










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkFitosanidad
versión impresa ISSN 1562-3009versión On-line ISSN 1818-1686
Fitosanidad v.14 n.3 Ciudad de la Habana jul.-sep. 2010
Diagnóstico fitosanitario
Diferenciación morfológica, cultural y biológica de guignardia citricarpa y guignardia mangiferae en frutos cítricos de Cuba
Morphologic, cultural and biologic differentiation of guignardia citricarpa and guignardia mangiferae in citric fruits of Cuba
Elsa I. Hidalgo Góngora y Luis Pérez Vicente
Laboratorio Central de Cuarentena Vegetal, Centro Nacional de Sanidad Vegetal. Ayuntamiento 231 e/ San Pedro y Lombillo, Plaza de la Revolución, Ciudad de La Habana, lperezvicente@live.com
RESUMEN
Se obtuvieron treinta y cinco aislamientos monospóricos de Guignardia citricarpa (Gc; Phyllosticta citricarpa) de síntomas de mancha negra (patogénicos) y cuatro de G. mangiferae (Gm; Phyllosticta capitalensis), asociados a puntos negros (no patogénicos) de la corteza de frutos cítricos. Se estudió el crecimiento y morfología de las colonias en PDA, agar avena (AA) y agar jugo V8 (V8), así como la formación, morfología y dimensiones de los teliomorfos y anamorfos de ambos hongos. Se realizó un análisis de componentes principales y de agrupamiento para determinar las características que más contribuyeron a la diferenciación de estas especies. Se concluyó que las dos especies concurren en frutos cítricos en Cuba y que las características que más aportan a su diferenciación fueron el crecimiento en PDA, la formación de peritecios en medio de cultivo, el tamaño mínimo del picnostroma y la formación de pigmento que se difunde al medio en V8 y AA. Gc es de crecimiento lento en PDA, forma picnidios abundantemente en el medio, el conidioma frecuentemente es menor de 145 µm, no forma peritecios en PDA (heterotálico) y produce un pigmento amarillo que se difunde al medio en agar V8. Gm es de crecimiento rápido en PDA, tiene un conidioma frecuentemente mayor de145 µm, forma peritecios en cultivo monospórico (homotálico) y no difunde pigmento amarillo al medio V8. Estas características permiten la diferenciación de ambas especies.
Palabras claves: Citrus spp., Guignardia, Phyllosticta, morfología fúngica, diagnóstico
ABSTRACT
Thirty five single spore isolates of Guignardia citricarpa (Gm, Phyllosticta citricarpa) from black spot symptoms and four from G. mangiferae (Gm; Phyllosticta capitalensis) associated to black dots (non pathogenic) were obtained in the rind of citrus fruits. The growth and morphology of the colonies, teleomorph and anamorph structures morphology and dimensions in both fungi were studied in PDA, oatmeal agar (AA) and V8 juice agar (V8). A principal components and cluster statistical analysis were performed to determine the characteristics that contribute the most to species differentiation. It were concluded that both species are concurrent in citrus fruits in Cuba and that the most important characteristics to differentiate them were growth in PDA, the perithecia development in vitro, the minimal size of the conidiostrome and the production and release of a yellow pigment in V8: Gc is slow growing in PDA, develop picnidia abundantly in media, the conidiostrome is frequently smaller than 145 µm, do not develop perithecia in PDA (heterotallic) and release a yellow pigment in V8. Gm es fast growing in PDA, frequently has a conidiostrome higher than 145 µm, develops perithecia in single spore cultures (homotallic) and do not produce and release a yellow pigment in V8. Those characteristic allow the differentiation of both species in citrus plants.
