










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkMedicentro Electrónica
versión On-line ISSN 1029-3043
Medicentro Electrónica vol.21 no.2 Santa Clara abr.-jun. 2017
ARTÍCULO ORIGINAL
Heredabilidad del fenotipo longevo en familias de excepcional larga vida, procedentes de Villa Clara
Heritability of longevity phenotype in families from Villa Clara with exceptionally long life
Manuela Herrera Martínez1, Douglas Fernández Caraballo1, Kenia Estrada López1, Mayelín Yera Vázquez2, Sonia Chaviano Machado1, Luz Marina Pérez Rodríguez3, Inti González-Herrera4
1. Unidad de Investigaciones Biomédicas. Universidad de ciencias Médicas Dr. Serafín Ruiz de Zárate Ruiz. Santa Clara, Villa Clara. Cuba. Correo electrónico: manuelahm@infomed.sld.cu
2. Servicio de Genética. Ranchuelo, Villa Clara. Cuba.
3. Servicio de Genética. Santa Clara, Villa Clara. Cuba.
4. Universidad de Bordeux. Francia.
RESUMEN
Introducción: la longevidad humana es un fenotipo de determinación compleja donde están implicados factores genéticos y ambientales, no completamente comprendidos.
Objetivos: determinar la heredabilidad de la longevidad en un grupo de familias de excepcional larga vida.
Métodos: con el objetivo de estudiar la epidemiología genética de la longevidad, se confeccionó el árbol genealógico a 340 longevos procedentes de familias de excepcional larga vida de Villa Clara; se analizó la concordancia del fenotipo en hermanos y cónyuges, la correlación del fenotipo mediante coeficiente de correlación de Pearson en 2 125 parejas de hermanos, evaluadas según sexo, y en 302 parejas longevo-cónyuge. La heredabilidad fue determinada en sentido estrecho, sobre la base de la correlación intraclase entre hermanos y de la correlación entre cónyuges.
Resultados: el fenotipo tuvo elevada concordancia entre hermanos del probando, superior a la encontrada entre los cónyuges; la heredabilidad, en un diseño que considera parejas que incluyen el probando, fue de 0,35. En el diseño que utilizó a todas las parejas independientes de hermanos, donde se excluyó el probando, la heredabilidad fue de 0,39 para familias con probando masculino y de 0,58 para familias de probandos femeninos.
Conclusiones: la heredabilidad moderada muestra el incremento de años vividos, como un genotipo de determinación continua. La concordancia elevada para la longevidad entre los esposos, sin que existiera parentesco entre ellos, indica que el componente ambiental del fenotipo no es insignificante.
DeCS: longevidad/genética.
ABSTRACT
Introduction: human longevity is a complex phenotype with multiple determinants where incomprehensible genetic and environmental factors are involved.
Objective: to determine heritability of longevity in a group of families with exceptionally long life.
Methods: a genealogical tree of 340 people belonging to families from Villa Clara with exceptionally long life was made for studying genetic epidemiology of longevity; concordance of phenotype in siblings and spouses was analyzed, as well as, the correlation of phenotype in 2 125 couples of siblings evaluated according to gender and in 302 couples of long-lived probands and spouses was analyzed by means of Pearson correlation coefficient. Heritability in the narrow sense was determined according to the intraclass correlation coefficient between siblings and according to the correlation between spouses.
Results: phenotype had a high concordance between siblings of the proband, being higher than the one found between spouses; heritability estimate was 0.35 in a design that consider couples including the proband. In the design, where all independent couples of siblings were used, but the proband was excluded, heritability was 0.39 for families with a male proband and 0.58 for families with female probands.
Conclusions: moderate heritability shows an increase of years lived as a continuous phenotype. High concordance for longevity between spouses without any parenthood between them demonstrates that environmental factor of the phenotype is not insignificant.
DeCS: longevity/genetics.
INTRODUCCIÓN
La longevidad humana satisfactoria es una aspiración del hombre, casi tan vieja como la humanidad; solo en los últimos años, los portentosos avances en la medicina y la biología han realizado aportes significativos al incremento notable de la esperanza de vida al nacer, aparejado, sin duda, a una mayor calidad de vida, dado el desarrollo tecnológico. Este incremento ha sido más notorio en los países desarrollados, si bien se ha experimentado en todos los países.
En el mundo existen 600 millones de longevos. La población centenaria en las Américas, para mediados del siglo, habrá ascendido a cerca de 689 000.1
Cuba presenta una de las mayores expectativas de vida y también una de las poblaciones más envejecidas de América Latina. La esperanza de vida es de 78,97 años para los hombres y de 80,0 para las mujeres; con una población mayor de 60 años de 19,3% para 2016, es uno de los países más envejecidos de América Latina y tendrá la población más longeva del continente en el 2025. En los próximos 15 años, las personas mayores de 80 años van a aumentar de forma acelerada, y llegarán a ser casi medio millón. En Cuba viven 2 500 centenarios.
