










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Habanera de Ciencias Médicas
versión On-line ISSN 1729-519X
Rev haban cienc méd vol.11 no.1 Ciudad de La Habana ene.-mar. 2012
CIENCIAS BÁSICAS BIOMÉDICAS
Universidad de Ciencias Médicas de La Habana
Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón" Departamento de Bioquímica
Kinasas y fosfatasas: el yin y el yan de la vida
Kinases and phosphatases: the life´s yin yan
Rolando A. Hernández Fernández
Especialista Segundo Grado en Bioquímica Clínica. Profesor Titular. Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón" correo electrónico bioquimica@giron.sld.cu
RESUMEN
Introducción: la existencia de tendencias opuestas en un proceso o fenómeno de la naturaleza es la fuente principal del desarrollo.
Objetivo: el principal mecanismo de control de la mayoría de los procesos biológicos al nivel molecular son las modificaciones postraduccionales y dentro de ella los ciclos de fosforilación/desfosforilación. Las proteínas al ser fosforiladas o desfosforiladas experimentan un cambio conformacional que modifica su actividad, sus propiedades, su localización celular y las interacciones con otras moléculas.
Resultados: las enzimas implicadas en este ciclo son las kinasas y las fosfatasas, cuyas acciones coordinadas y opuestas dan el soporte mecanístico necesario para el desarrollo de múltiples funciones celulares.
Conclusiones: en este trabajo se pretende demostrar, mediante el estudio de la participación de kinasas y fosfatasas en la división celular, que estas enzimas representan la expresión molecular de los contrarios, yin y yan, de la antigua Filosofía China.
Palabras clave: kinasas mitóticas, fosfatasas mitóticas, división celular, mitosis.
ABSTRACT
Introduction: opposite forces in the inside of a process are the main sources for development.
Objective: the principal mechanism to control most biological processes is the postranslational modifications of proteins. Among these the phosphorylation/ ephosphorylation cycle is the most extended in nature. Phosphorylation or dephosphorylation of proteins causes a conformational change that modifies their activities, properties, celular localization and interactions with other molecules.
Results: kinases and phosphatese are the main enzymes implied in this cycle which coordinated and opposite actions give the mechanistic support for several cellular functions.
Conclusions: by mean of an study about the involvement of kinases and phosphatases in cell division, this paper tray to show that these enzymes represent the molecular expression of the yin and yan, the opposite element on ancient Chinese philosophy.
Key words: mitotic kinases, mitotic phosphatases, cell division, mitosis.
INTRODUCCIÓN
La presencia en un mismo objeto o fenómeno de dos tendencias opuestas, cuyas interacciones (lucha) es el mecanismo fundamental del desarrollo, aparece por primera vez en la antigua Filosofía China. Estos dos aspectos contrapuestos fueron bautizados como el yin y el yan. Esta idea fue desarrollada siglos después por Heráclito de Éfeso para quien la existencia de todos los objetos era un permanente proceso de cambio determinado en última instancia por la conversión de un contrario en su opuesto.
Para Hegel, los contrarios intervienen en el desarrollo de la idea absoluta, pero Marx y Engel establecen que los contrarios son parte de los procesos y fenómenos que existen en la realidad objetiva y la lucha entre ellos constituye el principal motor para el desarrollo. En determinadas circunstancias, existe el predominio de uno de ellos, pero rebasados ciertos límites el otro se convierte en el dominante. Tal importancia le dieron a esta idea que la elevaron a la categoría de una Ley General del Desarrollo.
La vida como fenómeno de alta complejidad no escapa a las leyes del desarrollo, entre ellas la unidad y lucha de los contrarios. Los conocimientos cada vez más crecientes de los procesos vitales al nivel molecular permiten afirmar la validez de este principio. Los procesos vitales se caracterizan por la existencia de tendencias opuestas, cuyas acciones e interacciones determinan la existencia y reproducción de los seres vivos. El autor se propone en este trabajo ilustrar la unidad y lucha de contrarios expresados en dos actividades enzimáticas contrapuestas, las kinasas y las fosfatasas. La acción coordinada entre estos dos tipos de enzimas interviene tanto en el mantenimiento de la vida como en su reproducción.