Key words: Citrus spp., Guignardia, Phyllosticta, fungic morphology, diagnostic
INTRODUCCIÓN
La mancha negra de los cítricos (MNC), causada por Guignardia citricarpa Kiely (anamorfo Phyllosticta citricarpa (McAlpine) Van der Aa), es una enfermedad del follaje y de los frutos de los géneros Fourtunella sp., Poncirus y sus híbridos, naranjas (Citrus sinensis), mandarinas (C. reticulata), limas (C. aurantifolia), limones (C. limon), toronjas (C. paradisi) y otras Citrus spp. [Snowdon, 1990; Kotzé, 2000; EPPO, 2003]. Dos tipos de esporas pueden diseminar el hongo: las asexuales (conidios), que se desarrollan en los frutos y hojas en la planta, y las sexuales (ascosporas), que lo hacen en las hojas en descomposición.
La enfermedad constituye un problema para la producción en América del Sur, África, Australia y más recientemente en Cuba [Vázquez, 2007] y en Estados Unidos [NAPPO, 2010]. Los frutos afectados con MNC tienen un mal aspecto y son inservibles para el mercado de frutos frescos. La enfermedad causa además la reducción del tamaño y su caída prematura.
Los síntomas de MNC son de apariencia variable. El síntoma típico es la llamada mancha dura, constituida por lesiones superficiales de color negro con un centro gris y un anillo pardo oscuro de 3-10 mm de diámetro, que aparecen cuando los frutos comienzan a tornarse amarillentos. Frecuentemente, pero no siempre, se desarrollan los picnidios en su centro (Phyllosticta citricarpa). Cuando los síntomas de mancha dura coalescen se forma la mancha virulenta que puede tener también picnidios en su interior. Este no es el caso cuando aparecen las lesiones llamadas pecas o falsa melanosis, que consisten en manchas pequeñas (1-3 mm de diámetro), ligeramente deprimidas que pueden ser pardo claras, grises, pardas o no coloreadas en absoluto, y generalmente carecen de la presencia de picnidios. Estas pueden ser confundidas con los causados por otros patógenos de los cítricos [Kotzé, 2000].
La especie Guignardia mangiferae A. J. Roy (anamorfo Phyllosticta capitalensis P. Hennings) es un endofítico asociado a pequeñas lesiones puntuales no fácilmente distinguibles en frutos cítricos, que frecuentemente se confunde con G. citricarpa debido a su apariencia similar en las primeras fases de su desarrollo [McOnie, 1964; Baayen et al., 2002]. Esta especie está notificada como un organismo cosmopolita endofítico frecuentemente aislado de un amplio rango de plantas, incluidas Citrus sp., miembros de las familias Anacardiaceae, Theaceae [Baayen et al., 2002] y Ericaceae [Okane et al., 2001]. G. mangiferae es sinónimo de G. psidii de guayabas y de G. endophyllicola, así como P. capitalensis es sinónimo de P. anacardiacearum y de P. theacearum [Baayen et al., 2002; EPPO, 2003].
El diagnóstico de diferentes especies de hongos puede llevarse a cabo mediante criterios morfológicos (morfología y tamaño de cuerpos fructíferos, esporas y crecimiento de las colonias en diferentes medios de cultivo), biológicos (homo o heterotalismo) y moleculares (secuencia de los espaciadores internos transcritos del ADN ribosomal (ITS).
Los objetivos del presente estudio fueron caracterizar la variabilidad cultural, morfológica y fisiológica de los aislados de Guignardia spp. que concurren en frutos cítricos en Cuba, y establecer una metodología para el diagnóstico morfológico y biológico, y la diferenciación de las especies de Guignardia presentes en los frutos cítricos en Cuba.
Los objetivos del presente estudio fueron caracterizar la variabilidad cultural, morfológica y fisiológica de los aislados de Guignardia spp. que concurren en frutos cítricos en Cuba con vistas a permitir su identificación en muestras de cítricos.