La provincia de Villa Clara es considerada como la de mayor expectativa de vida al nacer, y la esperanza de hacer factible los 80 años para todos es una meta de salud inmediata, en la que existe un empeño especial.2
La duración de la vida parece estar genéticamente determinada; al menos, algunas características asociadas así lo evidencian. Los individuos de una misma especie tienen una duración característica, pero existe, no obstante, una considerable variación; aquellos que, dentro de una misma especie, sobreviven más que el promedio, se dice que son longevos. Numerosos estudios de cohorte _que han seguido la evolución de adultos envejecidos por un tiempo suficiente para identificar muchos factores determinantes en la duración excepcional de la vida y de la esperanza de vida, así como los que han examinado a personas longevas y controles mediante estudios de asociación del amplio genoma_ han identificado variantes genéticas asociadas con la longevidad. Con el creciente reconocimiento de la importancia de las variaciones raras, los estudios que utilizan diseños familiares de longevidad serán muy útiles. Esta área, reconocida como epidemiología de la longevidad, se ha expandido rápidamente. No obstante, existen controversias, por lo que dilucidar el papel de los diferentes genes será un trabajo arduo, como corresponde a un fenotipo complejo.3
Más recientemente, el secuenciado del exoma y de la expresión del amplio genoma, así como los estudios de epigenética, han comenzado a permitir entender mejor la variación funcional y la regulación de genoma en las vías biológicas de la longevidad.4
Los estudios genéticos resultan complejos, y su investigación en el campo de la biología celular y molecular requiere de cuantiosos recursos tecnológicos. Si se procede con estrategias investigativas que permitan identificar a las familias con más de una persona longeva (nonagenaria o centenaria), el costo de los estudios podría resultar menor, puesto que se estima que dichas personas presentan la combinación magistral de genes protectores contra los fenómenos oxidativos de la vejez, así como la ausencia de variantes genéticas y bioquímicas desencadenantes de las enfermedades comunes en los ancianos, que constituyen las principales causas de muerte en la tercera edad. Los descendientes de estas familias, por la carga genética que reciben, podrían ser sujetos de dichas investigaciones para lograr un mayor aprovechamiento de los recursos humanos y materiales que se destinen a este propósito.
La heredabilidad describe la proporción de la variación fenotípica de un carácter que se debe a factores genéticos, y ha sido muy empleada en los estudios de genética cuantitativa en muchas especies con fines científicos y prácticos. Hoy día es una técnica muy empleada en epidemiología genética humana, que está ayudando a caracterizar el componente genético de la variabilidad total de un carácter en una población determinada; ella mide los roles relativos de la herencia y el ambiente que son específicos en cada lugar, en función, sobre todo, de la diversidad de la variabilidad ambiental, y de un componente genético que, en los fenotipos de determinación compleja, es de base poligénica; por tanto, la heredabilidad es, por definición, la proporción de la varianza fenotípica total de un rasgo provocado por la varianza genética aditiva.
No se han realizado antes, en Villa Clara, estudios genéticos de la longevidad humana, ni existe el registro de familias informativas de este fenotipo; este trabajo informa los resultados de una investigación que tuvo el objetivo de determinar la concordancia y heredabilidad del fenotipo de longevidad humana, según el sexo, en familias de excepcional larga vida.
MÉTODOS
Se realizó un estudio genético de base poblacional _coordinado por el Laboratorio de Epidemiología Genética, de la Unidad de Investigaciones Biomédicas_ de 340 familias nucleares de larga vida procedentes de cuatro municipios (Santa Clara: 206, Sagua: 45, Ranchuelo: 29 y Remedios: 60), las que fueron seleccionadas de las familias informativas de longevidad de dichos municipios, por muestreo no probabilístico a conveniencia.
Se confeccionó el árbol genealógico de los individuos considerados casos índices o probandos de estas 340 familias nucleares o informativas para la longevidad, se incluyeron en ellos a todos los parientes de cualquier grado, y se establecieron los grados de parentesco a partir del probando, que fue siempre un longevo vivo de estas familias, residente en uno de los cuatro municipios participantes. De esta manera, se conformó lo que se considera en epidemiología genética, una cohorte transversal, reconstruida de familias nucleares para el fenotipo longevo, que proporcionaron un total de 8 084 familiares de primer, segundo o tercer grados, vivos o fallecidos.
Como criterios de inclusión se consideraron:
- Anciano con fenotipo de longevidad satisfactoria: Individuo que vive más que el promedio de los individuos de la población de la cual proviene, teniendo en cuenta la expectativa de vida en la provincia de 75 años como promedio; se consideraron aquellos ancianos con validísimo septuagenario (a partir de 75 años), octogenario, nonagenario o centenario.
- Validísimo: se midió aplicando los índices de Katz y Lawton, instrumentos previstos para evaluar la capacidad funcional de los ancianos.
- Familia informativa de longevidad satisfactoria: Existe certeza de que hay más de un longevo en la familia, vivo o fallecido, de primer, segundo o tercer grados, o sea, existe agregación familiar.
