










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Plantas Medicinales
versión On-line ISSN 1028-4796
Rev Cubana Plant Med vol.17 no.1 Ciudad de la Habana ene.-mar. 2012
ARTÍCULO ORIGINAL
Capacidad antioxidante de dos variedades de Fragaria x ananassa (Weston) Duchesne (fresa) sometidas a variaciones en la nutrición vegetal
Antioxidant capacity of two Fragaria x ananassa (Weston) Duchesne (strawberry) varieties subjected to variations in vegetal nutrition
MSc. Luz Marina Carvajal de Pabón,I Dr. C. El Hadi Yahia,II Dr. Regulo Cartagena,III Dr. Carlos Peláez,I Ing. Carlos A. Gaviria,III Dr. C. Benjamín Alberto RojanoIII
I Universidad de Antioquia. Medellín, Colombia.
II Universidad Autónoma de Querétaro. Querétaro, México.
III Universidad Nacional de Colombia sede Medellín. Medellín, Colombia.
RESUMEN
Introducción: la fresa pertenece a la familia Rosaceae, es una fuente de compuestos polifenólicos con actividad antioxidante, especialmente antocianinas, ácidos fenólicos y vitamina C; los cuales son protectores de la oxidación de muchos organelos. Las fresas poseen mayor actividad antioxidante que muchas frutas como: toronja, naranja, uva roja, kiwi. Sin embargo, los metabolitos secundarios en fresa se pueden potencializar haciendo una dosificación controlada de nutrientes en los suelos.
Objetivos: evaluar la actividad antioxidante, el contenido de antocianinas y el contenido de fenoles totales de dos variedades de fresa (Camarosa y Osogrande) variando la nutrición de suelos con nitrógeno, potasio, fósforo, azufre, calcio, magnesio y materia orgánica, para establecer la relación entre los niveles de fertilización y la respuesta de la capacidad antioxidante.
Métodos: se evaluaron las propiedades antioxidantes y algunos componentes polifenólicos de los extractos acuosos de muestras de fresas cultivadas en Colombia (Camarosa y Osogrande), provenientes de Antioquia, Colombia. El contenido de fenoles totales, se determinó por el método Follin Ciocalteu, las antocianinas por el método diferencial de pH y la actividad antioxidante usando el ensayo DPPH.
Resultados: la unidad experimental 2-2-1-1 (Camarosa) mostró una actividad antioxidante de IC50 65,90 mg de fruta seca, contenido de fenoles totales de 984,75 y un contenido de antocianinas entre 83,1892,83; superiores a los encontrados en zonas templadas.
Conclusiones: la unidad experimental 2-1-1 Camarosa, con niveles superiores de materia orgánica e inferiores de nutrición mineral, presentan la mayor actividad antioxidante, incrementando sus posibilidades terapéuticas y conservando un buena respuesta fenotípica.
Palabras clave: fresa, actividad antioxidante, fertilización, fenoles totales, antocianinas totales.
ABSTRACT
Introduction: the strawberry belongs to Rosaceae family, being a source of polyphenolic compounds with antioxidant activity, specially anthocyanins, phenolic acids and vitamin C, which protect many organelles from oxidation. Strawberries have more antioxidant activity than many fruits such as grapefruit, orange, red grape and kiwi. However, the secondary metabolites in strawberry can be potentiated by a controlled dosage of nutrients into the soil.
Objectives: to evaluate the antioxidant activity, anthocyanin content and total phenolic content of two varieties of strawberry (Camarosa and Osogrande) by changing nutrition of soils using nitrogen, potassium, phosphorus, sulfur, calcium, magnesium and organic matter, in order to establish the relationship between levels of fertilization and the response of antioxidant capacity.
Methods: the antioxidant properties and some polyphenolic components of the aqueous strawberry extracts from the region of Antioquia and grown in Colombia (Camarosa and Osogrande). The total phenolic content was determined by the Follin Ciocalteu´s method, the anthocyanins by the pH differential method and the antioxidant activity by DPPH assay.
Results: the 2-2-1-1 experimental unit (Camarosa) showed antioxidant activity of CI50 65.90 mg in dried fruit, the total phenolic content was 984.75 and anthocyanin content ranged from 83.18 to 92.83, these figures were higher than those found in temperate zones.
Conclusions: the 2-1-1 experimental unit Camarosa, with higher levels of organic matter and lower levels of mineral nutrients, had the greatest antioxidant activity, thus increasing its therapeutic options, and keeping good phenotypical response.
Key words: strawberry, fertilization, antioxidant capacity, total phenols, total anthocyanins.
