










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.32 no.4 La Habana oct.-dic. 2015
RESEARCH
Protein enrichement of fruit processing byproducts using solid state fermentation with Saccharomyces cerevisiae and Bacillus subtilis
Enriquecimiento proteico de subproductos frutícolas mediante fermentación en estado sólido con Saccharomyces cerevisiae y Bacillus subtilis
Karina M Rompato, Susana N Somoza
Laboratorio de Investigación en Alimentos, Facultad de Ciencias de la Salud, Universidad Nacional de Formosa. Av. Gutnisky 3200, Formosa, CP 3600, Argentina.
ABSTRACT
The byproducts of the agricultural industry are useful substrates for protein enrichment using solid state fermentation (SSF). Unicellular protein production is characterized by its high nutritional quality and Saccharomyces cerevisiae is the world’s most widely produced microorganism. Grapefruit bagasse and banana peels are residues from the food processing industry, the pods of the mesquite are not used for human feeding; they are found in large amounts, rich in sugars and suited for SSF with S. cerevisiae and Bacillus subtilis for the production of microbial protein. The study started with the assumption that a strain of B. subtilis, an efficient producer of endoglucanases, together with S. cerevisiae, may improve the production of protein through their hydrolytic action on the polysaccharides of the substrates. The aim was to assess the microbial protein enrichment using fruit byproducts in SSF according to the microorganisms, substrates and incubation time periods used. The experimental data confirm that it is possible to transform industrial waste into products of high nutritional value. The protein value of grapefruit bagasse and banana peels byproducts was increased in more than 150 and 115 %, respectively, and in 80 % for the mesquite pods, which reached the highest protein concentration of 23.6 g/100 g of byproduct.
Keywords: solid state fermentation, protein enrichment, single cell protein, Saccharomyces cerevisiae, Bacillus subtilis.
RESUMEN
Los subproductos agroindustriales constituyen sustratos útiles para el enriquecimiento proteico mediante fermentación en estado sólido (FES). La producción de proteína unicelular se caracteriza por su alta calidad nutricional y Saccharomyces cerevisiae es el microorganismo de mayor producción mundial. El bagazo de pomelo y las cáscaras de banana constituyen residuos agroindustriales, las vainas de vinal no se aprovechan para la alimentación humana, son matrices abundantes, ricas en azúcares y adecuadas para la FES con S. cerevisiae y Bacillus subtilis para la producción de proteína microbiana. Se parte del supuesto que una cepa de B. subtilis, eficiente productora de endoglucanasas, en consorcio con S. cerevisiae, podría mejorar la producción de proteínas debido a su acción hidrolítica sobre los polisacáridos de los sustratos. Este trabajo tuvo como objetivo evaluar el enriquecimiento proteico microbiano a partir de subproductos frutícolas en FES según microorganismos utilizados, sustratos, y tiempo de incubación. Los datos experimentales permiten afirmar que es posible transformar desechos industriales en producto de alto valor nutricional. El valor proteico de estos subproductos se incrementó en más de 150 %, para el bagazo de pomelo, en 115 % el de las cáscaras de banana y en 80 % las vainas de vinal, siendo éste último el sustrato con el que se alcanzó la mayor concentración de proteínas, llegando un máximo de 23.6 g/100 g.
Palabras clave: fermentación en estado sólido, enriquecimiento proteico, proteína de célula única, Saccharomyces cerevisiae, Bacillus subtilis.
INTRODUCTION
The process for microbial production basically requires a substrate in a medium under the appropriate conditions that would enable the multiplication of the selected microorganism. This process may have the purpose of obtaining the microorganisms per se, or through the application of a biotechnological method, that of obtaining certain byproducts such as proteins or other biomolecules.
Scientific literature refers to the unicellular protein using the English term ‘single cell protein’ (SCP), which is that obtained from the microbial biomass of algae, bacteria, yeast or filamentous fungi, grown under fermentative conditions that will ensure an appropriate growth rate, using inexpensive substrates composed by, or enriched with, carbon, nitrogen and phosphorus. It also covers dead and dry microorganisms used in animal feeding, without the mediation of protein extraction or purification processes [1], and the obtained SCP is characterized by its high nutritional quality and because it does not contain, or generate during processing, any toxic or potentially carcinogenic compound [2].
Generally, SCP is produced through a process called submerged fermentation, where the microorganism most widely produced in the world is Saccharomyces cerevisiae [3], and where three aspects are key components for the outcome: the microorganism, the substrate and the production process involved.
The production processes using alternative substrates, such as waste from the agricultural industry, require, in most cases, a prior constituent polysaccharide hydrolysis stage, generally using a strong acid, such as HCl, neutralization, sterilization and fermentation. Examples of this are the studies using cucumber and orange peel residues as substrates [4]; as well as potato, carrot, orange and apple peels, [5] the peels of banana [6], malt extract, apple, molasses and milk whey with four different yeasts [7]. Particularly, solid state fermentation (SSF) is used, this process involving the growth of microorganisms in humid solid materials [8]. The protein produced through SSF generally use substrates that are byproducts from the food industry, as for example sugarcane bagasse [9], and citrus fruit bagasse [10]. The advantages lie mainly in saving the energy used to dry the fermented material, store it and transfer it to its final destination. The use of residues from the food industry in solid state fermentation processes has the potential of obtaining byproducts such as organic acids, enzymes, aromatic compounds, fruit oligosaccharides, as well as bioactive agents such as gibberellic acid, antibiotics, bioinsecticides and bioethanol, for which a series of microorganisms, mainly of fungi and those of bacterial nature, have been evaluated [11].