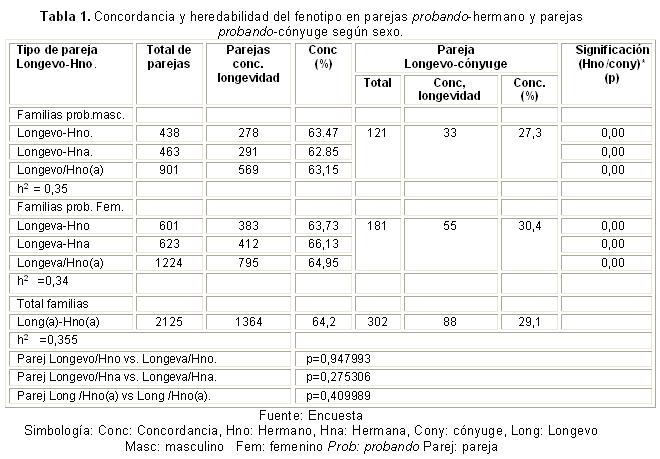
Para los estudios de concordancia del fenotipo, fueron seleccionadas las parejas de longevo probando con hermanos y con un hermano diferente con pareamiento por sexo (vivo o fallecido). Del mismo modo, se formaron parejas probando cónyuge.
Se constituyeron parejas entre el longevo probando y el hermano, considerando por separado las parejas según el sexo: longevo/hermano, longevo/hermana, longeva/hermana y longeva/hermano. Se determinó la concordancia simple (porciento) para la longevidad, considerando parejas concordantes a aquellas en las cuales ambos miembros alcanzaron los 75 años, sin limitación importante en las funciones físicas y psíquicas. Fueron discordantes aquellas parejas en que solo el probando era longevo. La misma consideración fue hecha para parejas longevo-cónyuge.
Las proporciones de concordancia entre los distintos tipos de parejas de hermanos con probando longevo masculino, se compararon mediante prueba de comparación de proporciones para muestras independientes, con la concordancia general de las parejas de longevo masculino y cónyuge femenino. Se procedió de la misma forma con los probandos femeninos.
La heredabilidad, en esta situación donde se incluyó el probando, se obtuvo de la diferencia de la concordancia simple, siguiendo la siguiente ecuación:
h2=Correlación parientes-Correlación cónyuges
h2=(Parientes C/C+D) - (Cónyuges C/C+D)
Donde: C=Concordantes y D= Discordantes
Para este estimado de heredabilidad, fueron consideradas las 2 125 parejas longevo- hermano (901 parejas integradas por un propósito masculino longevo con un hermano o hermana, y 1 224 parejas entre un propósito femenino longevo con un hermano o hermana) y 302 parejas longevo-cónyuge (121 parejas longevo masculino_cónyuge y 181 parejas longevo femenino_cónyuge).
Se calculó, además, la heredabilidad en sentido estrecho por un diseño de vías, empleando la correlación del fenotipo en todas las parejas independientes de hermanos que se formaron en las familias nucleares de excepcional larga vida, respecto a las parejas probando cónyuge. En estos cálculos, el probando es excluido de las parejas de hermanos, como se ha sugerido en la literatura, para evitar sesgos de muestreo.
De modo que cuando se excluyó el probando, la heredabilidad en sentido estrecho se determinó por la fórmula habitual, considerando la varianza genética como correlación intraclase en parejas independientes de hermanos, según sexo, siguiendo la siguiente ecuación:
h2 = VG / VP
donde: Vp (Varianza fenotípica total)= varianza genética(VG) + varianza ambiental(VA)
VG: varianza entre hermanos y VA: varianza entre cónyuges
La heredabilidad es la relación entre la varianza genotípica y la varianza fenotípica; cuando se estima en sentido estrecho, la varianza genética es la varianza del efecto aditivo de los genes, la que se estima según el grado de semejanza entre familiares, expresado en forma de coeficiente de correlación; como en la variabilidad del fenotipo, interviene la correlación ambiental entre individuos, se determinó la correlación del fenotipo entre cónyuges, la cual es debida al efecto de factores ambientales comunes, ya que no comparten dotaciones genéticas del mismo origen; dicha correlación ambiental, se supuso, no tiene diferencias significativas con la existente entre los parientes. La varianza fenotípica total se consideró la suma de la correlación existente entre hermanos (varianza genética aditiva), y la correlación entre cónyuges, la varianza ambiental. El resultado del cociente entre la varianza genética aditiva y la varianza fenotípica total se interpretó como la heredabilidad debida exclusivamente al efecto aditivo de los genes (sentido estrecho).