INTRODUCCIÓN
El consumo de frutas y vegetales ha sido asociado por una amplia variedad de estudios epidemiológicos con la disminución en la incidencia de enfermedades crónicas y degenerativas, incluidas algunas formas de cáncer y enfermedades cardiovasculares, entre otras.1,2 Esos efectos se han atribuido a una amplia variedad de compuestos que se encuentran de forma natural en las frutas y vegetales y que, tradicionalmente, no han sido considerados como nutrientes; entre los cuales se pueden encontrar los polifenoles y de estos a los flavonoides, cuya acción ha sido relacionada con la actividad antioxidante que presentan.3-5 Los efectos benéficos asociados a estas sustancias han promovido la investigación de los factores que pueden estimular su producción o afectar su contenido en las frutas y vegetales, tales como madurez al cosecharse, diferencias genéticas, condiciones de precosecha y poscosecha.6,7
Las fresas han atraído la atención de consumidores e investigadores debido a que poseen mayor actividad antioxidante total comparada con otras como la toronja, naranja, uva roja, kiwi, banano, manzana, tomate, pera y melón, evaluadas como frutas frescas.8 La actividad antioxidante de extractos de fresa se ha asociado con el contenido de fenoles totales, adicionalmente, las antocianinas, que son colorantes naturales con potente propiedad antioxidante, son las responsables del color brillante y atractivo de las fresas.9,10
El contenido de sustancias activas, entre ellas los antioxidantes, está controlado genéticamente, pero su expresión fenotípica está condicionada por variables ambientales como la luz, la temperatura, la humedad o el nivel de fertilización.11 El contenido de compuestos con actividad antioxidantes se ha investigado ampliamente para una gran variedad de productos vegetales.12,13 Existe poca información sobre la influencia del medio ambiente en la actividad antioxidante de las fresas. Wang evaluó el efecto de 4 combinaciones diferentes de temperatura día/noche sobre la capacidad antioxidante de 2 variedades de fresa de día largo (Earliglow y Kent) y encontraron que las plantas que crecen en días fríos y noches frías (18/12 °C) generalmente tienen menos ácidos fenólicos, flavonoides y antocianinas; un incremento en la temperatura nocturna de 12 a 22 °C con la temperatura del día constante en 25 °C tiene como resultado un incremento significativo en ácidos fenólicos, flavonoides y antocianinas; lo cual resulta en un incremento significativo de la actividad antioxidante.14 Connor comparó diferentes parámetros de calidad para una especie de arándano (Vaccinium sp.) y encontró alta correlación entre la actividad antioxidante (AA), el contenido de fenoles totales (FT) y el contenido de antocianinas totales (AT); moderada correlación negativa entre el porcentaje de pérdida de peso y la AA, FT, AT y correlación moderada positiva entre la AA, FT, AT y el contenido de sólidos solubles.15 Hernanz reporta las diferencias en la composición fenólica de cinco variedades de fresa (Fragaria x ananassa [Weston] Duchesne) cultivados en 2 sistemas de suelo diferentes; se encontró variaciones en el contenido de antocianinas, quercetina, kaempferol y ácido p-coumárico.16
Usualmente los nutrientes disponibles en los cultivos son inadecuados para obtener un óptimo desarrollo de la planta y algunos de ellos afectan de manera específica el desarrollo de las flores y los frutos, lo cual incide de modo directo en la productividad del cultivo; tal es el caso del calcio y el zinc cuya deficiencia afecta los frutos en el cultivo de tomate.17 En la fresa, los genotipos difieren en su capacidad para captar y usar los nutrientes, lo cual repercute en la producción, captación y uso. En un análisis comparativo de 2 genotipos diferentes de fresa (RCP37, un ecotipo clonal de Fragaria chiloensis y Fragaria x ananassa [Weston] Duchesne) se observaron eficiencias diferentes respecto al uso del nitrógeno; esto sugiere que la RCP37 tiene una gran capacidad de almacenar el exceso de nitrógeno en las hojas en forma de ribulosa 1,5 bifosfato, lo que contribuye a su alta eficiencia del uso del nitrógeno.18
Muchos problemas de calidad probablemente están relacionados con factores nutricionales o ambientales de los cultivos, como ruptura de dermis en algunos frutos tropicales en las etapas tempranas de desarrollo, caída de flores que impiden la formación de frutos, endurecimiento del mesocarpio, formación deficiente de azúcares o de ácidos, conversión indeseable de azúcares en lignina; por lo cual es de gran interés conocer cómo afecta la disponibilidad de nutrientes el desarrollo de las plantas y sus frutos.
En esta investigación se varió de forma controlada la nutrición con nitrógeno, potasio, fósforo, azufre, calcio, magnesio y materia orgánica de 2 variedades de fresa (Fragaria x ananassa [Weston] Duchesne) cv Camarosa y Osogrande y su efecto sobre la actividad antioxidante y el contenido de antocianinas y fenoles totales en sus frutos, en condiciones de plantación establecida en el trópico andino.
MÉTODOS
Instalación del cultivo: el cultivo se instaló bajo cobertizo y sobre mortero, en eras de 1,00 por 8 m, separadas entre unidades experimentales por ladrillo cocido, forradas con plástico color negro en su cara interna y plateada en la externa. Las plantas se sembraron simulando una densidad de siembra de 80 000 plantas por hectárea.
Fertilización: la fertilización de la planta se distribuyó en 3 etapas: en las primeras 12 semanas se agregó 20 % del fertilizante requerido, en el período comprendido entre las semanas 12 y 18 se aplicó 40 %, y entre las semanas 20 y 24 40 % restante (tabla 1). Fuentes naturales: cloruro de calcio, roca fosfórica, roca potásica, azufre natural, carbonato de magnesio, fueron usadas para la fertilización.
La fertilización con materia orgánica se hizo con 375 g.pl-1 de lombriabono como abono de fondo, complementado con abono fermentado denominado revitalizador de suelos (abono orgánico preparado con: leche, ácido acético, estiércol de bovino, leguminosas, plantas aromáticas, roca fosfórica, cal, ceniza) usado por los agricultores y se caracterizó según su composición y propiedades fisicoquímicas.19
A los cultivos establecidos se les realizó la aplicación de 2 niveles diferentes (N) de materia orgánica parcialmente compostada (MO), elementos mayores principales nitrógeno-fósforo-potasio (N-P-K) y elementos mayores secundarios azufre-calcio-magnesio (S-Ca-Mg), según se presenta en la tabla 2.
Prácticas culturales: se estableció como práctica la eliminación de flores hasta que 80 % de las plantas presente desarrollo de al menos 2 coronas o el cultivo tenga un desarrollo de 18 semanas, después de lo cual se mantiene la floración, se establece un sistema de riego manual aplicando 80 mL diarios de agua a cada planta y se hace control preventivo de plagas y enfermedades.
Recolección de la fruta: la madurez de la fruta para cosecha se determinó por el color No. 6 según norma técnica colombiana 4103; se cosecharon todos los frutos con el estado de madurez comercial elegido, se pesaron, clasificaron por calidad sanitaria y solo los frutos libres de enfermedad o daño físico se utilizaron para las evaluaciones de laboratorio. La fruta se cosechaba en la mañana durante un período de 2 semanas durante una cosecha y se sumergía directamente en nitrógeno líquido luego de su recolección y se conservaban congeladas a -20 °C en contenedor de vidrio color ámbar hasta el análisis en el laboratorio.