Other studies assess the fermentation conditions for protein production with different microorganisms, for example Candida MCCF 101 of marine origin for its use as feed for shrimp [12].
Regarding the microorganism used, its waste conversion capacity resides on its capability to secrete enzymes able to digest the byproducts used for SCP production, particularly glucanases. These are enzymes that degrade β-glucans and are classified into two large groups according to the mechanism used to hydrolyze the substrate, identified by the hydrolyses products: i) the exo-β-glucanases, which hydrolyze the substrate through the sequential rupture of glucose residues from the non-reducing end, and ii) the endo-β-glucanases that randomly break the β binding sites of the polysaccharide chain, releasing small oligosaccharides [13]. The cellulases, which are cellulose-degrading enzymes, are a mixture of hydrolytic enzymes that include endoglucanases (EGs), and cellobiohydrolases (CBHs). Since the cellobiose and cellodextrans are inhibitors of the CBHs, the presence of β-glucosidase is required for the complete hydrolysis of the cellulose [14]. This clarification is valid, from the point of view of the use given to both the terms, cellulases and glucanases, in the scientific literature where they are sometimes used as synonyms.
Several authors have used cellulase strains producing Bacillus to improve the nutritional quality of leaf meal. Bairagi et al. [15] demonstrated that it is possible to incorporate up to 40 % of Leucaena leaf meal inoculated with fish intestinal bacteria that produce enzymes for the rations of the fish Labeo rohita Hamilton. After 15 days of fermentation, the leucaena leaf meal showed a significant increase in the level of free amino acids and fatty acids; there was a decrease in crude fiber, cellulose, hemi-cellulose and anti-nutritional factors such as tannins, phytic acid and mimosine [15].
In the case of the substrate for SCP production, bibliography regarding fermentative processes of legumes such as soybean in the preparation of foods is widespread; these products are mainly originated and consumed in oriental countries, as for example Kinenma, Natto, Thua-Nao, Dawadawa using B. subtilis, which are traditionally used in Asian and African cultures [16, 17]. The fruits of Prosopis sp., largely found in America and Africa have hardly been studied as raw material for fermentation. Evans et al. studied the benefits of the fermented foods of Nigeria, some of which include seeds of Prosopis sp. [18]. The mesquite (visnal, viñal, olkhá, pao de espinho, quilín, tayt and yuncumarim) having the scientific name Prosopis ruscifolia, grows in Formosa Province. This species is less known and used compared to others of the same genus. It is currently estimated that this species is found in more than 2 million hectares of the Republic of Argentina, with large amounts growing in the Province of Formosa [19]. The records of the use of its fruits in fermentative processes are scarce.
Grapefruit bagasse is constituted by the residues remaining in the fruit after the juice is obtained, in other words, the peel, pulp and seeds. Formosa has white grapefruit with the highest level of sugar in all of Argentina, and therefore the companies at the southern part of the country use it to mix it with their own juices, so that it may respond to the taste of the most demanding European consumers [20]. At present, the main problem of the citrus fruits enterprises are the effluents having high organic loads, some of which are dumped into the canals that pour their waters into the basin.
Bananas are tropical fruits also used for the process mentioned, belonging to the genus Musa cavendishii, which are produced commercially or for their own consumption. Formosa produces between 40 and 50 thousand tons of bananas, representing from 40 to 50 percent of the country’s production. In contrast to the Ecuadoreans, the bananas produced in Formosa contain more sugar [21]. These crops generally yield a large amount of residues and excess fruit that are not suitable for human consumption. Noteworthy, reports referring to the nutritional composition of banana peel are scarce. Brazilian researchers tested its use as a fiber supplier in crackers made with banana peel meal. They determined the total neutral detergent fiber content (NDF) as being 32 g/100 g and proteins were at the level of 8.6 g/100 g [22].
Therefore, this work was aimed to assessing the microbial protein enrichment of fruit byproducts (grapefruit bagasse, banana peels and mesquite pods) under SSF in relation to the microorganisms, substrates and incubation time used. The main purpose is to enrich the rations based on local products with high biological value (microbial) SCP proteins.
MATERIALS AND METHODS
Plant substrates
Grapefruit bagasse consisting of the pulp and fruit peel was obtained from the local juice extracting industry. After eliminating the seeds, it was place at 46 °C in a dehydrator for 24 and 48 h; then it was ground in a blade mill and stored in glass flasks in a dry dark place for later use. The banana peels were obtained from local fruits purchased at the market; the same procedure used for grapefruit bagasse was followed. The mesquite was obtained by collecting the pods at the Departments of Matacos and Ramón Lista in the western part of the Formosa Province, Argentina. They were dried in an oven at 60 °C for 24 h, and then were ground in a blade mill and separated from the non-reducible woody material with a 1-mm sieve; they were then put away in glass flasks in the dark for later use.
Microorganisms
The S. cerevisiae yeast was obtained at the local market in a dry form (Levex, Argentina); it was dissolved in sterile saline solution and isolated with the use of a solid YM medium (yeast and mushrooms; Britania, Argentina) to obtain pure colonies. They were stored in a liquid Saboureaud medium under refrigeration. Before its use, the strain was activated by seeding it in a Saboureaud culture at 37 °C for 24 h.