MATERIALES Y MÉTODOS
Se obtuvieron y conservaron en el Laboratorio Central de Cuarentena Vegetal 39 aislamientos fungosos, de los cuales 35 se aislaron de frutos de Citrus sinensis y Citrus paradisi de diferentes localidades de Cuba, a partir del síntoma típico conocido como mancha negra o dura (considerados patogénicos) (Fig. 1), y cuatro de frutos de C. sinensis con pequeñas lesiones como puntos negros brillantes visibles al microscopio estereoscópico considerados no patogénicos, porque no desarrollan ninguno de los síntomas de mancha negra de los cítricos o cualquier patología que conduzca al rechazo de frutos comercializables por manchas o pudriciones (Fig. 2). Las muestras con síntomas se lavaron con abundante agua y detergente, se desinfectaron con hipoclorito de sodio al 1% durante 3 min, se lavaron sucesivamente tres veces con abundante agua estéril y finalmente se secaron con papel de filtro. Se tomaron asépticamente fragmentos de aproximadamente 1 mm que incluyeran la zona límite entre el tejido enfermo y el sano, y se sembraron en agar papa dextrosa (PDA) a razón de cuatro fragmentos por placas. Las placas se incubaron a 28o C en oscuridad continua.
A partir de los tres días se realizaron observaciones para detectar el desarrollo de colonias, que por su aspecto pudieran corresponderse con Phyllosticta sp. (Tabla 1). Después de los aislamientos puros se obtuvieron cultivos monoconidiales o monoascospóricos y se transfirieron a cuñas de PDA para realizar el resto de los experimentos.
Los 39 aislamientos obtenidos se sembraron en PDA y se incubaron a 28o C en oscuridad continua. Las colonias resultantes se observaron diariamente para detectar la formación y posterior maduración de los picnidios y peritecios. Se tomaron con agujas y midieron 50 cuerpos de cada aislado. Posteriormente se cortaron en secciones finas con una aguja y se tiñeron con azul de algodón. Estos cortes se montaron en portaobjetos con lactofenol y se describieron las características morfológicas observadas al microscopio óptico, según los criterios taxonómicos descritos por Sutton (1980). Se midieron de cada aislamiento 100 conidióforos y conidios (largo por ancho) cuando se formaron conidiomas (picnidios) y 100 ascas, y ascosporas cuando las estructuras formadas fueron peritecios. Los valores obtenidos fueron sometidos a análisis de varianza y las medias comparadas mediante la dócima de Tukey con un nivel de significación del 5%.


Catorce aislados representativos de la totalidad de la colección se sembraron en PDA y se incubaron durante 10 días. De las colonias obtenidas se tomaron discos de 0,5 mm, los cuales se transfirieron a placas con PDA, agar avena (AA) y agar jugo V8 (V8), y se incubaron a 28o C a oscuridad continua (un disco por placa y cinco placas por tratamiento). A los siete días se evaluaron las características culturales de las colonias en cada medio, que fueron color, forma de los bordes, el tipo de micelio, la producción de pigmentos y su aspecto general, según los criterios de Gams et al. (1988). Se determinó el diámetro de las colonias en los tres medios a los siete, catorce y veintiún días, y el crecimiento en PDA a los siete días para la totalidad de los aislados de la colección. Los valores obtenidos fueron sometidos a análisis de varianza simple, y las medias comparadas mediante un test de significación de Tukey al 5% de probabilidad de error.
Se realizó un análisis de componentes principales para determinar las características que más aportaron a la variabilidad del conjunto de los aislamientos obtenidos, donde se incluyeron los parámetros color y bordes de las colonias, producción de pigmentos solubles, el crecimiento en PDA a los siete días y el tamaño de los conidiomas. Posteriormente se efectuó un análisis de agrupamiento para determinar la existencia de grupos y el grado de similitud entre las cepas. Con las variables tamaño del conidioma, crecimiento en PDA a los siete días, bordes y color de las colonias y liberación de pigmento, se realizó un análisis de componentes principales para determinar qué variable aportaba más a la variabilidad de los aislados. Se analizaron los parámetros color de las colonias (negro claro, 1; negro intenso, 2; verde olivo, 3); bordes de la colonia (regulares, 1; +/_ regulares, 2; irregulares, 3); tamaño del conidioma; crecimiento en PDA a los siete días. Para todos los análisis estadísticos se utilizó el programa MINITAB para Windows versión 4.2.