DESARROLLO
La regulación y el control de las funciones celulares y el organismo tienen una importancia trascendental para el desarrollo y mantenimiento de la vida. Aspectos tan importantes como el metabolismo, el transporte de sustancias a través de las membranas, los procesos de conservación, transmisión y expresión de la información genética, y otros muchos, están todos sujetos a finos mecanismos de regulación y control. Una de las bases moleculares más importante sobre la cual descansan estos mecanismos es el ciclo de fosforilación y desfosforilación de proteínas. Este mecanismo requiere la acción concertada de dos tipos de enzimas: las proteínas kinasas que realizan la fosforilación y las fosfoproteínas fosfatasas que catalizan la desfosforilación.
El mecanismo se basa en el hecho de que las proteínas presentan propiedades diferentes cuando están fosforiladas y cuando no lo están. Así, la intensidad de su actividad, su sensibilidad a modificadores, su localización celular y las sustancias sobre las cuales actúan dependen del estado de fosforilación de la proteína. La adición del grupo fosfato a una proteína o la sustracción del mismo desde una proteína provoca en ella un fenómeno de transconformación que trae como consecuencia un cambio en sus propiedades biológicas y, por lo tanto, una modificación en las características de los procesos en los cuales participan estas proteínas. El nivel de fosforilación de una proteína dada en cualquier momento refleja las actividades relativas de las kinasas y las fosfatasas que catalizan el proceso de interconversión de las diferentes formas de la proteína.
Las proteínas kinasas
Se denominan proteínas kinasas (para este trabajo solamente kinasas) a aquellas enzimas, o sea, proteínas con actividad catalítica, que catalizan la transferencia de grupos fosforilos desde un nucleósido trifosfatado (generalmente el ATP) hacia aminoácidos hidroxilados que forman parte de una proteína. Por tanto, las kinasas pertenecen al grupo de las fosfotransferasas.
Las proteínas kinasas suelen clasificarse en tres grupos: Las que transfieren el grupo fosforilo hacia residuos de serina o treonina (seril(treonil)-proteína kinasa, S/TPK); las que lo hacen hacia residuos de tirosina (tirosil-proteína kinasa, YPK) y las terceras lo hacen hacia cualquiera de ellos (S/YPK).
Es común que estas proteínas pertenezcan a familias génicas y muchas de ellas presenten varios isoformas con características cinéticas y mecanismos de control sutilmente diferentes. Entre las SPK las dos familias más importantes son la AGC 1 (por sus miembros fundadores las proteínas kinasas A, G y C) y la familia de las kinasas de las proteínas activadas por mitógenos, MAPK (del inglés, Mitogen-Activated Proteín Kinases).2 La primera posee más de 80 miembros y la segunda, alrededor de 20 enzimas. Entre las YPK se distinguen dos tipos principales: Las que actúan como receptores de membrana (58 miembros agrupados en 20 familias) y las intracelulares (30 miembros que forman 11 familias).3
Todas las proteínas kinasas están organizadas por dominios. Al menos tres dominios integran la proteína: el dominio catalítico, en el cual se distinguen dos sitios: uno, de unión al ATP (lazo P) y el otro, que interviene en la catálisis (lazo T), que contiene un aminoácido hidroxilado, cuya fosforilación incide en el estado de actividad de la enzima; el otro dominio es el de regulación que, bien por cambios conformacionales, bien por la unión con otras moléculas, puede influir en la mayor o menor actividad de la enzima y el tercer dominio es de autoinhibición que, en la mayoría de los casos, contiene una secuencia de aminoácidos similar a la que es reconocida por la enzima durante la catálisis, pero no contiene el aminoácido hidroxilado. Este dominio se asocia con el sitio catalítico y mantiene la enzima en un estado inactivo, hasta el momento en que se produce la activación por el dominio regulador. En algunas proteínas kinasas, el dominio de autoinhibición está formando parte de otro polipétido y la proteína se presenta en forma de oligómeros inactivos, que al disociarse adquieren la actividad.