From the aquariums where juvenile pacu fish are raised, several glucanase-producing Bacillus strains were isolated; among these we selected strain 3 because it had the best glucanase activity with carboxymethylcellulose (CMC), and was later identified as B. subtilis. It was stored in Brain Heart Infusion (BHI) medium under refrigeration. At 24 h before starting the experiment, it was cultured in a liquid medium containing 1 % CMC, 1 % yeast extract and 1 % peptone.
SSF
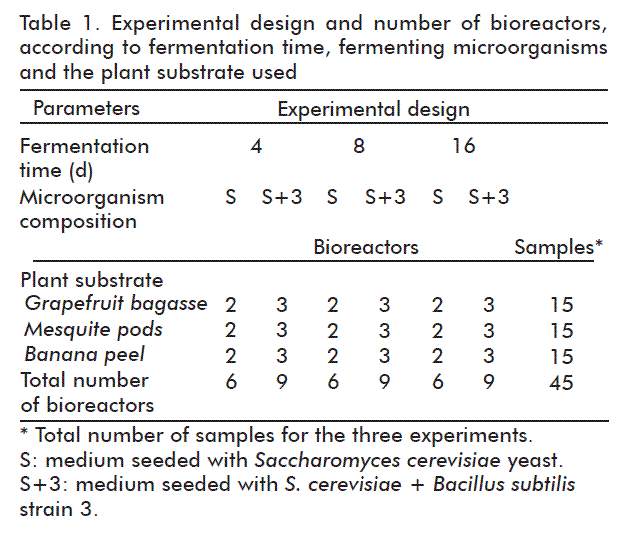
Glass bioreactors with a capacity of 500 mL having metal lids were used to carry out the fermentation. In each reactor, 20 g of the dry plant material were placed with 100 mL of a buffer consisting of 1 g KH2PO4, 1 g K2HPO4, 0.4 g MgSO4·7H2O, 5 g (NH4)2SO4, 0.05 g CaCl2 · H2O, and 0.0125 g FeSO4·7H2O, in 1000 mL of deionized water. The pH was adjusted to 5.0 only in the samples containing grapefruit bagasse, by adding 0.1 N NaOH, the rest had pH values above this level in the range 5.5-6.0. Substrates were sterilized in an autoclave at 121 °C for 15 min, left to cool and were seeded with the microorganisms, 1 mL of the yeast inoculum and 2 mL of the bacterial inoculum, both were grown for 24 h in the above described culture media, and they were cultured in an oven at 37 °C for 4, 8 and 16 days with the lids partially unscrewed so that oxygen could enter the bioreactors. As shown in Table 1, three assays using different time periods and lots were carried out for each plant matrix. Only the combination of yeast and bacteria S+3 (S. cerevisiae + B. subtilis strain 3) was seeded in the first assay, where we used 3 bioreactors. The other two assays consisted of the preparation and incubation of 6 bioreactors, 3 seeded only with yeast (S. cerevisiae), and 3 seeded with the S+3 combination. At 4 days, two bioreactors were retrieved per matrix, one S and one S+3, repeating the procedure on days 8 and 16.
Every other day the bioreactors were shaken manually to homogenize the material and favor the contact with the microorganisms. At the same time, the signs of deterioration, such as an unpleasant odor, or the presence of moss colonies, would imply the elimination of the corresponding bioreactors. After the fermentation period, the bioreactors were opened and the organoleptic properties, aroma, color and consistency of the process materials were assessed. Later they were dried in dehydrators for 24 to 48 h. After drying they were again ground and put away in airtight polyethylene bags placed in boxes to protect them from the light and humidity until they were processed.
Analytical determinations
Total protein, total soluble sugars, color and consistency were determined in all materials before and after fermentation as response variables. The bioreactors showing signs of contamination and deterioration were discarded.
The proteins were determined by the Kjeldahl method [23] and total sugars were determined by the Nelson-Somogyi method [24]. Glucose was determined in the grapefruit samples by the glucose-oxidase method [25](reagent from Wiener Lab, Argentina). The seeded inocula were counted by the plate counting method, using the YM culture medium for the yeasts and the BHI (Britania, Argentina) for the bacteria. Colonies counts were expressed as colony forming units per milliliter (c.f.u./mL) and transformed into logarithms for their statistical analysis.
Data processing and statistical analysis
The pre and post fermentation differences were calculated with the data of final protein, and the productivity indicators Yx/s were determined with the soluble sugars. This index is usually calculated with the weight of the biomass, x, obtained in relation to the concentration of the substrate, s; however, the index Yp/s can also be used, where p is the fermentation product that will be measured, which is in this case, protein concentration. The variation in protein concentration is attributed to the microbial protein. The productivity index (PI) was calculated using the (microbial) protein produced, and the total initial sugars (p and s, respectively).
Protein and sugars were determined in duplicates in each sample (45 samples) and an average value was used. A Student’s t test was carried out for mean comparison in order to evaluate the biological variability between the samples in the same substrate matrix. The data, obtained in the different fermentation periods and microorganisms used, were statistically processed using the free version InfoStat program [26] for the analysis of variance (ANOVA), and means were compared by the Fisher’s test. A 2×4 block factorial design was applied, using time period as the treatment with four levels (1, 4, 8 and 16 days) and the microbial composition with two levels: S and S+3. This design made it possible to study two factors simultaneously, as well as their combined effects on the response variable. In this way, the inference base was increased in relation to each factor, and the degree and form of the effect of time through the interaction was determined by using two different microbial compositions, on the amount of proteins produced under the experimental conditions established.