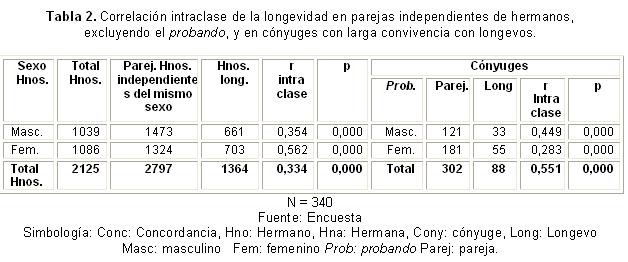
Para calcular la heredabilidad (en sentido estrecho) se consideraron 3 539 parejas independientes de hermanos, resultado del número de observaciones de covarianzas realizadas en función del tamaño de los grupos de hermanos y el sexo. Fueron procesados 1 039 hermanos masculinos que formaron 1 562 parejas, y 1 086 hermanos femeninos que formaron 1 977 parejas independientes.
Para calcular las parejas de hermanos independientes, se empleó la fórmula n= m (m-1)/2, donde m es el tamaño del grupo de hermanos.
Para los análisis de heredabilidad, se calculó la correlación entre parejas de hermanos así como en cónyuges, mediante el cálculo del coeficiente de correlación intraclase de Pearson, con el paquete SPSS 21.0. Se trabajó con una confiabilidad del 95 %.
La interpretación de los valores de heredabilidad se hace considerando que valores de h2 por debajo de 0,30 se consideran bajos; la heredabilidad entre 0,30 y 0,59 se valora como moderada, y es elevada cuando su valor es igual o mayor que 0,60.5
RESULTADOS
La evaluación de la concordancia del fenotipo longevidad entre el longevo masculino (probando) y la pareja que se forma con sus hermanos, mostró que concordaron el 63,47 % y con sus hermanas el 62,85 %. Los longevos concordaron con sus esposas en el 27,3 %. Cuando las parejas se formaron entre una longeva con sus hermanos, coincidieron el 63,7 %; con las hermanas, el 66,1%. Las longevas concordaron con sus esposos en el 30,4 % de los casos (tabla 1).
Se muestran los resultados de la comparación de la proporción de concordancia de las parejas formadas por el longevo y su correspondiente hermano respecto a las parejas integradas por el longevo y su cónyuge, las que muestran diferencias significativas.
Al considerar el sexo del longevo, no se encontraron diferencias estadísticas entre la proporción de concordancia para el fenotipo alcanzada entre los probandos con sus hermanos para los hombres longevos, respecto a las mujeres longevas; las comparaciones entre parejas de hermanos, respecto a los sexos, no difieren significativamente.
La concordancia general de la longevidad es de 64,2 %, toda vez que de los 2 125 hermanos de las familias nucleares, 1 364 llegaron a ser longevos. La heredabilidad, medida por la concordancia simple del fenotipo entre longevos probandos y sus hermanos y parejas longevo-cónyuge, fue de 0,355, asumiendo que la concordancia observada entre los esposos de 29,13 % (concordaron 88 de 302 cónyuges) fue la misma que estuvo presente entre hermanos
En la tabla 2, aparece la correlación intraclase respecto a la longevidad en las parejas de hermanos de igual sexo, excluyendo el probando, que se forman como parejas independientes de hermanos en las familias estudiadas de excepcional larga vida. La correlación fue positiva, moderada y significativa en los hermanos masculinos, en los femeninos y en la totalidad de hermanos, sin considerar el sexo. La correlación intraclase en los cónyuges de probandos masculinos (sexo femenino) fue significativa, al igual que la encontrada en los cónyuges de probandos femeninos (individuos masculinos).
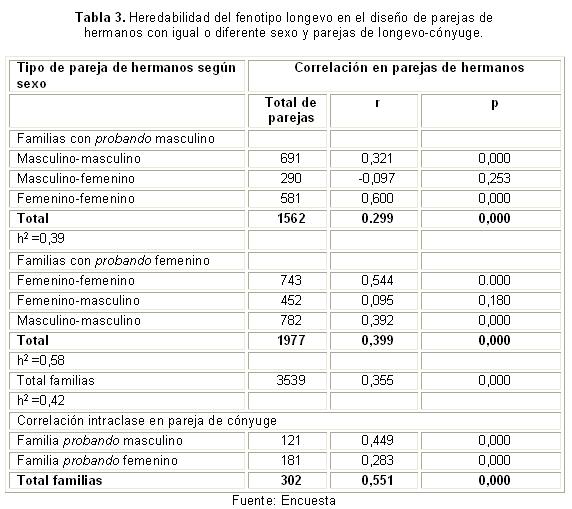
En la tabla 3, se presenta la heredabilidad en sentido estrecho mediante el diseño de parejas de hermanos, considerando el sexo del probando (el cual es excluido en los análisis de correlación intraclase). La heredabilidad fue de 0,42 en el total de las familias nucleares, según este diseño. La heredabilidad fue de 0,39 en las familias de probandos masculinos y de 0,58 en las familias de probandos femeninos. Como varianza ambiental en este diseño, se utilizó la correlación intraclase en cónyuges, que fue de 0,44 en familias de probandos masculinos y de 0,28 en familias con probando femenino. En esta tabla pueden observarse los valores de la correlación genética, resultado de la correlación intraclase del fenotipo entre hermanos en las distintos tipos de familias.
