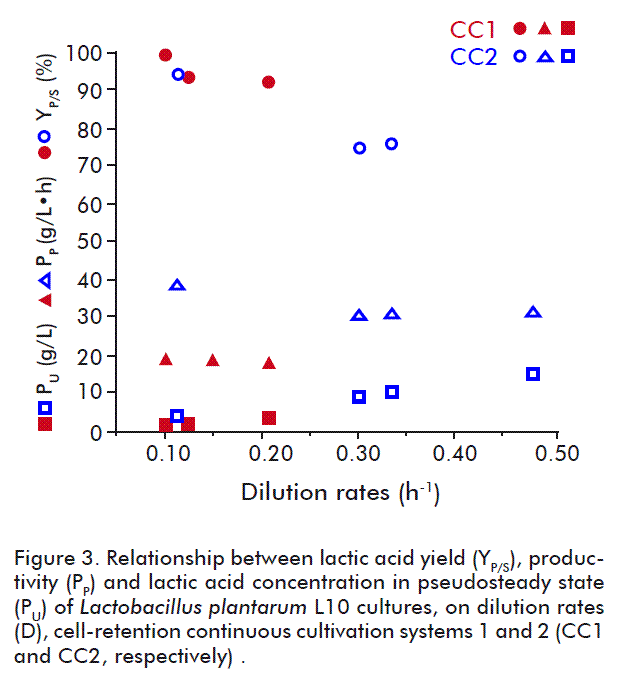
The influence of the dilution rate on the parameters evaluated is shown in Figure 3. Remarkably, the increase of the dilution rate caused an increase in productivity. The lactic acid productivity is a crucial parameter to be considered, but in terms of isolation and purification it is preferable to choose the higher lactic acid yield instead of the higher productivity of the system.
The feeding of fresh medium in the CC2 system (Figure 2) was started when the glucose and dry biomass concentrations in the medium were approximately 1.47 g/L and 9 g/L, respectively (after batch cultivation BCII). The fresh medium II (4 % w/v glucose concentration) was used in the feeding. In this experiment, working even at the highest dilution rates, the residual glucose concentration did not exceed 1 g/L. Here, the rise in the dilution rate increased the productivity of the system, but at almost constant yields and lactic acid concentrations in the permeate, even at dilution rate values higher than 0.3 h-1 (Figure 3). This phenomenon can be explained by the ever-increasing concentration of cells in the system. High cell concentrations in the medium are explained by the lack of effect of the increased dilution rate on residual glucose concentrations in the medium under pseudosteady state. At the end of the CC2 process (86 h), the dry biomass concentration in the medium reached 54.76 g/L.
In the CC1 system, the increase of the dilution rate slightly decreased lactic acid yields, while in the CC2 system its increase from 0.11 to 0.30 h-1 caused a drastic fall in lactic acid yields, at levels that remained significantly unchanged, even after increasing the dilution rate. This phenomenon is caused by the poor concentration of yeast extract in the feeding medium II used in the CC2 system as compared to its glucose ratio. A balanced concentration of nitrogen and carbon source in the medium is very important for production of lactic acid as reported by Estela et al. [16]. At the end of the experiment, the change of dilution rate from 0.48 to 0.13 h-1 was related to the replacement of fresh medium II by fresh medium III. Thus, glucose concentration in the medium increased but the rest of components remained equal. As a consequence of this change, a further decline of lactic acid yields to 69.43 ± 18 % was observed, the formation of lactic acid limited by the lack of nitrogen sources because of it use in biomass production.
On the contrary, the limitation of lactic acid formation due to the lack of nitrogen sources was not observed during the CC1 process, with yields above 92.75 ± 0.18 %. This was explained by the yeast extract/glucose concentration ratio in the feeding medium I, two- and four-fold as compared with that of feeding media II and III, respectively. From these results it can be concluded that even higher lactic acid yield and productivity values could be achieved in the case of CC, by using feeding media with higher yeast extract content and under similar conditions as used in the
CC2 system. The best results of lactic acid yield were obtained in the CC1 system at 0.10 h-1 dilution rate (99.63 ± 0.20 %). In terms of lactic acid productivity, the best results were obtained in the CC2 system at a dilution rate of 0.48 h-1 (14.42 ± 0.08 g/L• h).