RESULTS
Initial sugars and proteins on each individual substrate matrices were analyzed by the Student’s t test, but no significant differences were found in the parameters analyzed in regard to the samples of the different assays, thereby assuming that the initial conditions were similar. The three matrices were processed in three different assays: one corresponding to the three repeats of the yeast/bacteria combination, and two of them corresponding to the two repetitions yeast fermentation alone were tested: Only the combination of yeast/bacterium (S+3) was studied in assay I; the test with yeast alone (S) was not made.
The data analyses in each matrix was made using a 2×4 block factorial design employing as classification variables, the microbial composition (S and S+3) and the time in its initial and fermentation levels at 4, 8 and 16 days. Below we present the results in relation to the matrices assessed separately.
Grapefruit bagasse
No fermentation was discarded due to alterations in the organoleptic properties; all of them had very pleasant aromas and none of them was contaminated. The solid product obtained after processing was clear and had a citrus fruit aroma.
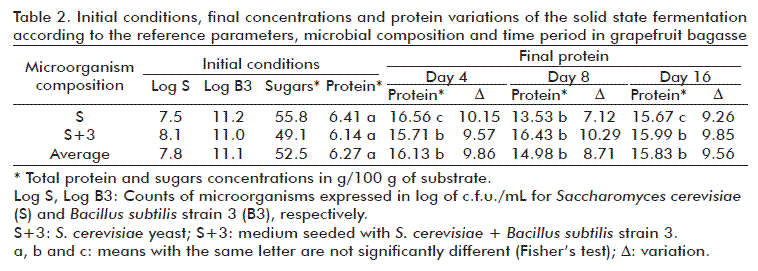
The initial samples contained 5.87, 7.78 and 8.41 g/ 100 g of protein and 35.68, 51.89 and 59.80 g/ 100 g of total soluble sugars in each assay. Table 2, shows the average data of the initial composition, the concentration of protein and sugars, the amount of microorganisms seeded in log of c.f.u./mL, and the results of final proteins at 4, 8 and 16 days of fermentation; also the difference in protein content compared to the initial value, which corresponds to microbial protein.
The analysis of variance determined that the final protein concentration of the bioreactors containing only the yeast shows that there is a statistically significant difference between the initial values and the time period of the assay. Considering the number of days of the trial for level S, there is a statistically significant difference (p < 0.05) between day 4, having the highest protein concentration, and day 8. For the level S+3 the results show significant differences (p < 0.05) between the means of the initial protein and the days tested separately (for each trial).
No statistically significant differences were found in the protein concentrations obtained using S and S+3, in the complete experiment.
In the bioreactors containing grapefruit bagasse meal as the substrate and only S. cerevisiae as the microorganism, the maximum total protein was obtained on day 4, 16.6 g, corresponding to a production of 10.2 g of microbial protein per 100 g of the substrate. In the bioreactors containing S. cerevisiae and B. subtilis, the protein average of the experiment was of 16 g of total protein during the time periods of the trial (without any significant differences between them). In other words, 9.8 g of microbial protein were produced.
The data on total proteins obtained on day 4 (the day of the highest protein production) in the trials containing only yeast (S) did not differ significantly from those obtained in the bioreactors with S+3.
For the trials with yeast (S), the increase in protein is of 156 % and for the trials with the combination of yeast/bacteria (S/S+3), an increase of 157.3 % of protein was obtained, as shown in Table 3.
The PIs calculated using the data obtained from the microbial protein (on day 4) and the initial sugar content were: 0.18 for the bioreactors containing yeast (S) and 0.21 for those containing the combination S+3.
Total residual sugars were also evaluated; we found means in g/100 g of the sample, of 3.99 at 4 days, 5.93 at 8 days and 4.58 at 16 days. In relation to the initial reducing sugars, corresponding to glucose, it was at the range of 4.8 and 8 g/100 g of the sample.
Banana peel
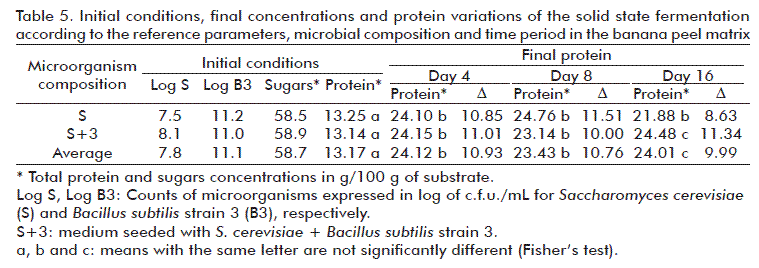
It was not necessary to discard any of the fermentations since all of them had pleasant odors and none of them showed any sign of contamination. After processing, the solid material obtained was dark and had a pleasant banana odor. The mean values of each assay of the initial parameters of protein concentration, sugars, and microorganism count in the inocula are shown in the initial conditions and in the results of the process, which are summarized in Table 4.