DISCUSIÓN
Los estudios de base poblacional tienen una gran importancia, en la actualidad, para dilucidar el papel de los genes y el ambiente en la determinación de la longevidad y el envejecimiento humanos.5
Los estudios de concordancia mostraron que, en los longevos, las parejas formadas entre el longevo con sus hermanos, independiente del sexo, tuvieron porcientos superiores a 60, a pesar de que los estilos de vida de los hermanos pueden variar mucho al llegar a la adultez, y emprender sus vidas independientes en nuevas familias, o que algunos hermanos no hayan alcanzado aun el fenotipo longevo, aunque posean los determinantes genéticos.
Los hallazgos descritos de mayor concordancia entre parejas de parientes de primer grado que entre las parejas de los cónyuges, avalan el componente genético. Estos determinantes genéticos son poligenes de determinación y funciones complejas, algunos de los cuales se han publicado. Así el gen de la apolipoproteína E (APOE) y la HSP 70.1 fueron mostrados asociados a la longevidad humana.6
El componente genético que está en la base de la mayor concordancia del fenotipo longevo entre hermanos que entre cónyuges ha sido ampliamente estudiado, pero en los estudios de genes candidatos, basados en estudios de asociación; hasta el presente, la mayoría de los datos apuntan a vías específicas, no solamente a nuevos loci puntuales de longevidad, si bien existen dificultades para replicar los hallazgos de los estudios de asociación, con independencia de la población estudiada, y persisten dificultades para identificar polimorfismos de longevidad universales.7
El gen AKT1 ha sido considerado como un posible candidato funcional, además de APOE y FOX3, y se ha encontrado asociación en varios estudios, aunque una evaluación en nonagenarios y centenarios daneses y alemanes no confirmaron esta asociación; por ello, no puede decirse que sea un gen universal asociado a la longevidad. Se planteó la existencia de clúster de genes para la longevidad, se rastrearon las regiones cromosómicas de 6p y 11 p, en busca de genes candidatos para la longevidad, y se encontraron tres genes (TNFalpha, TNFbeta, HSP70.1) en 6p21.3 y cinco genes (SIRT3, HRAS1, IGF2, INS, TH) en 11p15.5, pero no se han confirmado universalmente.8
Numerosos estudios de asociación de polimorfismos genéticos y longevidad no solo han considerado el número de años cumplidos, sino que han buscado asociaciones específicas de los alelos favorecedores a los hallazgos concretos en las pruebas que evalúan la capacidad cognitiva y funcional de ancianos nonagenarios y centenarios.9
La concordancia en los cónyuges fue importante, si se tiene en cuenta que no hay base genética del mismo origen; la concordancia para las esposas de los hombres longevos fue bastante elevada. En los esposos de las mujeres longevas de estas familias nucleares, la concordancia resultó igualmente inferior, lo que avala la agregación familiar y el componente de heredabilidad de la longevidad humana; pero, sin dudas, lo anterior permite hacer otras interpretaciones, puesto que este fenotipo presenta también una elevada concordancia entre cónyuges que solo comparten un ambiente común, por lo que quedan abiertas importantes puertas al componente ambiental de la longevidad.
En el sentido de explicar el componente ambiental presente en estas familias, que puede inferirse indirectamente a partir de los resultados alcanzados, se comentan algunas evidencias de la literatura. La metilación de islotes CpG es generalmente observada cerca de los promotores de genes que regulan la expresión de otros genes,10 aunque también se han descrito ciertos niveles de metilación en islotes CpG fuera de los promotores de los genes, pero relacionados con su expresión. En el tópico que nos ocupa, la deficiencia de folato puede perturbar la expresión de genes que están controlados por metilación. Se ha observado que los niveles de metilación del ADN cambian con la edad, lo que implica que las alteraciones en los patrones de metilación son una consecuencia inevitable del envejecimiento. Una variedad de alteraciones epigenéticas afectan todas las células y tejidos a lo largo de la vida.11
El envejecimiento es un proceso que ocurre por cambios en varios niveles de las células debidos a la función de los genes. La longevidad es la habilidad de copiar y reparar el daño que resulta de estos cambios; está bien descrito como resultado de un óptimo funcionamiento del sistema inmune y la sobrexpresión de variantes de secuencias antinflamatorias de genes de inmunidad e inflamación.