RESULTADOS Y DISCUSIÓN
Todos los aislados produjeron conidiomas (picnidios). En los provenientes de síntomas típicos de MN comenzaron a madurar a partir de los siete a ocho días (Fig. 3). En las Tablas 1 y 2 se muestran las dimensiones de los picnidios, los conidióforos y conidios. En solo uno de los aislados no patogénicos (MMCF1) fue posible observar la maduración de escasos picnidios. En los tres restantes se formaron tardíamente y nunca maduraron.
Las dimensiones de las estructuras de los aislados patogénicos y no patogénicos estuvieron comprendidas en el rango descrito para G. citricarpa (P. citricarpa) y G. mangiferae (P. capitalense), según Sutton y Watterston (1966) y Van der Aa (1973), respectivamente, y no se detectaron diferencias entre ellas para la mayoría de las estructuras estudiadas, con excepción del tamaño de los conidiomas de los aislados no patogénicos, que fue siempre mayor de 145 µm, lo que constituye una diferencia morfológica entre ambos grupos de aislamientos.
Los conidióforos siempre fueron menores de 12 µm. Los conidióforos de la cepa EHA3 (5,88 µm) fueron significativamente menores que los del resto de los aislamientos. El ancho de los conidios fue menor en la mayoría de las cepas al descrito en la literatura para ambas especies, con excepción de las cepas EHB1, EHB7, PT y MMF1.
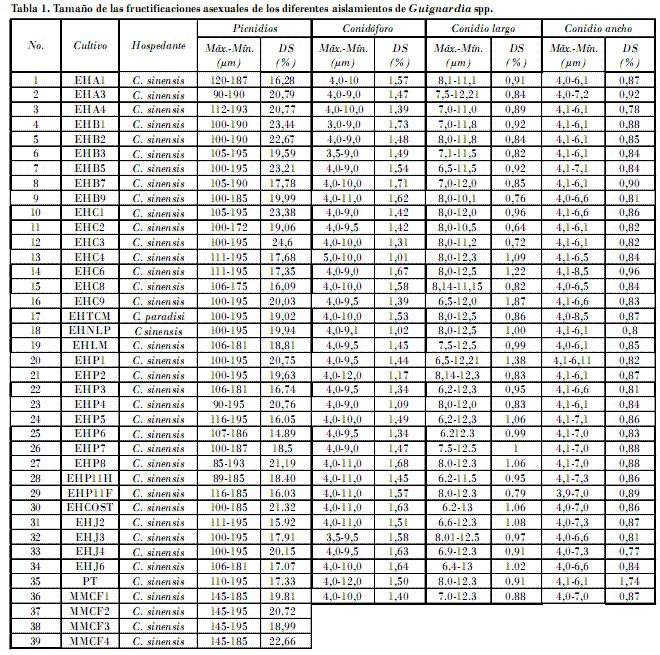
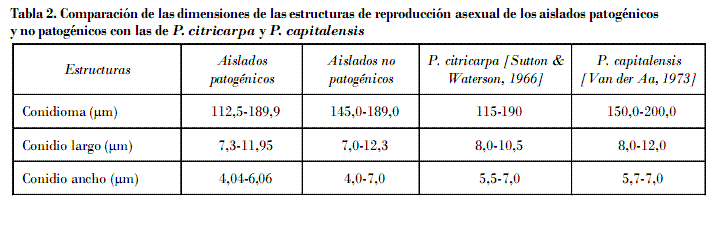
Las estructuras de reproducción asexual mostraron características morfológicas similares en los aislados patogénicos y no patogénicos, como picnidios globosos, inmersos, de color café oscuro a negro, paredes de hasta cuatro células de grosor, ostiolo más oscuro, levemente papilado y circular (Figs. 3 y 4); las picnidiosporas fueron de ovoides a piriformes, hialinas, aseptadas, multigotuladas, con ápice levemente plano y un apéndice incoloro, que termina en una punta aguda y fina, y base truncada; se formaron como blastosporas desde conidioforos hialinos, unicelulares y cilíndricos (Fig. 5).