Using the ANOVA we determined that there were significant differences (p < 0.05) between the initial protein concentration and the number of days of the fermentation period in both bioreactor groups. In the same way as for the grapefruit bagasse meal substrate using a 2×4 block factorial design, we determined that there were significant differences between the protein concentrations of days 4 and 8 and between days 8 and 16 for the bioreactors S (only yeast), where the highest value was on day 4. However, in the bioreactors containing S+3 there was a statistically significant difference between the protein concentrations obtained on day 16, compared to days 4 and 8. The highest protein concentration for level S was obtained on day 4, and it was of 14.8 g/100 g, and the highest mean of the S+3 was obtained on day 16 and it was of 15.50 g/100 g. This corresponds to an increase of 115 % of protein in the bioreactors containing yeast (S) and of 98 % in the bioreactors containing the combination S+3.
The pPIs calculated using the microbial protein data obtained (on the day of the maximum concentration for each level) and the initial sugar content, were of 0.166 for the bioreactors containing yeast (S) and 0.172 for those containing the combination (S+3). The residual sugars were at a range of 3.6 to 5 g/100 g.
Mesquite pods
This plant matrix showed the highest amount of soluble sugars, 50.59, 50.81 and 66 g/100 g of the sample. In relation to the assessment of the organoleptic properties of the bioreactors, two bioreactors from the second assay were discarded because they showed signs of contamination and unpleasant odors. Both bioreactors were seeded with yeast alone. The rest had a vanilla aroma, which is characteristic of that fruit.
Table 5 describes the initial composition of the reaction and the results of the protein concentrations obtained in the bioreactors with mesquite pods.
The analyses of the averages of protein concentration, through the block factorial design, resulted in a statistically significant difference (p < 0.05) between the initial values and the intervals tested separately, but not between days 4, 8 and 16 (in other words, there are no statistically significant differences between the protein concentrations obtained on day 4 and those obtained on day 8, or those obtained on day 8 with those of day 16). Therefore, the mean value of the protein concentration for the periods of the trial at the S level was of 23.58 g/100 g and at S+3 it was of 23.92 g/100 g.
In this case, the calculation of the productivity indices was made taking into account the means of the microbial protein on the days tested and the initial sugar content of each trial; for the bioreactors containing the yeast (S) it was of 0.182 and 0.179 for those containing the combination. In relation to the residual sugars, the maximum value found was of 5.63 g/100 g of residual sugars.
Analyzing the statistical design in a global manner, it was found that the level of significance attained with the block factorial design reached, in all cases, a value of p < 0.05, for the model and the time period factor. This enables us to reject the null hypothesis for all trials and matrices; in other words, a statistically significant difference was produced between the initial and final protein concentrations in all the substrates in their respective levels of microbial constitution. However, it was not possible to reject the null hypothesis for the ‘microbial composition’ treatment, in any of the substrates analyzed.
In order to evaluate if the protein concentrations obtained according to the substrates analyzed are significantly different, as well as the possible interactions among the treatments, a three factor design was proposed, considering substrate, time period and microbial composition. The results of the ANOVA made it possible to reach the following conclusions: reject the null hypothesis of the substrate and time period factors, since statistically significant differences were found between the concentration values of the final protein obtained with the different substrates: banana peel meal, grapefruit bagasse meal and mesquite pod meal; accept the alternative hypothesis in relation to the time factor: significant statistical differences were found between the initial and the different number of days of fermentation and in the interaction between both factors, time period per substrate. The rest of the interactions were not statistically significant.
Table 6 compares the characteristics of the fermentation process of this study from a practical perspective, considering the time needed to reach the maximum amount of proteins in each substrate and the microbial composition.
The analysis of variance indicates that there are no significant differences in the microbial protein concentration produced between the grapefruit bagasse and the banana peel substrates, or with the mesquite pod substrate, although statistical differences were observed between the concentrations of microbial protein produced with the banana peel and the mesquite pod substrates. This observation is repeatedly present in the bioreactors with the different microbial compositions.
Taking into account the productivity index used in this experiment, which establishes a relationship between the microbial protein unit (response variable) generated, in relation to the soluble sugars unit (i.e. reducing and non-reducing sugars), it should be considered that the material having the highest PI is the most efficient one, which is the one producing the largest amount of microbial protein (under the fermentation conditions established that are similar in all cases) per gram of sugar. Under the ideal fermentation conditions established for S. cerevisiae the value of Yx/s (where X and S represent the concentration of the biomass and the substrate respectively) should ideally be of about 0.5 g of dry biomass per gram of sugar absorbed, but since the specific product evaluated is microbial protein concentration, the yield of the product will be given by Yp/s, where P would be the protein produced per gram of sugar absorbed [3]. When considering that the amount of protein contained in the yeast is of approximately 40 % of the biomass, then the ideal value should be of approximately 0.2 g of protein per gram of sugar. In this study, the highest productivity index was 0.21 and it was obtained with grapefruit bagasse meal as the substrate and the yeast/ bacteria combination, but no statistically significant differences were found between the indices obtained.
Although all the substrates used achieved a similar microbial protein concentration, they all started with different initial protein levels and therefore the increase in protein produced by solid state fermentation implies a different percent increase for each substrate. Table 3 summarizes the data on the protein increase in relation to the material analyzed. As shown, a maximum percent increase in protein is obtained in the grapefruit bagasse meal and the lowest increase was in the mesquite pod meal, although the latter substrate reached the maximum absolute value of final total protein. The bar graph format in the table makes it possible to compare the data from the same column graphically.