Las Fosfoproteínas fosfatasas
Las fosfoproteínas fosfatasas (en este artículo solamente fosfatasas) son enzimas que catalizan la hidrólisis del enlace éster fosfórico. Al igual que las kinasas, existen tres grandes grupos de fosfatasas: Las que catalizan la hidrólisis del enlace del fosfato a residuos de serina o treonina, (S/TPP); las que lo hacen sobre residuos de tirosina (YPP) y las que tienen especificidad dual (S/YPP).
También pertenecen a un reducido número de familias génicas. Hasta el momento se han identificado cuatro grandes familias: las PPP, las PPM (la M porque dependen de metales para su actividad), las PTP y las FCP1, de la cual solamente existe un miembro. Las dos primeras codifican S/TPP 4 mientras que la tercera lo hace para YPP y para las S/YPP 5 y la cuarta es específica para el dominio carboxilo terminal de la ARN polimerasas II. Dentro de cada familia se logra una gran diversidad estructural generada por la unión a las subunidades catalíticas de subunidades reguladoras o de dirección.
Entre las S/TPP las más estudiadas han sido la 1 (PP1) y la 2 (PP2). Las PP4, PP5, PP6 y PP7 han sido menos estudiadas y no se mencionan en el artículo. A su vez, las PPP2 se clasifican en tres subtipos atendiendo al requerimiento de cationes para su actividad. Así, la PPP2A no requiere de ningún catión, la PPP2B requiere de Ca2+ y la PPP2C requiere de Mg2+ ,esta última de la familia PPM.
La PPP1 6 está formada por una subunidad catalítica de 37 kDa, de la cual existen cuatro isoformas. La holoenzima se completa con la asociación de subunidades reguladoras que pueden inhibir la actividad enzimática, limitar la especificidad de sustrato o dirigir la subunidad catalítica a los compartimentos celulares. En humanos, se han identificado aproximadamente 180 de estas subunidades. La PPP1 es, por lo tanto, de localización en partículas.
En el caso de la PP2A 7, la subunidad catalítica (C) está formada por 309 aminoácidos para una masa molecular de 36 kDa y de ella existen dos isoformas. La subunidad reguladora general (A, también conocida como PR65) es de 65 kDa y también existen dos isoformas. Existen, al menos, 15 subunidades reguladoras específicas que se agrupan en cuatro familias de acuerdo con su peso molecular PR54, PR55, PR72, y PR110. Asimismo, existen tres formas de PR55 y dos de PR72. Las combinaciones de estas subunidades contribuyen a la gran diversidad estructural y funcional de las PPP2A, pues con ellas pueden formarse alrededor de 70 combinaciones.
No se describirán la PPP2B ni la PP2C porque no está demostrado su participación en la mitosis.
Las cuatro familias de fosfatasas están representadas en el núcleo. 8 Algunos miembros de la familia PPM son exclusivamente nucleares y, por lo tanto, poseen la señal de localización nuclear en su secuencia de aminoácidos. Aparte de la PP2B que existe en el núcleo en cantidades pequeñas y variables la concentración de los miembros de la familia PPP es de 2 a 10 veces más alta en el núcleo que en el citoplasma. Es de notar que las subunidades catalíticas de estas enzimas no poseen la secuencia de localización nuclear por lo cual es posible que sean transportadas hacia el núcleo en unión de alguna de sus subunidades reguladoras.