Preparación de muestras: la porción comestible de la fruta se homogenizó por 2 min, se centrifugó a 10 000 g por 10 min y, finalmente, se filtró por papel Watman No. 4, a partir del cual se prepararon las diferentes soluciones de evaluación.
Evaluación de la capacidad antioxidante por el método de DPPH: se empleó el método de Brand-Williams con algunas modificaciones.20,21 Se evaluó la capacidad de las muestras para atrapar el radical DPPH, por medio de la disminución en la absorbancia leída, luego de 30 min de reacción, en un espectrofotómetro Jemway 6405, UV/VIS, a una longitud de onda de 517 nm. Para cada muestra evaluada se calculó la concentración del filtrado que inhibe 50 % de los radicales DPPH· presentes al inicio de la reacción (Ic50).
Contenido de fenoles totales: el contenido de fenoles totales se realizó por el método colorimétrico de Folin-Ciocalteu, mediante la construcción de una curva patrón usando como estándar ácido gálico.22 Se diluyó el extracto a una concentración en la cual el contenido de fenoles se encontraba dentro del intervalo de la curva patrón. Los resultados se expresaron como mg de ácido gálico/100 g de fruta fresca.
Contenido de antocianinas totales: las antocianinas totales se determinaron mediante el método diferencial de pH.23 La absorbancia se midió en un espectrofotómetro JeanWay 6405 UV/Vis a 530 nm y 700 nm en buffers pH 1,0 y 4,5, usando A= (A530nm - A700nm) pH1,0 - (A530nm - A700nm) pH4,5, con un coeficiente de extinción molar para el cianidin-3-glucósido de 26 900. Los resultados se expresaron como miligramos equivalentes de cianidin-3-glucósido por 100 g de fruta fresca.
Diseño experimental y análisis estadístico: se empleó un diseño experimental que permitiera establecer comparaciones entre diferentes tratamientos, variando en 2 niveles los aportes de nitrógeno, potasio, fósforo (Tto1), magnesio, calcio y azufre (Tto2) y materia orgánica (Tto3), a 2 variedades de fresa (Tto4), controlando las condiciones de suministro de agua y otros nutrientes y midiendo las condiciones de iluminancia (28 600 lx a las 13 h), humedad (33,3 % a nivel de planta a las 13 h) y temperatura en cobertizo (31,7 ºC a nivel de planta a las 13 h) durante el desarrollo de la evaluación.24,25 Se consideraron variables de respuesta a los tratamientos el contenido de fenoles y antocianinas totales y la actividad antioxidante. Se aplicó un diseño completamente al azar (DCA), con estructura factorial de los tratamientos 24, con 3 repeticiones, para un total de 48 unidades experimentales (eras), con 8 plantas por unidad experimental. Las variables exógenas resultaron controladas o conocidas. Las prácticas agronómicas fueron similares para todos los tratamientos.
El control experimental se realizó mediante la selección de material vegetal homogéneo, siembra simultánea de las unidades experimentales, aleatorización de la asignación de tratamientos, 3 repeticiones por unidad experimental y por cada prueba de laboratorio, preparación y dosificación de cada tratamiento por un solo investigador, aplicación de prácticas culturales preventivas programadas (fertilización, control fitosanitario, podas, riegos), manejo estandarizado de las muestras, estandarización previa de las técnicas experimentales y uso de referencias
La asignación de los tratamientos a las unidades experimentales se hizo al azar sobre la disponibilidad de área, utilizando un listado de números seudoaleatorios. La unidad experimental de control se asignó con 0 aporte de materia orgánica y niveles inferiores de tratamiento N-P-K y S-Ca-Mg. Los resultados se analizaron por el método del rango múltiple de Duncan. Se usó el software estadístico Stargrafics Centurion XVI. La diferencia significativa aceptada resultó p= 0,05 en la respuesta a los tratamientos aplicados.
RESULTADOS
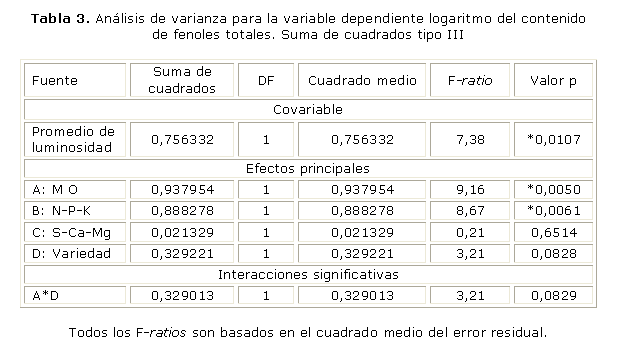
El contenido de fenoles es afectado por la luminosidad (valor p= 0,0107) y las variables que ejercieron efectos principales fueron: valor p= 0,0050 y la nutrición con N-P-K valor p= 0,0061 (tabla 3). Aunque está por fuera del rango del nivel de significancia elegido (p= 0,05), el valor p= 0,0828 para el factor variedad, podría ser suficiente para considerar esta variable.
Para la variedad Camarosa el efecto de la variación en la nutrición es menor, puesto que sus valores medios se distribuyen en 3 grupos de 5 posibles, con contenidos equivalentes cuando se somete a condiciones inferiores y superiores de aplicación de materia orgánica e inferior de N-P-K. Tratamiento 212 que produce 709,67 mg de ácido gálico/100 g de fruta fresca, valor que forma grupo homogéneo con el tratamiento 112 que produce 768,29 mg de ácido gálico/100 g de fruta fresca (tabla 4).