Se han considerado algunos factores ambientales relacionados con la expresión de genes involucrados en las vías biológicas de la longevidad, como la resistencia al estrés y elementos de la dieta, como es el caso del zinc; se ha informado una asociación entre polimorfismos del gen A2M (alfa 2 macroglobulinemia) y los procesos inflamatorios mediados por la homeostasis del Zn. Recientes trabajos en Caenorhabditis elegans han identificado la existencia de parejas genes-dieta, donde la consecuencia de una mutación de un gen específico solamente se realiza con una dieta específica. Muchas vías metabólicas han sido conservadas entre esta especie y los humanos; por tanto, aunque hasta ahora solo se han descubierto completamente unos pocos de estos pares, se espera que, potencialmente, no sean cientos sino miles de estas interacciones que podrían explicar la variabilidad en la proporción de envejecimiento humano y la incidencia de enfermedades relacionadas con la edad.12
Los valores de heredabilidad encontrados atestiguan que, en el contexto de esta investigación, el fenotipo longevo tuvo una heredabilidad moderada de 0,35 cuando se incluyó el probando en los estudios, lo que resulta ligeramente superior a lo informado por algunos autores.
La longevidad y el envejecimiento favorable se encuentran entre los fenotipos más complejos estudiados hasta la fecha. La heredabilidad de la edad, al morir en la adultez, es aproximadamente de 25 % según varias investigaciones.13
Los estudios realizados en individuos con excepcional larga vida muestran que la heredabilidad es mayor, mientras mayor sea la edad del longevo. Los estudios de ligamiento de familias con excepcional larga vida soportan ahora la existencia de un locus de longevidad en el cromosoma 3, mientras que otros posibles loci difieren entre estudios. Evaluaciones de genes candidatos han identificado variantes de APOE y FOXO3A asociados con la longevidad; otros genes muestran resultados inconsistentes. Estudios de asociación del amplio genoma (GWAS) de centenarios, respecto a controles jóvenes, revelan solamente APOE como significativo a nivel de genoma completo.
Aunque en 1996 se planteó que los factores genéticos contribuyen a la variación de la duración de la vida, aproximadamente en un 25 %, esta contribución se considera ahora mínima antes de los 60 años, y más importante desde los 85 años en adelante. Fuera de definir que la heredabilidad es de 25-32 %, el componente genético de la longevidad no está profundamente explicado.3
Respecto a los diseños de estudio de la heredabilidad, se han informado numerosos métodos, y en algunos de ellos se ha sugerido no incluir en los análisis los probandos, por lo que en esta investigación, además de la determinación de la diferencia simple de la concordancia del fenotipo entre el probando longevo con la encontrada en sus parejas de cónyuges, que se acaba de discutir, se hicieron otros análisis donde el probando fue excluido, y se evaluaron los hermanos, como se discutirá más adelante.14
Al analizar los resultados de la influencia del sexo en los estudios de concordancia del fenotipo en las distintas parejas, aunque estos no mostraron diferencias significativas, indican que pudieran relacionarse, de alguna forma, con los hallazgos reiterados de una mayor longevidad en el sexo
femenino, lo que podría tener alguna conexión con una base genética en su determinación. La comparación de la concordancia, según sexo, en las parejas longevo/hermano respecto a las parejas longeva/hermano, y longevo/hermana respecto a las parejas longeva/hermana, no mostraron diferencias estadísticas significativas, con mayor concordancia para el sexo femenino. Resumiendo: en todos los casos, la concordancia para la longevidad con un tipo de pariente específico fue mayor en las longevas que en los longevos.
Por otra parte, la correlación intraclase del fenotipo, en la totalidad de los hermanos femeninos de esta cohorte de familias con excepcional larga vida, fue mayor que la de los hermanos masculinos; este es otro elemento que apunta a las diferencias de la prevalencia del fenotipo entre los sexos y que se convierte en justificación para realizar los análisis de longevidad por separado, según el sexo del longevo. Debe observarse que para los cónyuges ocurre lo mismo; la correlación intraclase es mayor en las familias con probandos masculinos, que obviamente son los cónyuges del sexo femenino, a pesar de que han convivido con probandos masculinos, en los que la longevidad fue menor.
Al encontrar menor correlación de la longevidad entre parejas de hermanos masculinos, _contrario a lo que sucede en hermanas hembras, donde hubo una correlación del fenotipo elevada, que es incluso superior al 0,5 esperado para hermanos si se tratara de un modelo dominante_ se sugiere que, al menos en el sexo femenino, la heredabilidad del fenotipo longevo debe ser importante y, con ello, la contribución de poligenes de efecto aditivo.
En los resultados de la heredabilidad que finalmente se encuentran en el estudio, intervienen los factores ambientales, que pueden hacer disminuir el peso relativo de la influencia de los genes; es por ello que la influencia genética es valorada, en lo fundamental, por la correlación del fenotipo entre hermanos; así, por ejemplo, mecanismos epigenéticos están rápidamente emergiendo como un aspecto crítico del envejecimiento y la longevidad. Los centenarios retardan cambios en la metilación relacionados con la edad; ellos pueden transmitir la habilidad de preservar la metilación a sus descendientes, a través de factores no genéticos.13
Los polimorfismos de los genes ADIPOQ y LEP pueden desempeñar roles específicos, diferentes en cada género, en las vías biológicas de la longevidad en el hombre.15
La heredabilidad de la longevidad, considerando solo hermanos y excluyendo el probando, fue superior a la encontrada mediante el diseño que incluyó los probandos, pero también resultó moderada.