La importancia de estas enzimas puede evidenciarse por el hecho de que en el genoma humano existen 518 genes que codifican proteínas kinasas (90 YPK y 428 de S/TPK). Debía suponerse un número aproximadamente igual de genes codificadores de fosfatasas; sin embargo, existen 107 genes para YPP e increíblemente sólo ~40 genes para las subunidades catalíticas de las S/TPP. No obstante, el considerable número de subunidades reguladoras y sus variantes por empalme alternativo dan a este grupo un número considerable de holenzimas. De esta manera, el número de kinasas y fosfatasas es prácticamente el mismo.
EL UNO SE CONVIERTE EN EL OTRO
Como proclamara Heráclito, una de las características de los contrarios es la transformación del uno en su opuesto y de esta forma determinar el desarrollo de los procesos y fenómenos de la Naturaleza. Las kinasas y las fosfatasas también pueden hacer esta conversión, pero de una forma peculiar. Por lo general, tanto la actividad de las kinasas como de las fosfatasas están controladas mediante ciclos de fosforilación y desfosforilación. Tanto unas como otras forman parte de vías de transferencia de información que tienen enzimas de un tipo u otro como intermediarios. En el ejemplo seleccionado, se ilustrará esta idea con algún grado de detalle.
Para demostrar la tesis planteada en el título de este trabajo solamente se expondrá un proceso de sobrada importancia en la naturaleza viva, la reproducción celular.
CONTROL DE LA REPRODUCCIÓN CELULAR
La reproducción es una de las características más sobresalientes de los seres vivos. En las células eucariontes, esta se realiza mediante un complejo proceso denominado mitosis. La participación de kinasas y fosfatasas en el control de la progresión de este proceso es tan prominente que pudiera dividirse la mitosis en dos grandes etapas: etapa de las kinasas que abarca desde finales de G2 hasta la metafase y la etapa de las fosfatasas que comprende desde el inicio de la anafase hasta el comienzo de la fase G1. Sin embargo, kinasas y fosfatasas están unidas y opuestas durante todo el proceso dando un magnífico ejemplo de la tesis que se pretende demostrar en este trabajo.
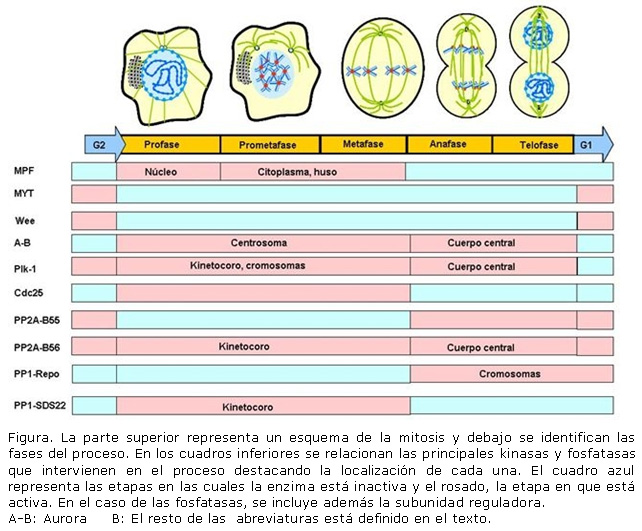
Aunque el número de ambos tipos de enzimas que intervienen en el proceso es impreciso, las principales kinasas mitóticas son la kinasa 1 dependiente de ciclina, Cdk1 (del inglés, Cyclin-Dependent Kinase),9 las kinasas de la familia Aurora (Aurora A, B y C) 10 y la familia de las kinasas poloides, Plk (del inglés, Polo-Like Kinases) 11 (Plk1 a Plk4). Por su parte las principales fosfatasas son las PP1, PP2A, y la Cdc25 que es de especificidad dual.12 A continuación, se hará una breve descripción de los eventos moleculares que garantizan la progresión de la mitosis. La exposición se hará en forma simplificada para justificar la tesis del trabajo. Una exposición exhaustiva de los mecanismos moleculares de la mitosis sobrepasa el alcance de este artículo. La participación de kinasas y fosfatasas en la mitosis se resume en la Figura 1.