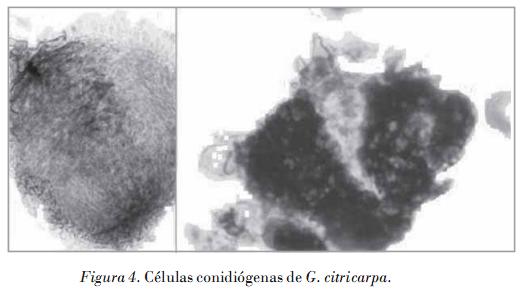
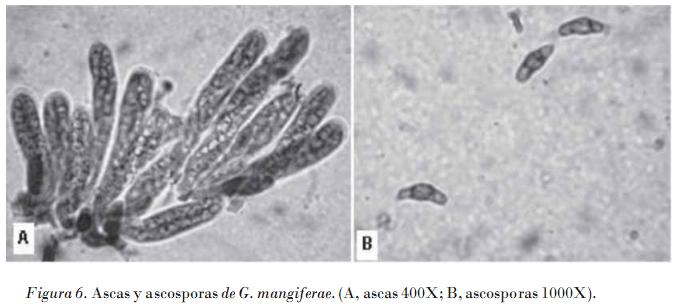
Los aislamientos monospóricos no patogénicos siempre produjeron peritecios en medio de cultivo, los cuales maduraron con facilidad a partir de los 10 días, lo que nunca ocurrió en los aislados patogénicos. Los peritecios son de piriformes a globosos, de color café oscuro a negro, con ostiolo papilado y circular; ascas de cilíndricas a clavadas, con ocho esporas; las ascosporas son aseptadas, hialinas, multigutuladas, cilíndricas pero hinchadas en el centro, con extremos obtusos, cada uno con un ápice incoloro.
Las dimensiones de las estructuras sexuales de los cuatro aislamientos no patogénicos (Tabla 3) estuvieron en el rango de los valores descritos en la literatura para G. citricarpa y G. mangiferae, lo que no permite su diferenciación. Se detectaron diferencias significativas en las dimensiones de los peritecios de los aislamientos, lo que indica una gran variabilidad entre los aislamientos de esta especie en este sentido; sin embargo, no se detectaron diferencias significativas con relación al tamaño de las ascas y ascosporas de las cuatro cepas.
En la Tabla 4 aparecen las dimensiones de las estructuras de reproducción sexual de los aislados no patogénicos obtenidas en PDA en comparación con las descritas para G. citricarpa y G. mangiferae.



Las dimensiones y morfología de los estados teleomórficos y anamórficos de las cepas no patogénicas, así como sus hábitos de crecimiento en los diferentes medios de cultivo, concuerdan con lo informado por Roy (1967) y Baayen et al. (2002) para G. mangiferae A. J. Roy (anamorfo P. capitalensis Henn.) (Tabla 4). Esta constituye la primera evidencia de la presencia en Cuba de G. mangiferae en frutos cítricos. Las características culturales de los aislados en diferentes medios de cultivo fueron:
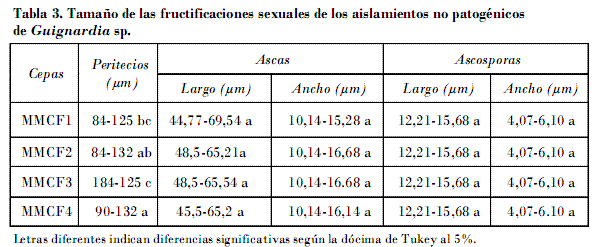
En PDA, a los siete días, el diámetro de las colonias de los aislados de G. citricarpa estuvieron entre 24,9 y 13,72 mm, significativamente inferior al de las cepas de G. mangiferae, que osciló entre 40,35 y 47,56 mm (Fig. 7). Las colonias de los aislados de G. citricarpa fueron de color gris durante los estados tempranos del crecimiento micelial, y se tornaron rápidamente negras debido a la abundante formación de picnidios que comenzaron a madurar a los siete u ocho días. Se observó un amplio halo traslúcido de micelio inmerso incoloro, que se correspondió con la zona joven de la colonia. Los bordes fueron más o menos regulares. Las cepas P5, EHP8, EHC4 y EHB7 mostraron bordes algo lobulados. El carácter borde de las colonias no constituye un criterio confiable para diferenciar ambos grupos. Aunque en la mayoría de las cepas los bordes son enteros, puede haber variaciones como de hecho se detectó. Raramente se formaron peritecios que siempre permanecieron estériles. En todos los casos se observó la abundante formación de la fase espermacial Leptodothiorella (Fig. 8).