Sin embargo, al considerar el sexo del longevo, estos resultados de la heredabilidad, por uno u otro método, son diferentes para los sexos, y si bien resultaron solo ligeramente superiores en las familias con probando masculino (0,399 vs 0,345), fueron evidentemente superiores en las familias con probandos femeninos (0,58 vs 0,35). Ello repercute en que la heredabilidad general del conjunto de familias sea superior a la encontrada en el otro método (0,42 vs 0,355). En este diseño, la heredabilidad también resultó moderada.
Los hallazgos de diferencias para la heredabilidad de la longevidad en parejas, según el sexo, coincide con una característica común descrita en los trastornos genéticos de base multifactorial; se ha evaluado que la esperanza de vida o tasa de envejecimiento está determinada por miles de genes que funcionan en mecanismos altamente complejos, únicos para cada célula o tejido del organismo. Podría pensarse en genes dialélicos determinantes de la longevidad, que resulten diferenciados en los sexos; estudios recientes en ratas para evaluar el efecto citotóxico de quimioterápicos en el cerebro envejecido, han mostrado diferencias en el comportamiento de los genes en los sexos; la mayoría de los cambios inducidos por la quimio en los tejidos de la corteza prefrontal ocurren durante el envejecimiento del cerebro, y se observó que decrece la metilación y se incrementa la hidroximetilación del DNA, en específico en los tejidos de ratas hembras; de modo que los cambios en la expresión génica son más pronunciados en los tejidos cerebrales de las hembras;16 por otro lado, están los hallazgos de los efectos de asociación y regiones genómicas iguales por descendencia en 234 tríos integrados por un centenario-hijo-sobrino, concordantes o discordantes para la longevidad.8
Aunque varios estudios plantean que la repercusión genética en la duración máxima de la vida está determinada entre un 25 y 30 % por la expresión fenotípica, referida a polimorfismos que modulan el tiempo de vida individual, en este grupo de familias la influencia fue mayor.
Recientemente se ha mostrado que la esperanza de vida es influenciada simultáneamente por muchos polimorfismos de simple nucleótidos (SNPs, por sus siglas en inglés), cada uno con un pequeño efecto individual, y se ha dilucidado el posible mecanismo biológico de su acción; muchos de estos 27 SNP se encontraban dentro de los genes (74 %). La relevancia funcional de los respectivos genes de envejecimiento y enfermedades mayores soporta la relación causal entre SNP identificados y la esperanza de vida.17
Esta influencia de múltiples genes en la longevidad se ha probado, no solo para modelos poligénicos, sino para modelos epistáticos.18
Estos resultados sugieren un componente genético de base poligénica. Estudios de epidemiología y demografía genética similares se han efectuado en otros países. Se ha encontrado un número elevado de asociaciones entre longevidad y variantes alélicas de genes, donde se destacan las evaluaciones de polimorfismos en centenarios; no obstante, se recogen controversias, y muchos estudios no han podido replicarse, tal vez por errores metodológicos o por la existencia de diferencias étnicas reales.19
Aparte del status de APOE, las variaciones en el gen FOXO3A están solamente confirmadas como una contribución genética a la supervivencia dentro de la edad adulta mayor, y la actividad de este está subregulado en varios cánceres, lo que indica que la larga vida pudiera estar relacionada con genes que disminuyen la probabilidad de sufrir determinadas enfermedades características del envejecimiento.20
Por último, cabe especificar que los valores de heredabilidad _como resultado de una frecuencia relativa_ dependen de ambos componentes del cociente y que, por tanto, solo son válidos en un contexto determinado y en condiciones específicas; de ahí el valor de haber determinado esta en un grupo de familias procedentes de nuestra provincia.
CONCLUSIONES
Esta investigación aportó evidencias para afirmar que la longevidad tuvo una elevada prevalencia en las familias nucleares estudiadas; existió una elevada concordancia del fenotipo longevidad entre hermanos, que fue superior, en todos los casos, a la encontrada entre parejas de cónyuges. La concordancia encontrada para este fenotipo entre los esposos fue lo suficientemente alta como para inferir que, no existiendo parentesco entre ellos, el componente ambiental en la determinación del fenotipo no es insignificante. La mayor prevalencia de longevidad entre los parientes femeninos de esta cohorte, así como la mayor concordancia del fenotipo en todas las parejas formadas con tipos de parientes específicos, cuando el longevo es femenino, sugieren un fuerte componente genético en la longevidad humana que avala la contribución de los genes a la base de la duración de la vida; ello indica el incremento de años vividos con validismo, como un genotipo de determinación continua aditiva o multiplicativa, como sugieren los valores encontrados de heredabilidad moderada.