DISCUSSION
The maximum fermentation time of 16 days was established as a function of the results obtained by Bairagi et al. [11] who used two strains of cellulase-producing Bacillus in the fermentation of Leucaena leaf meal where they obtained an increase in the protein concentration and other modifications of the substrate. To evaluate the process, the proteins were measured at 4 and 8 days. A factorial design was used for the analysis of the experimental data obtained in the solid state fermentation process of each substrate. There were two hypotheses. The first hypothesis assumed that significant differences should occur in the protein concentration between the initial time and the fermentation periods of 4, 8 and 16 days. The second expressed that differences between the protein concentrations obtained at levels S and S+3 would be expected. The main characteristic of the isolated Bacillus strain used in this study is the production of endoglucanase. Since the plants used are complex matrices, the assumption was that a combination of yeast and bacteria would enable the endoglucanases, of the Bacillus strain used, to act on the polysaccharides and increase the availability of simple sugars, in order to favor the yeast growth.
The results of the analyses of variance confirm the first hypothesis for all substrates since there were statistically significant differences between the initial and final protein concentrations at the levels with different microbial compositions, and in all cases, the difference was established between days 1 and 4.
The statistical analysis indicates that, in grapefruit and mesquite when the incubation time is longer than 4 days, there are no significant modifications in protein concentration on using either S. cerevisiae or the combination of S. cerevisiae/B. subtilis. This makes it necessary to learn the dynamics of the system between 0 and 4 days of the study in more detail.
Banana peel reached its maximum protein concentration on day 16 using S. cerevisiae/B. subtilis, with significant differences in the protein concentrations with less fermentation time. In the samples seeded with S. cerevisiae, a maximum protein concentration is also obtained on day 16, although there was no statistically significant difference with the other fermentation periods.
These experimental data led to the fact that it is possible to increase the protein content of the materials used in the study with S. cerevisiae and B. subtilis, under the experimental conditions stated. With grapefruit bagasse, there was an increase of over 150 % in the amount of protein of the original material. This was in contrast to that obtained by Oboh et al. [14], who found an increase from 6.03 to 12.8 g/100 g in the amount of protein of the grapefruit peel, which is an increase of 112 %, in a very similar experiment using S. cerevisiae at 14 days of fermentation.
It is possible that the composition in complex carbohydrates of the different substrates would involve an unequal susceptibility to the attack of the microbial endoglucanases. In the grapefruit substrate the prevailing polysaccharides are pectins, and in mesquite these are gums [27], both of which form part of the soluble edible fiber. The pectins and gums are healthy products because of their capacity to absorb water; this produces the sensation of fullness to repletion and they act as moderators of glucemia, while helping to prevent diseases of the digestive system, mainly colon cancer.
In the food industry, the pectins and gums are very useful as gellants (pectins) and as thickeners (gums), although in other types of industries, such as those of juices and wines, they must be eliminated. With this objective, we have used enzymes produced by B. subtilis, such as xylanases that act on the arabinoxylans (composed by 5 carbon sugars that cannot be degraded by the endogenous enzymes of the animals) mainly found in cereals. Cuyvers et al. demonstrated that a strain of B. subtilis produced xylanases that act on the soluble and insoluble fractions of the xylans [28]. These also produced a hydrolyzate of xylans that could be metabolized by the bacterial genera Lactobacillus and Bifidobacteria, offering a competitive advantage in relation to other bacterial genera that are unable to use this type of substrate as a probiotic. This means that the action of the xylanases on the arabinoxylans produces substances with probiotic potential [29].
Particularly, banana peels are made of starch in approximately 40 % of their dry matter, and another 40 % is formed by neutral detergent fiber (NDF) components, cellulose, hemicellulose and lignin [30]. This polysaccharide constitution is probably responsible for the fermentative behavior that differentiates it from the other two substrate matrices.
Other factors affecting enzymatic activity, besides the constitution of the carbon hydrates and the susceptibility of the microbial enzymes, are the pH, temperature, the presence of ions such as calcium, among others, in which their maximum activity is reached. Soriano Lasheras [31] determined that the pectate lyase enzyme obtained from B. subtilis presented its maximum activity at pH 10 and 65 °C in the presence of 2 mM Ca2+ [31], which are quite different from the conditions used in this study.
The Bacillus strain is not the only endoglucanase producer. The presence of glucanases was also demonstrated in S. cerevisiae. Hien and Fleet [32] isolated 6 types of (1-3) β-glucanases, two of them exoenzymes and the other 4 were 4 endoenzymes [32]. More recently, Restuccia et al. isolated a total of 80 strains belonging to the S. cerevisiae genus of wine must with β-glucosidase activity [33]. Also isolated from wine are the wild strains of S. cerevisiae that were evaluated under the conditions having the maximum activity of the β-glucosidase, where it was found that in grape juice at a pH 4.0, the aerobic conditions and the temperature of 40 to 50 °C favor the activity of these enzymes [34]. Several publications now deal with the use of S. cerevisiae having β-glucosidase in order to improve wine aroma [35, 36].
The Bacillus genus produces characteristic anti-microbial substances [37] that may confer protection and inhibit the development of undesirable microorganisms in the bioreactors containing it. This may be an explanation as to why the bioreactors that were contaminated were those that only contained S. cerevisiae and the matrix with the highest amount of sugars, i.e. mesquite.