En PDA los aislados de G. mangiferae formaron colonias de color gris a gris verdoso en los primeros días; pero rápidamente evolucionaron hacia un negro intenso con un pequeño halo traslúcido y bordes muy irregulares. Produjeron abundantemente peritecios que maduraron a partir de los 10 días, y muy raramente picnidios que solo en una ocasión maduraron (aislamiento HECF1). La fase espermacial Leptodothiorella se observó en aisladas ocasiones.
En medio AA las colonias de los aislados de G. citricarpa eran inicialmente de color amarillo mostaza, y luego se tornaron verde olivo oscuro, con bordes muy irregulares, micelio aéreo escaso restringido al centro de la colonia, con el resto del micelio inmerso y poco desarrollo de conidiomas que no maduraron hasta los 21 días. Las colonias de los aislados no patogénicos eran de color verde olivo oscuro y aclararon hacia los bordes en forma de círculos concéntricos hasta llegar a un verde grisáceo, lo que corresponde a la zona joven de la colonia. Los bordes eran lobulados con un diseño muy característico (zonado) y con halo amarillo.
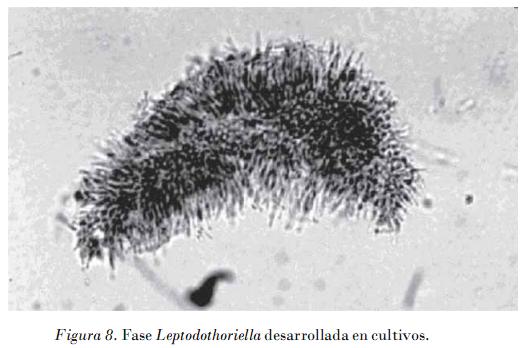
En medio V8 las colonias de los aislamientos de G. citricarpa fueron de color más claro hacia el exterior, con formación abundante de picnidios, los bordes regulares y definidos y el micelio raso. A partir de los bordes se pudo observar la liberación de un pigmento amarillo difusible que se evidencia por la formación de un halo de este color alrededor de la colonia, el cual fue uniformemente presente en estos aislamientos (Fig. 9). Los aislados no patogénicos en este medio formaron colonias pequeñas con abundante micelio aéreo algodonoso de color gris pajizo, con bordes irregulares y poco definidos. No se observó formación de pigmentos.
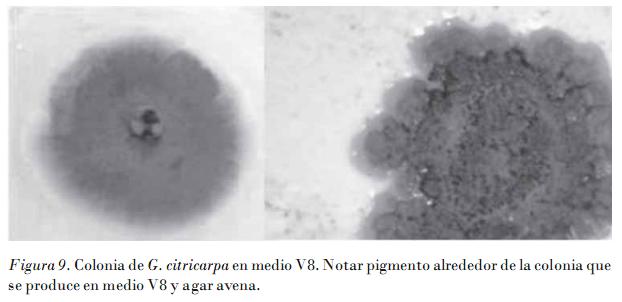
En la Fig. 10 se muestra el crecimiento a los siete días de incubación en PDA de todos los aislamientos de la colección. Los aislamientos no patogénicos son de crecimiento rápido, y las colonias alcanzan a los siete días un diámetro superior a los 40 mm, mientras que los aislamientos patogénicos no alcanzan los 30. Este comportamiento se aprecia en los tres medios de cultivo estudiados, pero es más evidente en PDA y AA.