Tabla 4. Contenido de fenoles y antocianinas totales y actividad antioxidante en fresas de las variedades Camarosa
y Osogrande bajo diferentes condiciones de nutrición
| Tratamiento | Fenoles totales1 | Diferencia entre grupos | Antocianinas totales2 | Diferencia entre grupos | Ic50 DPPH●3 | |
| MO-N-P-K-S-Ca-Mg | ||||||
| Oso Grande | ||||||
| 1-1-1 | 605,5 | ab | 44,62 | Abc | 139,96 | C |
| 1-1-2 | 740,99 | abc | 30,43 | A | 109,25 | Ab |
| 1-2-1 | 984,07 | cd | 48,49 | Abc | 92,12 | Ab |
| 1-2-2 | 1138,62 | d | 41,21 | Abc | 65,87 | A |
| 2-1-1 | 500,31 | a | 48,43 | Abc | 148,71 | C |
| 2-1-2 | 455,03 | a | 36,98 | Ab | 144,72 | C |
| 2-2-1 | 742,7 | abc | 48,95 | Abc | 98,31 | Ab |
| 2-2-2 | 704,23 | ab | 45,43 | Abc | 118,67 | Ab |
| 733,93 | 43,07 | 114,70 | ||||
| Camarosa | ||||||
| 1-1-1 | 675,29 | ab | 77,59 | Cde | 128,38 | Ab |
| 1-1-2 | 768,23 | abc | 80,46 | Ef | 125,41 | Ab |
| 1-2-1 | 963,3 | cd | 92,83 | F | 89,67 | Ab |
| 1-2-2 | 943,89 | cd | 83,19 | Ef | 85,33 | Ab |
| 2-1-1 | 984,75 | cd | 58,51 | Abcd | 65,91 | A |
| 2-1-2 | 709,67 | ab | 77,37 | Cde | 130,72 | Ab |
| 2-2-1 | 688,57 | ab | 45,64 | Abc | 111,93 | Ab |
| 2-2-2 | 741,33 | abc | 74,19 | Cde | 94,11 | Ab |
| 809,38 | 73,72 | 103,93 | ||||
1: (mg ácido gálico/100 g fruta fresca), 2: mg/100 g fruta fresca, 3: (mg de fruta seca). Orden de presentación de variables
en los tratamientos: MO-N-P-K-S-Ca-Mg, valor 1 para nivel 1 (N1) de cada variable, valor 2 para nivel 1 (N2) de cada variable.
Las medias de la misma columna seguidas de diferentes letras fueron estadísticamente diferentes con un valor p< 0,05.
El contenido de antocianinas totales expresadas como mg equivalentes de cianidin3glucósido/100 g de fruta fresca en las variedades Camarosa varió entre 45,64 y 92,83; en la variedad Osogrande presentó una variación entre 30,43 y 48,95 (tabla 4). Los resultados obtenidos con la variedad Camarosa desarrollada en condiciones de Trópico Andino aplicando niveles inferiores de materia orgánica y superiores de N-P-K sin importar el nivel de S-Ca-Mg, corresponden con los resultados de Wang con las variedades Allstar y Honeoye; ellos encontraron que bajos niveles de suplementación del suelo con compost y S-Ca-Mg incrementó el contenido de antocianinas en estas variedades (28,1-83,3 mg).26
La aplicación de S-Ca-Mg y la aplicación de materia orgánica presenta una interacción significativa sobre la producción de antocianinas (p= 0,0465), lo cual puede deberse a la optimización de la proporción entre el S y el N, para asegurar la asimilación de S (como SO4) por reducción en los cloroplastos mediante una reacción dependiente energéticamente del ATP.24
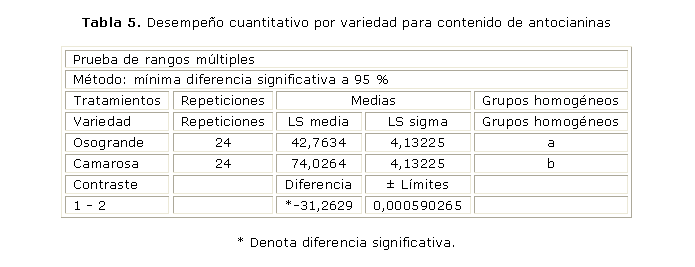
En la tabla 5 se presenta un procedimiento de múltiples comparaciones para la variable contenido de antocianinas referida a los tratamientos y se encuentra que las medias este contenido son significativamente diferentes para cada variedad con un nivel de confidencia de 95 %. También los promedios de producción de antocianinas por tratamiento (42,76 mg para Osogrande y 74,02 mg para Camarosa, expresados como equivalentes de cianididina-3-glucósido). La comparación de la respuesta por variedad muestra cómo la producción de antocianinas responde a la variabilidad genética, acorde con los resultados de Cano, que encontró variaciones en la composición y cantidad de las antocianinas presentes en los frutos de 4 variedades de frambuesa.27
DISCUSIÓN
Los efectos principales sobre el Ic50 significativos para la nutrición con elementos mayores principales (nitrógeno, fósforo y potasio con valor p= 0,0365), aunque están por fuera del rango del nivel de significancia elegido, un valor p= 0,0997 para la interacción entre los factores materia orgánica y la nutrición con S-Ca-Mg, podría ser suficiente para considerar esta variable.
La mejor respuesta promedio por variedad, la tiene la variedad Camarosa con un Ic50 medio de 103,93 mg de fruta base seca, contra un Ic50 promedio de 114,70 mg de fruta base seca para la variedad Osogrande (tabla 3), estos valores están muy cercanos y de acuerdo al análisis de varianza no pueden considerarse significativos para variedad, lo cual indica que la actividad antioxidante en las variedades de fresa evaluadas depende de factores externos.
La aplicación de niveles superiores de N-P-K puede aumentar significativamente la respuesta antioxidante, la cual es mayor para el tratamiento 122 en la variedad Osogrande (nivel inferior de materia orgánica, superior de N-P-K y superior de S-Ca-Mg) con un promedio de Ic50 igual a 65,87 mg de fruta medida en base seca.
El efecto de los mayores valores de Ic50 debido al aporte de niveles superiores de N-P-K se conserva en 6 de los 8 valores menores de Ic50 (tabla 4). Esta respuesta es significativamente diferente a la lograda para el tratamiento 121, con la variedad Osogrande, para iguales aplicaciones de materia orgánica y N-P-K pero menor aplicación de S-Ca-Mg, con un Ic 50 de 92,12, lo cual muestra cómo un nivel superior de S-Ca-Mg puede mejorar la respuesta de la capacidad antioxidante, quizá por la participación del azufre y el calcio en la formación de aminoácidos o en las vías de transducción de señal, respectivamente.