Eventos previos a la mitosis
Durante la fase S en el núcleo se produce la duplicación del ADN y las dos moléculas resultantes del proceso se mantienen unidas mediante un complejo multiproteínico denominado cohesinas que se va depositando y forma un anillo alrededor de las nuevas moléculas, en la misma medida en que se van integrando. Por otra parte, en el citoplasma se realiza la duplicación de los centríolos que es un evento que depende de la actividad de la Aurora A. También depende de esta kinasa el reclutamiento hacia ese organelo de varias proteínas entre ellas la U-tubulina. Aunque el papel preciso de esta kinasa en el proceso no está totalmente dilucidado su participación en el mismo está demostrada.13
También a mediados de esta fase, comienza a incrementarse la concentración de la Plk1. Durante la fase G2, Plk1 fosforila a SA2 (un componente del complejo de las cohesinas) promoviendo la apertura del anillo y la separación de las moléculas de ADN. Sin embargo, esta acción es contrarrestada por la PP1 que desfosforila a SA2. Al principio predomina la acción de PP1, pero en la medida en que aumenta la concentración de Plk1 la actividad de esta se impone y las cohesinas son fosforiladas y el anillo se abre. Esta oposición entre la kinasa y la fosfatasa previene la separación prematura de las moléculas de ADN y la formación de las cromátidas. Solamente permanecen las cohesinas localizadas alrededor del centrómero, pues la proteína Shogusin, SGO, asociada al centrómero, recluta hacia ese sitio a la PP2A (con la subunidad reguladora B56) que contrarresta la acción de Plk1.
Estas acciones contrarias hacen posible que la mitosis visualizada con el microscopio óptico comience con la aparición de pares de filamentos largos y delgados unidos en un punto. Fue precisamente la presencia de estos filamentos lo que dio nombre al proceso, pues mitosis proviene del griego mitos que significa hilos.
Etapa de las kinasas
Al nivel molecular, el evento clave para el comienzo de la mitosis es la activación del factor promotor de la mitosis, MPF (del inglés, M-phase Promoting Factor) compuesto por la Cdk1, y la ciclina B. La Cdk1 se forma constitutivamente durante todo el ciclo celular, pero la ciclina B solamente comienza a sintetizarse a mediados de la fase S y se asocia con la Cdk1. Sin embargo el complejo permanece inactivo, debido a la actividad de las kinasas MYT1 (del inglés, Membrane-associated Tyr/Thr kinase) y Wee (porque mutaciones en el gen producen organismos de pequeño tamaño) que fosforilan a CDK1 en la Treonina 14 (T-14) y la tirosina 15 (Y-15) respectivamente inhibiendo su actividad. La activación del MPF depende además de la unión a la ciclina B de la desfosforilación en T-14 y Y-15 y la fosforilación en T-160.9
La fosforilación en T-160 que incrementa la actividad de la Cdk1 es catalizada por la kinasa activadora de Cdk, CAK (del inglés, Cdk Activating Kinase) que está formada por la subunidad catalítica Cdk7 y la ciclina H. Esta reacción puede ocurrir antes o después de las desfosforilaciones de T-14 y Y-15.
La desfosforilación de la Cdk1 es catalizada por la fosfatasa de especificidad dual Cdc25, de las cuales en los humanos existen tres isoformas, Cdc25A, Cdc25B y Cdc25C, siendo la Cdc25A la principal fosfatasa del inicio de la mitosis. En los momentos finales de G2, Cdc25A es fosforilada y activada por Plk1 y esto le permite la desfosforilación de la T-14 y Y-15 de la Cdk1, con lo cual el MPF adquiere su máximo grado de actividad, lo cual coincide con el inicio de la profase.11
Por otra parte, la separación de las cromátidas hace posible que la kinasa Aurora B fosforile a la histona H3 en serina-10 de la cromatina acelerando la condensación de esta que se observa durante la profase y prometafase.14
El MPF fosforila a la lámina B y nucleoporinas del complejo del poro nuclear y con estas dos acciones promueve el desensamblaje de la envoltura nuclear con lo cual finaliza la profase y da comienzo la prometafase.