En la Fig. 11 se muestra el crecimiento en PDA, AA y V8 a los siete días de incubación de una selección de 10 aislamientos de G. citricarpa, y los cuatro no patogénicos de G. mangiferae. En AA se observa un comportamiento del crecimiento similar al obtenido en PDA. En V8, sin embargo, no existen diferencias apreciables de velocidad de crecimiento entre los aislamientos patogénicos y no patogénicos que permita una diferenciación segura de ambos hongos.

En la Tabla 5 aparece el resultado del análisis de componentes principales basado en el tamaño de los conidiomas, el crecimiento en PDA a los siete días y el color de las colonias.

Las variables quedaron divididas en dos componentes principales que explican el 74,2 y el 25,4% de la variabilidad total, respectivamente, y juntas el 99,6%. Dentro de la primera componente principal, el crecimiento en PDA es la variable más importante, y en la segunda el tamaño del conidioma; por tanto, estas son las características que más aportan a la variabilidad de los aislados.
Se realizó un análisis de agrupamiento para determinar la existencia o no de estos grupos, y en caso positivo determinar el grado de similitud entre ambos. Fueron utilizadas las variables tamaño del conidioma y crecimiento en PDA a los siete días, porque fueron las variables que más aportaron a la variabilidad total.

Los aislamientos se dividieron en dos grupos. Uno primero (I) formado por 34 cepas de G. citricarpa aisladas a partir de los diferentes síntomas de la mancha negra de los cítricos, y un segundo grupo (II) por las cuatro cepas de G. mangiferae (aislamientos 36, 37, 38, 39) aisladas a partir de las manchas puntuales. Entre ambos grupos existe el 70% de disimilitud. Los aislamientos del grupo I (G. citricarpa) se caracterizan por tener crecimiento lento en PDA y forman abundantes conidiomas maduros más pequeños que los del grupo II (G. mangiferae). Los aislamientos del grupo II (G. mangiferae) tienen crecimiento rápido en PDA, forman peritecios maduros abundantes y escasos picnidios de un tamaño mínimo superior a los formados por G. citricarpa, y no liberan el pigmento amarillo en agar V8.
El tamaño del conidioma y el crecimiento del PDA a los siete días, junto a la producción de pigmento amarillo en medio V8 presente en G. citricarpa y ausente en G. mangiferae, así como la producción de pseudotecios en cultivos monospóricos de G. mangiferae, ausentes en G. citricarpa, permiten una clara diferenciación de ambas especies a partir de criterios morfológicos y biológicos.
La diferenciación en Cuba de los grupos de aislamientos concuerda con los informes de McOnie (1964), el cual fue el primero en dividir los aislados de Guignardia sp. en dos grupos, uno de crecimiento lento o patogénico y otro de crecimiento rápido o no patogénico. Aunque detectó la formación de escasos ascocarpos en medio de cultivo en los aislados patogénicos, nunca logró su maduración a pesar de utilizar medios de cultivo suplementado con vitaminas y extracto de hojas. Baayen et al. (2002) nunca observaron la formación de peritecios en medio de cultivo en aislamientos de G. citricarpa, y concluyeron que el hongo es heterotálico, lo que concuerda con las presentes observaciones, aspecto que debe ser verificado en el futuro a través de estudios de apareamiento con aislamientos monoascospóricos y con el empleo de marcadores moleculares. La producción de peritecios observada en los aislamientos monospóricos de G. mangiferae indica que esta especie es homotálica, lo que constituye otro carácter que diferencia a ambas especies.
CONCLUSIONES
• Existen dos especies de Guignardia en frutos cítricos en Cuba: Guignardia citricarpa Kiely y Guignardia mangiferae A. J. Roy.
• G. citricarpa se caracteriza por tener crecimiento lento en PDA, producir picnidios maduros en medio de cultivo cuyas dimensiones son frecuentemente menores de 145 µm, liberar un pigmento amarillo en agar V8 y AA, y no producir peritecios a partir de aislamientos monospóricos.