The productivity results obtained in the CC2 cultivation system (D = 0.30 h-1; table 2) were two-fold those obtained by Bomrungnok et al. [17]. They attained a productivity of 4.53 g/L•h in continuous cultivation (D = 0.4 h-1) without a cell-recycle system, by using a semi synthetic medium containing cassava starch as carbon source and by cultivating Lactobacillus plantarum SW14 at 30 °C.
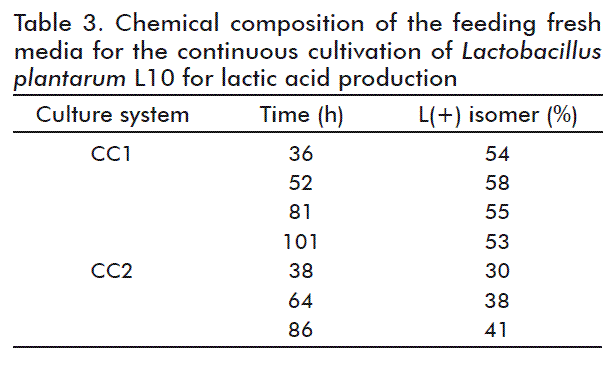
The L(+)-isomer of lactic acid concentrations obtained in the product are summarized in table 3, this parameter significantly influenced by the temperature. Cultivation at 37 ºC caused the reduction of L(+)-isomer, while a racemic mixture was obtained at 30 ºC. Otherwise, the dilution rate had no clear effect on the fraction of the lactic acid L(+)-isomer.
As previously reported [20], yeast extract concentrations higher than 1 % (w/v) reduces the optical purity of the product, because it contains a certain amount of lactic acid [20]. In our CC experiments, the concentration of yeast extract in the medium was only 0.5 % (w/v), but in the case of batch cultivation was 1.33 % (w/v), indicative of higher demands for nitrogen compounds at the initial stages of growth and production. For this reason, we determined the total concentration of lactic acid (0.02 g/L) and L(+)-isomer in the yeast extract solution (13.3 g/L in water). The results indicated that the influence of yeast extract (Oxoid, United Kingdom) on the optical purity of lactic acid was negligible.
Regarding the effect of temperature on the L(+)/D(-)-isomer ratio of lactic acid in the product at the end of the experiment, a racemic mixture was obtained at 30 ºC in batch cultivations, this parameter drooping at 37 °C. Nevertheless, there was a slightly higher increase of the L(+)-isomer content from batch to continuous cultivation systems.
CONCLUSIONS
The production of lactic acid was shown to be influenced by several cultivation conditions such as: medium composition (mainly nitrogen source availability), cultivation temperature and the dilution rate. In the CC1 system, working at low dilution rates (0.10 h-1), 30 °C and a low glucose concentration medium (20 g/L), the highest lactic acid yield was obtained. Meanwhile, the highest productivity was attained in the CC2 system, working at 37 °C, a higher dilution rate (48 h-1) and using a feeding medium at 40 g/L glucose. Regarding the balance of L(+) and D(–)- lactic acid isomers in the product, only the cultivation temperature showed a significant effect, since lower cultivation temperature favored L(+)-lactic acid formation. On the other hand, yeast extract showed a negligible effect on the optical purity of lactic acid.
These results could support the combination of the cell-retention CC systems, based on the factors detected as influencing each of the lactic acid production parameters, for increased yields and productivity.
REFERENCES
1. Gao C, Ma C, Xu P. Biotechnological routes based on lactic acid production from biomass. Biotechnol Adv. 2011;29(6):930-9.
2. Okano K, Tanaka T, Ogino C, Fukuda H, Kondo A. Biotechnological production of enantiomeric pure lactic acid from renewable resources: recent achievements, perspectives, and limits. Appl Microbiol Biotechnol. 2010;85(3):413-23.
3. Abdel-Rahman MA, Tashiro Y, Sonomoto K. Recent advances in lactic acid production by microbial fermentation processes. Biotechnol Adv. 2013;31(6):877-902.