Wang encontró igualmente una respuesta positiva a la aplicación de niveles superiores de fertilizante y de compost a la capacidad atrapadora de los radicales: ROO., O2.-, H2O2, OH- al evaluar las variedades de fresa Allstar y Honeoye, igualmente encontraron correlación significativa entre la actividad antioxidante y el contenido de glutatión (GSH).26
En general, la menor respuesta en la capacidad antioxidante se presenta en condiciones de baja aplicación de N-P-K y se puede apreciar el efecto positivo de la interacción de la aplicación de niveles superiores de S-Ca-Mg cuando hay poca materia orgánica, al lograr incrementar la capacidad antioxidante desde 139.96 para el tratamiento 111 (Osogrande con niveles inferiores de materia orgánica, N-P-K y S-Ca-Mg) hasta 109,25 para 112 (Osogrande con niveles inferiores de materia orgánica y N-P-K y superiores de S-Ca-Mg) así como 125,41 para 112 (Camarosa con niveles inferiores de materia orgánica, N-P-K y superiores de S-Ca-Mg), las cuales forman un grupo de respuesta homogéneo. Los resultados de la actividad antioxidante, permiten afirmar que solo la aplicación de niveles superiores de N-P-K tienen un efecto significativo positivo en su incremento, con independencia de cual de las 2 variedades sea evaluada.
La unidad experimental 211 (Camarosa, con aplicación de niveles superiores de materia orgánica e inferiores de nutrición mineral) presentó la mejor respuesta antioxidante frente al radical DPPH., la que podría considerarse una respuesta bioquímica de las plantas, desarrollada en condiciones de restricción de nutrientes minerales, diferente a la esperada con respecto a la capacidad antioxidante; ocasionada quizá por condiciones de estrés producto de la materia orgánica aplicada. La evidencia de mayor actividad antioxidante, posiblemente es el resultado de un desbalance en los mecanismos de oxidación-reducción de las plantas, que puede lograr un cambio en la eficiencia enzimática o en la expresión del perfil genético y podría convertirse en una evidencia de la posibilidad de mantener un excedente de actividad antioxidante en presencia de factores ambientales negativos.
El mayor contenido de fenoles totales para la variedad Camarosa fueron obtenidos en iguales condiciones superiores de aplicación de S-Ca-Mg, al reducir la aplicación de materia orgánica y con incremento al nivel superior de aplicación de N-P-K (943,89 mg de equivalentes de ácido gálico/100 g de fruta fresca) para formar grupo homogéneo con el valor máximo absoluto encontrado en esta variedad (984,74 mg de equivalentes de ácido gálico/100 g de fruta fresca). Wang reportó un contenido de 103 mg de equivalentes de ácido gálico/100 g de fresa fresca evaluados por el método de Folin-Ciocalteau, y Meyers reportó un valor máximo de 273 mg de equivalentes de ácido gálico por 100 g de fruta al evaluar 9 razas diferentes de fresa.12,28 Estos datos se refieren a condiciones de manejo agronómico en plantaciones de zonas templadas, y se constituye en uno de los pocos referentes del tema.
La respuesta de la concentración de fenoles a los diferentes tratamientos presenta resultados positivos significativos para la variedad, nivel superior de N-P-K e inferior de materia orgánica, lo cual podría explicarse debido a que los compuestos fenólicos responden a una expresión genético dependiente inducida por factores ambientales como luz, nutrición y estrés. La producción de metabolitos secundarios es un mecanismo de defensa de las plantas.29
En la formación de polifenoles participan 2 rutas metabólicas, la vía de los policétidos, minoritaria en plantas superiores, y la vía del ácido shiquímico; a veces, ambas vías pueden participar de conjunto en la formación de fenoles. Este proceso está catalizado por un complejo enzimático que incluye a la enzima fenilalanina amonioliasa PAL- (enzima marcadora de la vía fenilpropanoide).30 Esta enzima es activada por la luz así como la concentración de diferentes hormonas vegetales. La actividad de la PAL aumenta cuando los vegetales son sometidos a situaciones de estrés, como la deficiencia de agua, infecciones fúngicas o bacterianas y radiaciones UV. Otro factor activador de la PAL es el frío. Lafuente y otros efectuaron el rastreo diferencial de una librería de cDNA preparada de tejido de flavedo de frutos de mandarina "Fortune" almacenados durante 21 d a 2 °C, aislaron e identificaron 5 cDNA que se expresan diferencialmente por las bajas temperaturas. Mediante transcripción reversa-reacción en cadena de la polimerasa (TR-PCR) se han aislado 2 cDNA que codifican para la fenilalanina amonio liasa (PAL), lo cual demuestra que esta es una enzima codificada por genes inducibles.31 La mayor parte de la información está disponible para la inducción de los genes dependientes de la luz que codifican la producción de ácido hidrocinámico, antocianinas en hojas y tejidos florales y la producción de proantocianidinas en semillas.