• Por primera vez se documenta la presencia en Cuba de Guignardia mangiferae, la cual no está asociada a manchas de importancia económica en frutos cítricos. Se caracteriza por tener crecimiento rápido en PDA, formar abundantes peritecios fértiles en medio de cultivo, escasos picnidios, cuyas dimensiones son frecuentemente mayores de 145 µm y no libera pigmento amarillo en agar V8.
• Ambas especies de Guignardia en cítricos pueden ser diagnosticadas mediante los criterios morfológicos y biológicos anteriormente descritos.
AGRADECIMIENTOS
Se agradece a los colegas ingeniero Leandro Herrera y a la máster María Pueyo por la contribución con aislamientos de G. citricarpa obtenidos en el LCCV; a la licenciada Ángela Porras por su apoyo y consulta en el procesamiento estadístico de los datos; al licenciado Einar Martínez por su contribución en la obtención de las fotos al microscopio y en la revisión del texto.
REFERENCIAS
Baayen, R. P.; P. J. M. Bonants; G. Verkley; G. C. Carroll; H. A. Van der Aa; I. R. Brouwershaven; G. C. Schutte; W. Maccheroni; C. Glienke de Blanco; J. L. Azevedo: «Nonpathogenic Isolates of the Citrus Black Spot Fungus, Guignardia citricarpa, Identificate as a Cosmopolitan Endophyte of Woody Plants, G. mangiferae (Phyllosticta capitalensis)», Phytopathology 92:464-477, EE. UU., 2002.
EPPO: «Guignardia citricarpa. Diagnostic Protocols for Regulated Pests. PM 7/17(1)», EPPO Bulletin 33:245-247, Inglaterra, 2003.
Gams, W.; E. S. Hoekstra; A. Aptroot: Course of Mycology. Centraalbureau Voorc Shimmelcultures, 4th ed., Baarn/Delf, Holanda, 1988.
Kotzé, J. M.: Black Spot. Compendium of Citrus Diseases, 2nd. ed., APS Press, St. Paul, Minnesota, EE. UU., 2000, pp. 23-25.
McOnie, K. C.: «The Latent Occurrence in Citrus and Other Hosts of a Guignardia Easily Confused with G. citricarpa, the Black Spot Pathogen», Phytopathology 54:40-43, EE. UU., 1964.
NAPPO: «Black Spot, Citrus», Florida, EE. UU., First report», http:// www.pestalert.org/oprDetail.cfm?oprID=421 (consultado en el 2010).
Okane, I.; A. Nakagiri; I. Tad: «Identity of Guignardia sp. Inhabiting Ericaceous Plants», Can. J. Bot. 79:101-109, Canadá, 2001.
Roy, A. J.: «Some Fungi from Almora», Indian Phytopathology 20:340-341, EE. UU., 1967.
Snowdon, A. L.: «A Color Atlas of Postharvest Diseases and Disorders of Fruits and Vegetables. General Introduction and Fruits», vol. 1, Wolfe Scientific, Reino Unido, 1990.
Sutton, B. C.: The Coelomycetes. Fungi Imperfecti with Pycnidia, Acervuli and Stromata, CMI, Kew, Surrey, Inglaterra, 1980, pp. 403-405.
Sutton, B. C.; J. M. Waterston: «Guignardia citricarpa. CMI Descriptions of Plant Pathogenic Fungi and Bacteria», no. 85, CAB International, Wallington, Inglaterra, 1966.
Vázquez, H.: «Notificación de plaga: Mancha negra de los cítricos causada por el hongo Guignardia citricarpa Kiely (Phoma citricarpa Mc Alpine)», Centro Nacional de Sanidad Vegetal. Ministerio de la Agricultura, La Habana, 2007.
 Todo el contenido de esta revista, excepto dónde está identificado, está bajo una Licencia Creative Commons
Todo el contenido de esta revista, excepto dónde está identificado, está bajo una Licencia Creative Commons