The questions that arise from this study for future research are mainly related to the analysis of which and how these fermentation factors affect each particular matrix, in order to optimize the process and to be able to reach a higher scale of production. In this sense, Ray et al. found that humidity, temperature and the nitrogen source, play a crucial role in the production of cellulases in Bacillus strains isolated from fish intestine, although the variables concerning the amount of the inoculum and pH were also evaluated [38]. Another aspect of fermentation conditions that should be included in further studied is that related to the amount of O2, which is crucial for the development of these aerobic microorganisms, particularly the effect of shaking. Another important factor that should be evaluated is the possibility of performing the inoculation of the microorganisms at different time periods, and the influence of all these factors in order to decrease fermentation time. The main challenge may be the design of a bioreactor that will simplify scaling up, and enable the best evaluation capacity of the above variables.
Noteworthy, although the solid state fermentation process has been practiced for several centuries in the preparation of traditional food, its application in the production of new biotechnological developments is limited, as it is the case in Argentina.
CONCLUSIONS
This study has demonstrated that solid state fermentation with S. cerevisiae and B. subtilis enables the production of microbial protein. Using the grapefruit bagasse meal as a substrate with a 4 day fermentation, we obtained a product with a maximal protein concentration of 16.4 g/100 g, of which more than 60 % is of microbial origin; this substrate had the best productivity index with the use of S. cerevisiae/B. subtilis. Dry ground banana peels that were fermented for 16 days produced a material with the maximum protein content of 15.5 g/100 g of which 50 % is microbial protein. The pods of the dry ground mesquite produced a maximum protein content of 23.92 g/100 g of which 45 % is microbial protein, and this matrix had the highest amount of microbial protein produced.
Although the use of the combination of S. cerevisiae + B. subtilis strain 3 (S+3) did not show any statistically significant differences in the final protein concentration of each substrate, the inclusion of B. subtilis would offer an additional benefit based on its probiotic potential.
REFERENCES
1. Chacón Villalobos A. Perspectivas actuales de la proteína unicelular (SCP) en la agricultura y la industria. Agron Mesoamer. 2004;15(1):93-106.
2. Nasseri AT, Rasoul-Amini S, Morowvat MH, Ghasemi Y. Single Cell Protein: Production and Process. Am J Food Technol. 2011;6(2):103-16.
3. Ertola R, Yantorno O, Mignone C. Microbiología Industrial. Washington: Organización de los Estados Americanos; 1998.
4. Mondal AK, Sengupta S, Bhowal J, Bhattacharya DK. Utilization of fruit waste in producing single cell protein. Int J Sci Environ Technol. 2012;1(5):430-8.
5. Bacha U, Nasir M, Khalique A, Anjum AA, Jabbar MA. Compararive assessment of various agro-industrial waste for Saccharomyces cerevisiae biomass production and its quality evaluation as single cell protein. J Plant Anim Sci. 2011;21(4):844-9.
6. Kandari V, Gupta S. Bioconversion of vegetable and fruit peel wastes in viable product. J Microbiol Biotech Res. 2012;2(2):308-12.
7. Díaz-Plascencia D, Rodríguez-Muela C, Mancillas-Flores P, Angulo C, Salvador F, Ruíz O, et al. Desarrollo de un inoculo con diferentes sustratos mediante fermentación sólida sumergida. RedVet. 2011;12(1):1-10.
8. Julián Ricardo MC, Ramos Sánchez L. Fermentación en estado sólido (I). Producción de alimento animal. Tecnol Quím. 2007;XXVII(3):17-22.
9. Elías A, Lezcano O, Lezcano P, Cordero J, Quintana L. Reseña descriptiva sobre el desarrollo de una tecnología de enriquecimiento proteico en la caña de azúcar mediante fermentación en estado sólido (Saccharina). Rev Cubana Cienc Agr. 1990;24(1):1-12.
10. Oboh G, Ademosun O, Lajide L. Improvement of the nutritive value and antioxidant properties of citrus peels through Saccharomyces cerevisae solid substrate fermentation for utilization in livestock feed. Livestock Res Rural Dev. 2012;24(1):1-10.
11. Mussatto SI, Ballesteros LF, Martins SLF, Teixeira JA. Use of agro-industrial wastes in solid-state fermentation processes. In: Yeow SK, Xinxin G, editors. Industrial Waste. Rijeka: Intech; 2012. p. 121-40.
12. Poulose S, Singh B. Optimization of culture conditions for the production of single cell protein for marine yeast Candida MFF101 as feed supplement in acquaculture. J Aquat Biol Fish. 2014;2(1):283-9.
13. González I, Infante D, Peteira B, Martínez B, Arias Y, González N, et al. Caracterización bioquímica de aislamienots de Trichoderma spp. promisiorios como agentes de control biológico. II. Expresión de actividad glucanasa. Rev Protección Veg. 2011;26(11):23-9.
14. Dashtban M, Maki M, Leung KT, Mao C, Qin W. Cellulase activities in biomass conversion: measurement methods and comparison. Crit Rev Biotechnol. 2010;30(4):302-9.
15. Bairagi A, Sarkar Ghosh K, Sen SK, Ray AK. Evaluation of the nutritive value of Leucaena leucocephala leaf meal, inoculated with fish intestinal bacteria Bacillus subtilis and Bacillus circulans in formulated diets for rohu, Labeo rohita (Hamilton) fingerlings. Aquacult Res. 2004;35(5):436-46.
16. Shrestrha AK, Dahal NR, Ndungutse V. Bacillus Fermentation of Soybean: A Review. J Food Sci Technol Nepal. 2010;6:1-9.