Hay algunas interacciones funcionales entre los diferentes tipos de factores de trasncripción que aún no han sido aclaradas, sin embargo, se podría inferir de los resultados obtenidos, que un factor de estrés relacionado con la alta radiación de la zona tropical, algunos factores de patogenicidad o toxicidad incluidos por la adición de materia orgánica (la cual podría no encontrarse en un estado adecuado de compostaje), o ambos, inducen un evento de respuesta e incrementan el contenido de fenoles necesarios para equilibrar la producción de especies reactivas de oxígeno.32 En este caso, la aplicación de nutrientes mayores principales (N-P-K) no se constituye en un factor de estrés, al parecer con mayores concentraciones aplicadas (en el rango definido), la tendencia es al incremento de fenoles, además de contribuir al desarrollo de la planta, así como podrían estar aportando compuestos requeridos por el sistema de señal de la planta mediante el proceso de desnitrificación de suelos mediado por microorganismos. En este se produce, entre otras, la molécula de óxido nítrico, que ha sido definida como un posible segundo mensajero en la cascada de señales que desencadenan respuestas del interior de la célula a las señales externas, las cuales están muy bien documentadas para la fisiología animal y solo empieza a dilucidarse para los vegetales.33-36
La respuesta de la concentración de antocianinas a los diferentes tratamientos presenta resultados positivos significativos para la variedad, así como la aplicación de nivel inferior de materia orgánica. El efecto producido por la aplicación de los diferentes niveles de N-P-K no mostró resultados significativos, lo cual indica que en los rangos de nutrición establecidos, aunque estaban en los extremos máximos y mínimos reportados por la literatura, no alcanzaron a ser factor de estrés por deficiencia. Esta respuesta también indica que la ruta biosintética de producción de antocianinas en la investigación es diferente a la de la producción de los fenoles, la cual genera la mayor capacidad antioxidante, como se puede corroborar con la falta de correlación entre la producción de fenoles totales y antocianinas, por lo tanto, el incremento en la producción de antocianinas puede estar relacionado con la mayor luminosidad de la zona tropical, una respuesta dependiente del genotipo de la planta y con las condiciones de estrés ocasionado, como ya se mencionó por los aportes de la materia orgánica usada.37,38
Wei encontró en un estudio por TR-RCP cuantitativo, una fuerte correlación entre la expresión transcripcional, el incremento de la biosíntesis de antocianinas y la radiación lumínica, quizá actuando la biosíntesis de antocianinas como respuesta al estrés por incremento en la radiación visible y UV, con la finalidad de minimizar la producción de radicales libres por procesos fotooxidativos.39,40
Los resultados muestran igualmente una correlación relevante positiva entre la variedad Camarosa y la aplicación de niveles superiores de S-Ca-Ma, quizá por el mayor aporte de azufre como insumo de la biosíntesis de glutation, el cual participa como dador del grupo azufrado con la intermediación de una glutatión transferasa, que se ha evidenciado como molécula crítica en el transporte de antocianinas a la vacuola.41,42 Wang, encontró que el uso de compost como enmienda del suelo puede incrementar los niveles de ácido ascórbico y glutation, indicando que esta molécula responde a la nutrición vegetal.26
El contenido de fenoles totales y la actividad antioxidante se encuentran relacionados y responden a un modelo exponencial con un valor p= 0,000, un nivel de confidencia de 99 % y coeficiente de correlación de - 0,785894, que indica una fuerte relación. No se encontró correlación significativa entre la actividad antioxidante y el contenido de antocianinas al evaluar los datos globales o individuales por variedad (Camarosa y Osogrande). Meyers encontró una fuerte correlación entre la actividad antioxidante determinada mediante la metodología TOSC y el contenido de fenoles totales, sin embargo, no encontró correlación significativa entre la actividad antioxidante y los flavonoides totales o la actividad antioxidante y el contenido de antocianinas totales.28
El impacto del contenido de fenoles en la actividad antioxidante puede estar relacionada en mayor proporción con la acción antioxidante de flavonoides con una alta actividad antioxidante encontrados en las fresas, diferentes a las antocianinas como: p-cumaroil-glucosa, dihidroflavonol, quercitin-3glucósido, quercitin-3-glucorónido, kamperol y kamperol 3-glucorónido, o efectos sinérgicos o antagónicos; por lo que no es posible predecir la actividad antioxidante de estas variedades de fresa estimando solo su contenido de antocianinas.43,44 En el caso de las fresas, el contenido de flavonoles derivados del ácido gálico se considera importante, y es conocido que los derivados del ácido gálico presentan una actividad antioxidante en medio acuoso superior a sus moléculas homólogas no-derivadas de ácido gálico.45,46 Wang reportó la presencia de compuestos fenólicos como antocianinas y taninos con potente actividad antioxidante en extractos acuosos de frutas.10 Kähkönen observó al evaluar la actividad antioxidante de 92 extractos de diferentes plantas que contienen compuestos fenólicos, que esta no necesariamente se encuentra correlacionada con el contenido de fenoles.47
El método de Folin-Ciocalteau para la determinación de fenoles totales, no evalúa solo compuestos fenólicos sino que determina simultáneamente otros compuestos reducidos; tampoco estima alguna posible interferencia por la presencia de otros compuestos químicos presentes en la muestra como azúcares o ácido ascórbico.48,49 Rivas-Gonzalo reporta igualmente, con resultados obtenidos por peroxidación lipídica, elevada actividad antioxidante para extractos de frutas con elevado contenido de antocianinas como fresa, cereza, frambuesa, mora y arándano; pero otras frutas con altos contenidos de antocianinas como la granada, grosella y uva tinta, no presentaron actividad antioxidante elevada. Por esta razón es necesario el estudio de la actividad antioxidante derivada de la presencia de antocianinas de los extractos acuosos de frutas por fracciones aisladas de vitaminas y azúcares.50 La correlación entre Ic50 y el contenido de antocianinas está limitada igualmente por la fracción antioxidante evaluada.51,52
Wang reporta correlación lineal entre el contenido de fenoles totales y la actividad ORAC (medida de la habilidad del componente antioxidante para inhibir un generador de radical peroxilo) y entre el contenido de antocianinas y los valores ORAC.12 Otros autores no hallaron correlación significativa entre la actividad antioxidante y el contenido de antocianinas. Rivas-Gonzalo, encontró que los valores de la actividad antioxidante y el contenido de flavonoides agrupados por la presencia o no de antocianinas para 28 diferentes frutas y evaluados por 2 métodos distintos (TEAC: trolox equivalent antioxidant capacity y Ic50) no presentaron una correlación significativa; en general las frutas de color rojo muestran una buena capacidad antioxidante, lo cual sugiere que este pigmento puede contribuir a esa actividad, sin embargo, otras frutas con presencia de antocianinas como la granada, la grosella roja y la uva roja no presentan una alta actividad antioxidante.45 Wang evaluó la actividad antioxidante de antocianinas y flavonas homólogas con la misma hidroxilación por el método ORAC y encontraron que ninguna de las antocianinas evaluadas tenía actividad mayor que las flavonas homólogas.10
La respuesta de la unidad experimental 2-1-1 (Camarosa, con aplicación de niveles superiores de materia orgánica e inferiores de nutrición mineral) a la actividad antioxidante permite concluir, que hay una respuesta bioquímica de las plantas desarrollada en condiciones de restricción de nutrientes minerales, que tiene posibilidad de mantener un excedente de compuestos con actividad antioxidante en presencia de factores ambientales negativos, incrementando de esta forma sus posibilidades terapéuticas y conservando al mismo tiempo un buena respuesta fenotípica (a la firmeza y al rendimiento; en ambos casos la respuesta se localiza en 50 % de la respuesta superior).