4. Cui F, Li Y, Wan C. Lactic acid production from corn stover using mixed cultures of Lactobacillus rhamnosus and Lactobacillus brevis. Bioresour Technol. 2011;102(2):1831-6.
5. Tashiro Y, Kaneko W, Sun Y, Shibata K, Inokuma K, Zendo T, et al. Continuous D-lactic acid production by a novel thermotolerant Lactobacillus delbrueckii subsp. lactis QU 41. Appl Microbiol Biotechnol. 2011;89(6):1741-50.
6. Castillo-Martinez FA, Balciunas EM, Salgado JM, Domínguez-González JM, Converti A, Pinheiro de Souza Oliveira R. Lactic acid properties, applications and production: A review. Trends Food Sci. Technol. 2013;30(1):70-83.
7. Jem K, van der Pol J, de Vos S. Microbial lactic acid, its polymer poly(lactic acid) and their industrial applications. In: Chen, GGQ, editor. Plastics from bacteria. Matural functions and applications. Berlin: Springer; 2010. p. 323-46.
8. Akerberg C, Hofvendahl K, Zacchi G, Hahn-Hagerdal B. Modelling the influence of pH, temperature, glucose and lactic acid concentrations on the kinetics of lactic acid production by Lactococcus lactis ssp. lactis ATCC 19435 in whole-wheat flour. Appl Microbiol Biotechnol. 1998;49:682-90.
9. Hofvendahl K, Hahn-Hagerdal B. Factors affecting the fermentative lactic acid production from renewable resources(1). Enzyme Microb Technol. 2000;26(2- 4):87-107.
10. Hetényi K, Németh A, Sevella B. Role of pH-regulation in lactic acid fermentation: Second steps in a process improvement. Chem Eng Process. 2011;50(3):293-9.
11. Bustos G, Moldes AB, Cruz JM, Dominguez JM. Formulation of low-cost fermentative media for lactic acid production with Lactobacillus rhamnosus using vinification lees as nutrients. J Agric Food Chem. 2004;52(4):801-8.
12. Wee YJ, Ryu HW. Lactic acid production by Lactobacillus sp. RKY2 in a cell-recycle continuous fermentation using lignocellulosic hydrolyzates as inexpensive raw materials. Bioresour Technol. 2009;100(18):4262-70.
13. Dey P, Pal P. Direct production of L(+) lactic acid in a continuous and fully membrane-integrated hybrid reactor system under non-neutralizing conditions. J Membr Sci. 2012;389(1):355-62.
14. Pal P, Dey P. Process intensification in lactic acid production by three stage membrane integrated hybrid reactor system. Chem Eng Process. 2013;64:1-9.
15. Abdel-Rahman MA, Tashiro Y, Sonomoto K. Lactic acid production from lignocellulose-derived sugars using lactic acid bacteria: overview and limits. J Biotechnol. 2010;156(4):286-301.
16. Estela W, Rychtera M, Melzoch K, Quillama E, Egoavil E. Production of lactic acid by Lactobacillus plantarum L10 in batch and continuous cultivation. Rev Peru Biol. 2007;14(2):271-5.
17. Bomrungnok W, Sonomoto K, Pinitglang S, Wongwicharn A. Single step lactic acid production from cassava starch by Lactobacillus plantarum SW14 in conventional continuous and continuous with high cell density. APCBEE Procedia. 2012;2:97-103.
18. Kulozik U. Physiological aspects of continuous lactic acid fermentations at high dilution rates. Appl Microbiol Biotechnol. 1998;49(5):506-10.
19. Melzoch K, Votruba J, Habova V, Rychtera M. Lactic acid production in a cell retention continuous culture using lignocellulosic hydrolysate as a substrate. J Biotechnol. 1997;56(1):25-31.
20. Bruno-Barcena JM, Ragout AL, Cordoba PR, Sineriz F. Continuous production of L(+)-lactic acid by Lactobacillus casei in two-stage systems. Appl Microbiol Biotechnol. 1999;51(3):316-24.
Received in April, 2014.
Accepted in July, 2015.
Waldir D Estela-Escalante. Facultad de Química e Ingeniería Química. Pabellón B. Universidad Nacional Mayor de San Marcos. Av. Universitaria s/n. Lima 1, Lima, Perú.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}