17. Shin EC, Lee JH, Hwang CE, Lee BW, Kim HT, Ko JM, et al. Enhancement of total phenolic and isoflavone-aglycone contents and antioxidant activities during Cheonggukjang fermentation of brown soybeans by the potential probiotic Bacillus subtilis CSY191. Food Sci Biotechnol. 2014;23:531-8.
18. Evans E, Musa A, Abubakar Y, Mainuna B. Nigerian indigenous fermented foods: Processes and prospects. In: Makun HA, editor. Mycotoxin and Food Safety in Developing Countries. Rijeka: Intech; 2013. p. 153-80.
19. Freyre M, Astrada E, Blasco C, Baigorria C, Rozycki V, Bernardi C. Valores nutricionales de frutos de vinal (Prosopis ruscifolia): consumo humano y animal. Cienc Tecnol Aliment. 2003;4(1):41-6.
20. Urbieta J. El codiciado pomelo blanco de Formosa. En: La Nación. Economías regionales: Citricultura. 2010 Aug 7 [cited 2015 Aug 10]. Available from: http://www.lanacion.com.ar/1291722-el-codiciado-pomelo-blanco-de-formosa
21. Rios E. El 50% de las bananas que produce Argentina se cultiva en Formosa. En: La Nación. Economías regionales: Citricultura. 2010 Aug 7 [cited 2015 Aug 10]. Available from: http://SietepuntasDigital.com
22. De Camargo M, Sturion G, Bicudo M. Avaliaçao química e biológica da casca de banana madura. Archivos Latinoamericanos de Nutrición 1996;46(4):320-4.
23. AOAC International. Official Methods of Analysis (962.10). 18th ed. Rockville: AOAC International; 2011.
24. Nelson N. A photometric adaption of the Somogyi method for the determination of glucose. J Biol Chem. 1944;153:375-80.
25. Trinder P. Determination of Glucose in Blood Using Glucose Oxidase with an Alternative Oxygen Acceptor. Ann Clin Biochem. 1969;6:24-5.
26. Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L, Tablada M, Robledo CW. InfoStat versión 2013. Córdoba: Grupo InfoStat. 2015 [cited 2015 Jul 18]. Available from: http://www.infostat.com.ar
27. Freyre MR, Bernardi CMH, Baigorria CM, Rozycki VR, Piagentini AM, Presa M. Parámetros de interés nutricional en semillas de mesquite (Prosopis ruscifolia). Rev FAVE - Ciencias Agrarias. 2010;9(1-2):89-96.
28. Cuyvers S, Dornez E, Rezaei MN, Pollet A, Delcour JA, Courtin CM. Secondary substrate binding strongly affects activity and binding affinity of Bacillus subtilis and Aspergillus niger GH11 xylanases. FEBS J. 2011;278(7):1098-111.
29. Vandeplas S, Bodin JC. Acción de una xilanasa producida por Bacillus subtilis. Selecciones Avícolas. 2012;54(11):19-22.
30. Monsalve JF, Medina VI, Ruiz AA. Producción de etanol a partir de la cáscara de banano y de almidón de yuca. Dyna. 2006;73(150):21-7.
31. Soriano Lasheras M. Análisis de sistemas pectinolíticos bacterianos. Aislamiento y caracterización de las pectinasas PelA de Paenibacillus sp BP.23 y e YVpa de Bacillus subtilis. Barcelona: Facultad de Biología. Departamento de Microbiología. Universidad de Barcelona; 2004.
32. Hien NH, Fleet GH. Separation and characterization of six (1 leads to 3)-beta-glucanases from Saccharomyces cerevisiae. J Bacteriol. 1983;156(3):1204-13.
33. Restuccia C, Pulvirenti A, Caggia C, Giudici P. A β-glucosidase positive strain of Saccharomyces cerevisiae isolated from grape must. Ann Microbiol. 2002;52:47-53.
34. Hernandez LF, Espinosa JC, Fernandez-Gonzalez M, Briones A. Beta-glucosidase activity in a Saccharomyces cerevisiae wine strain. Int J Food Microbiol. 2003;80(2):171-6.
35. Vernocchi P, Ndagijimana M, Serrazanetti DI, Chaves López C, Fabiani A, Gardini F, et al. Use of Saccharomyces cerevisiae strains endowed with β-glucosidase activity for the production of Sangiovese wine. World J Microbiol Biotechnol. 2011;27(6):1423-33.
36. Vernocchi P, Patrignani F, Ndagijimana M, Chavez C, Suzzi G, Gardini F, et al. Trebbiano wine produced by using Saccharomyces cerevisiae strain endowed with β-glucosidasa activity. Ann Microbiol. 2015;65(3):1565-71.
37. Raaijmakers JM, De Bruijn I, Nybroe O, Ongena M. Natural functions of lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics. FEMS Microbiol Rev. 2010;34(6):1037-62.
38. Acharya S, Chaudhary A. Optimization of fermentation conditions for cellulases production by Bacillus licheniformis MVS1 and Bacillus sp. MVS3 isolated from Indian hot spring. Braz Arch Biol Technol. 2012;55(4):497-503.
Received in August, 2015.
Accepted in December, 2015.
Karina M Rompato. Laboratorio de Investigación en Alimentos, Facultad de Ciencias de la Salud, Universidad Nacional de Formosa. Av. Gutnisky 3200, Formosa, CP 3600, Argentina. E-mail: karinarompato@gmail.com.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}