AGRADECIMIENTOS
Esta investigación se realizó con apoyo económico y técnico del CODI (Universidad de Antioquia), C.I Antioqueña de Fresas, La Universidad Autónoma de Querétaro; los autores agradecen a Sara Luz Restrepo su trabajo en el laboratorio.
REFERENCIAS BIBLIOGRÁFICAS
1. Valko M, Leibfritz D, Moncol J, Cronin MT, Mazur M, Telser J. Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol. 2007;(39):44-84.
2. Slattery ML, Caan BJ. Nutrition and Colon Cancer. En: Coulston AM, Rock CL, Monsen ER, editores. Nutrition in the Prevention and Treatment of Disease. San Diego: Academic Press; 2001. p. 357-72.
3. Havsteen BH. The biochemistry and medical significance of the flavonoids. Pharmacol Ther. 2002;(96):67-202.
4. Pascual-Teresa S, Sanchez-Ballesta MT. Anthocyanins: from plant to health. Phytochem. 2007;(7):281-99.
5. Wang J, Mazza G. Inhibitory effects of anthocyanins and other phenolic compounds on nitric oxide production in LPS/IFN-activated RAW 264.7 macrophages. J Agric Food Chem. 2002;(50):850-7.
6. Navarro JM, Flores P, Garrido C, Martinez V. Changes in the contents of antioxidant compounds in pepper fruits at different ripening stages, as affected by salinity. Food Chem. 2006;(96):66-73.
7. Kalt W, Ryan DAJ, Duy JC, Prior RL, Ehlenfeldt MK, Vander Kloet SP. Interspecific variation in anthocyanins, phenolics and antioxidant capacity among genotypes of highbush and lowbush blueberries (Vaccinium Section Cyanococcus spp.). J Agric Food Chem. 2001;(49):4761-7.
8. Wang H, Cao G, Prior RL. Total antioxidant capacity of fruits. J Agric Food Chem. 1996;(44):701-5.
9. Rekika D, Khanizadeh S, Deschenes M, Levasseur A, Charles T, Tsao R, et al. Antioxidant capacity and phenolic content of selected strawberry genotypes. Hortscience. 2005;(40):1777-81.
10. Wang H, Cao GH, Prior RL. Oxygen radical absorbing capacity of anthocyanins. J Agric Food Chem. 1997;(45):304-9.
11. Suzuki T, Iwai K. Constituents of Red Pepper Species: Chemistry, Biochemistry, Pharmacology, and food Science of the Pungent Principle of Capsicum Species. En: Arnold Brossi, editor. The Alkaloids: Chemistry and Pharmacology. San Diego: Academic Press; 1984. p. 227-99.
12. Wang SY, Lin H. Antioxidant activity in fruits and leaves of blackberry, raspberry, and strawberry varies with cultivar and developmental stage. J Agric Food Chem. 2000;(48):140-6.
13. Wang SY, Jiao H. Scavenging capacity of berry crops on superoxide radicals, hydrogen peroxide, hydroxyl radicals, and singlet oxygen. J Agric Food Chem. 2000;(48):5672-6.
14. Zheng W, Wang SY. Effect of plant growth temperature on antioxidant capacity in strawberry. J Agric Food Chem. 2001;(49):4977-82.
15. Connor AM, Luby JJ, Hancock JF, Berkheimer S, Hanson EJ. Changes in fruit antioxidant activity among blueberry cultivars during cold-temperature storage. J Agric Food Chem. 2002;(50):893-8.
16. Hernanz D, Recamales AF, Melendez-Martinez AJ, Gonzalez-Miret ML, Heredia FJ. Assessment of the differences in the phenolic composition of five strawberry cultivars (Fragaria × ananassa Duch.) grown in two different soilless systems. J Agric Food Chem. 2007;(55):1846-52.
17. Phililips DR, Hawson MG. Fertilizers for Tomatoes. Advisers, Horticultural División. U.S.A.: Department of Agriculture; 1998.
18. Fallahi ER, Moon JW, Huang Y, Jensen R. Effects of ribulose 1,5 biphosphate carboxylase/ oxygenase content, leaf nitrogen and leaf morphology on CO2 assimilation in strawberry genotipes. Acta Hort. 2000;(527):177-83.
19. Brito J. Pruebas de metodología para la extracción de substancias húmicas en los suelos. Agronomía Trop. 1972;(22):517-33.
20. Brand-Willams W, Cuvelier ME, Berset C. Use of free radical method to evaluate antioxidant activity. Food Sci Technol. 1995;(28):25-30.
21. Rojano BA, Gaviria CA, Gil MA, Saez JA, Schinella G, Tournier H. Actividad antioxidante del isoespintanol en diferentes medios. Vitae. 2008;(15):173-81.
22. Dewanto V, Wu X, Adom KK, Liu RH. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J Agric Food Chem. 2002;(50):3010-4.
23. Giusti M, Wrolstad RE. Characterization and Measurement of Anthocyanins by UV-Visible Spectroscopy. En: Wrolstad RE, editor. Current Protocols in Food Analytical Chemistry. New York: John Wiley & Sons, Inc.; 2001. p. F1.2.1-F1.2.13.