REFERENCIAS BIBLIOGRÁFICAS
1. Pardo Andreu G. Consideraciones generales sobre algunas de las teorías del envejecimiento. Rev Cubana Invest Bioméd [internet]. 2003 [citado 14 ene. 2015];22(1):[aprox. 6 p.]. Disponible en: http://bvs.sld.cu/revistas/ibi/vol22_1_03/ibi08103.htm
2. Comité Estatal de Estadísticas. Datos de la provincia de Villa Clara. La Habana: MINSAP; 2014.
3. Salvioli S, Olivieri F, Marchegiani F, Cardelli M, Santoro A, Bellavista E, et al . Genes, ageing and longevity in humans: Problems, advantages and perspectives. Free Radic Res. 2006 Dec.;40(12):1303-23.
4. Newman AB, Murabito JM. The epidemiology of longevity and exceptional survival. Epidemiol Rev. 2013;35:181-97.
5. Tan Q, Kruse TA, Christensen K. Design and analysis in genetic studies of human ageing and longevity. Ageing Res Rev. 2006 Nov.;5(4):371-87.
6. Dato S, Carotenuto L, De Benedictis G. Genes and longevity: a genetic-demographic approach reveals sex- and age-specific gene effects not shown by the case-control approach (APOE and HSP70.1 loci). Biogerontology. 2007 Feb.;8(1):31-41.
7. Soerensen M, Dato S, Tan Q, Thinggaard M, Kleindorp R, Beekman M, et al. Human longevity and variation in GH/IGF-1/insulin signaling, DNA damage signaling and repair and pro/antioxidant pathway genes: cross sectional and longitudinal studies. Exp Gerontol. 2012 May;47(5):379-87.
8. Nygaard M, Soerensen M, Flachsbart F, Mengel-From J, Tan Q, Schreiber S, et al. AKT1 fails to replicate as a longevity-associated gene in Danish and German nonagenarians and centenarians. Eur J Hum Genet. 2013 May;21(5):574-7.
9. Dato S, Soerensen M, Lagani V, Montesanto A, Passarino G, Christensen K, et al. Contribution of genetic polymorphisms on functional status at very old age: a gene-based analysis of 38 genes (311 SNPs) in the oxidative stress pathway. Exp Gerontol. 2014 Apr.;52:23-9.
10. Deaton AM, Bird A. CpG islands and the regulation of transcription. Genes Dev. 2011;25:1010-22.
11. Najar RA, Rahat B, Hussain A, Thakur S, Kaur J, Kaur J, et al. Gene speci?c epigenetic regulation of hepatic folate transport system is responsible for perturbed cellular folate status during aging and exogenous modulation. Mol Nutr Food Res. 2016;00:1-131.
12. An Yen Ch, Curran SP. Gene-diet interactions and aging in C. elegans. Exp Gerontol. 2016 Dec. 15;86:106-12.
13. Brooks-Wilson AR. Genetics of healthy aging and longevity. Hum Genet. 2013 Dec.;132(12):1323-38.
14. Posthuma D, Boomsma DI. A Note on the Statistical Power in Extended Twin Designs. Behav Genet. 2000;30(2).
15. Khabour OF, Mesmar FS, Alatoum MA, Gharaibeh MY, Alzoubi KH. Associations of polymorphisms in adiponectin and leptin genes with men's longevity. Aging Male. 2010 Sep.;13(3):188-93.
16. Kovalchuk A, Rodriguez-Juarez R, Ilnytskyy Y, Byeon B, Shpyleva S, Melnyk S, et al. Sex-specific effects of cytotoxic chemotherapy agents cyclophospha-mide and mitomycin C on gene expression, oxidative DNA damage, and epigenetic alterations in the prefrontal cortex and hippocampus - an aging connection. Aging (Albany NY). 2016 Apr.;8(4):697-711.
17. Yashin AI, Wu D, Arbeev KG, Ukraintseva SV. Polygenic effects of common single-nucleotide polymorphisms on life span: when association meets causality. Rejuvenation Res. 2012 Aug.;15(4):381-94.
18. Tan Q, Soerensen M, Kruse TA, Christensen K, Christiansen L. A novel permutation test for case-only analysis identifies epistatic effects on human longevity in the FOXO gene family. Aging Cell. 2013 Aug.;12(4):690.
19. He YH, Lu X, Yang LQ, Xu LY, Kong QP. Association of the insulin-like growth factor binding protein 3 (IGFBP-3) polymorphism with longevity in Chinese nonagenarians and centenarians. Aging (Albany NY). 2014 Nov.;6(11):944-56.
20. Däumer C, Flachsbart F, Caliebe A, Schreiber S, Nebel A, Krawczak M. Adjustment for smoking does not alter the FOXO3A association with longevity. Age (Dordr). 2014 Apr.;36(2):911-21.
Recibido: 14 de octubre de 2016
Aprobado: 10 de diciembre de 2016
Manuela Herrera Martínez. Unidad de Investigaciones Biomédicas. Universidad de ciencias Médicas Dr. Serafín Ruiz de Zárate Ruiz. Santa Clara, Villa Clara. Cuba. Correo electrónico: manuelahm@infomed.sld.cu


{kind=link}
{kind=link}