24. Salisbury FB, Ross CW. Fisiologia Vegetal. México D.F.: Grupo Editorial Iberoamérica; 1994. p. 336-7.
25. Sánchez H. Fresa: requerimientos y normas de cultivo. Proplantas: y comercializadores de plantas; 2001. p. 8.
26. Wang SY, Lin HS. Compost as a soil supplement increases the level of antioxidant compounds and oxygen radical absorbance capacity in strawberries. J Agric Food Chem. 2003;(51):6844-50.
27. de Ancos B, González E, Cano MP. Differentiation of raspberry varieties according to anthocyanin composition. Z Lebensm Unters Forsch. 1999;(208):33-8.
28. Meyers KJ, Watkins CB, Pritts MP, Liu RH. Antioxidant and Antiproliferative Activities of Strawberries. J Agric Food Chem. 2003;(51):6887-92.
29. Häkkinen S. Flavonols and Phenolic Acids in Berries and Berry Products. Finland: Kuopio University Printing Office; 2000.
30. Lespinasse M, Moncada X, Pérez LM. Actividad de PAL en Sclerotium rolfsii: ¿un mecanismo adicional de infección que utiliza este patógeno?. Simiente. 1999;69(3-4):32-69.
31. Lafuente MT, Mallent D, Sala JM, Zacarías L, Marcos JF, Alférez F, et al. Cambios fisiológicos y moleculares asociados a la susceptibilidad de frutos tropicales y subtropicales a la baja temperatura de conservación. Instituto de Agroquímica y Tecnología de Alimentos: Proyecto IATA [citado 4 Mar 2010]; 1999. Disponible en: http://www.iata.csic.es/iata/dcie/pcos/pacti.htf?pyi
32. Kevin M, Davies E, Schwinn KE. Transcriptional regulation of secondary metabolism. Functional Plant Biology. 2003;(30):913-25.
33. Zumft WG. The biological role of nitric oxide in bacteria. Arch Microbiol. 1993;(160):253-64.
34. Palmer RMJ, Ferrige AG, Moncada S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature. 1987;(327):524-6.
35. Cueto M, Hernández-Perea O, Martin R, Ventura ML, Rodrigo J, Lamas S, et al. Presence of nitric oxide synthase activity in roots and nodules of Lupinus albus. FEBS Lett. 1996;(398):159-64.
36. Rodrigo J, Alonso D, Fernández AP, Serrano J, López JC, Encinas JM, et al. El óxido nítrico: síntesis, neuroprotección y neurotoxicidad. ANALES. 2000;(23):195-236.
37. Jansen NAK, Gaba V, Greenberg M. Higher plants and UV-B radiation balancing damage, repair and acclimation. Trends Plant Sci. 1998;(3):119-59.
38. Tevini M. Erhöhte UV-B Strahlung: Ein Risiko für Nutzpflanzen? Biologie Unserer Zeit. 1996;(26):245-54.
39. Zhang W, Furusaki S. Production of Anthocyanins by Plant Cell Cultures. Biotechnol. Bioprocess Eng. 1999;(4):231-52.
40. Gould K. Nature's Swiss Army Knife: the diverse protectiveroles of anthocyanins in leaves. J Biomed Biotechnol. 2004;(5):314-20.
41. Conn S, Curtin C, Bézier A, Franco C, Zhang W. Purification, molecular cloning, and characterization of glutathione S-transferases (GSTs) from pigmented Vitis vinifera L. cell suspension cultures as putative anthocyanin transport protein. J Exp Bot. 2008;(59):3621-34.
42. Taiz L, Zeiger E. Plant Physiology. 2 ed. Massachusetts: Sinauer Associates Inc. Publisers; 2003.
43. Pratt DE. Natural antioxidants from plant material. En: Huang IMT, Ho CT, Lee CY, editors. Phenolic compounds in food and their effects on health. New York: Am. Chemical Society; 1992. p. 54-71.
44. Pineda-Alonso D, Slucci M, Lázaro R, Maiani G, Ferro-Luczzi A. Actividad antioxidante y potencial de sinergismo entre los principales constituyentes antioxidantes de algunos alimentos. Rev Cubana Aliment Nutr. 1999;(13):104-11.
45. Rivas-Gonzalo JC, García-Alonso M, Pascual-Teresa S, Santos-Buelga C. Evaluation of the antioxidant propeties of fruits. Food Chem. 2003;(84):13-8.
46. Plumb GW, De Pascual-Teresa S, Santos-Buelga C, Cheynier V, Williamson G. Antioxidant properties of catechins and proanthocyanidins: Effect of polymerisation, galloylation and glycosylation. Free Radical Res. 1998;(29):351-8.
47. Kähkönen MP, Hopia AI, Vuorela HJ, Rauha J, Pihlaja K, Kujala TS, et al. Antioxidant activity of plant extracts containing phenolic compounds. J Agric Food Chem. 1999;(47):3954-62.
48. Santos-Buelga C, Scalbert A. Proanthocyanidins and tanin-like compounds: nature, ocurrence, dietary intakeand effectson nutricion and health (review). Agric Food Chem. 2000;(80):1094-117.
49. Singlenton VL, Rossi JA. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am J Enol Vitic. 1965;(16):144-58.
50. Einbond L, Reynertson KA, Luo XD, Basile MJ, Kennelly E. Anthocyanin antioxidants from edible fruits. Food Chem. 2004;(84):23-8.
51. Sichel G, Scalla M, Di Biblio AJ, Bonomo RP. In vitro Scanverger activity of some flavonoids and melanin against O2•-. Free Radical Biol Med. 1991;11(1):1-8.
52. van Acker SA, Tromp MN, Haenen GR, van der Vijgh WJ, Bast A. Flavonoids as scavengers of nitric oxide radicals. Biochem Biophys Res Commun. 1995;(214):755-9.
Recibido: 3 de febrero de 2011.
Aprobado: 27 de septiembre de 2011.
Luz Marina Carvajal de P. Facultad de Química Farmacéutica. Universidad de Antioquia. Medellín, Colombia. Calle 67 Nº 53·108 - Bloque 2 Oficina 134 Correo electrónico: lcarvaja@farmacia.udea.edu.co


{kind=link}
{kind=link}
{kind=link}
{kind=link}