Mientras tanto, en el citoplasma se produce la separación de los centríolos y comienza la formación del huso evento asociado a la fosforilación de varias proteínas por Aurora A y algunas de ellas son reclutadas hacia el extremo (+) del huso donde cumplen una función estabilizadora.
Durante la prometafase, Aurora B, es reclutada hacia los centrómeros, gracias a la fosforilación de la histona H3 en la treonina 3 (H3T3p) catalizada por la kinasa haspin. En ese lugar, la enzima constituye la subunidad catalítica del complejo cromosómico viajero formado además por la survivina, la borealina y la proteína centromérica E (CENPE) que funciona como un motor celular del kinetocoro.15
La unión de las fibras del huso al kinetocoro es controlada por la acción coordinada y opuesta de Aurora B y PP1. Proteínas del kinetocoro reclutan la PP1 (con la subunidad reguladora SDS22) hacia el kinetocoro y allí su acción contrarresta la de Aurora B tanto por la desfosforilación de sus sustratos como de la propia kinasa. Por su parte, Aurora B inhibe el reclutamiento de PP1 fosforilando CENPE en los sitios de unión de PP1. Este mutuo control puede establecer un estado dinámico de las fosforilaciones en la zona externa del kinetocoro cuando el mecanismo que genera la tensión separa la Aurora B hacia el interior y la PP1 hacia el exterior del kinetocoro, permitiendo una rápida respuesta ante errores en la unión de los cromosomas al huso.16
Simultáneamente, el MPF fosforila a las proteínas Cdc20 y Cdh1 (del inglés, Cdc20 homologue 1), que son coactivadores del complejo promotor de la anafase, APC (del inglés, Anaphase Promoting Complex) así como algunas subunidades del APC. La Cdc20 fosforilada es secuestrada en el huso por el complejo de control del ensamblaje del huso, SAC (del inglés, Spindle Assembly Checkpoint) 17 y la Cdh1 en su forma fosforilada no puede unirse al APC. Las fosforilaciones del APC por el MPF son contrarrestadas por otras fosforilaciones catalizadas por la PKA que tienen un efecto inhibitorio. Cuando se establece la unión bipolar de los cromosomas se disocia el SAC y se libera la Cdc20 fosforilada que se une al APC. La PP1 elimina entonces las fosforilaciones provocadas por la PKA conduciendo a la activación total del APC.
El APC es un complejo multiproteínico, cuya función es la transferencia de ubiquitina a proteínas que serán degradas por el proteasoma.18 Las proteínas sustratos del APC están determinadas por sus coactivadores, Cdc20 y Cdh1. En el tránsito de la metafase a la anafase el APC-Cdc20 marca a la segurina, una proteína inhibidora de la separasa, que al quedar libre del efecto inhibitorio hidroliza una proteína componente del complejo de cohesinas del centrómero, permitiendo así la separación de las cromátidas impulsada por la tensión del huso y la acción de proteínas motoras. Con este evento, se da inicio a la anafase. El otro sustrato importante del APC-Cdc20 es la ciclina B. La degradación de la ciclina B produce una disminución considerable de la actividad de kinasas y un aumento relativo de la actividad de las fosfatasas.
Etapa de las fosfatasas
Una de las primeras consecuencias del incremento de la actividad de fosfatasas es la desfosforilación de Cdc20 (que se separa del APC) y Cdh1 (que se une al APC) con lo cual cambia la especificidad de sustrato del APC. Sin embargo, la ciclina B sigue siendo un sustrato de APC-Cdh1 con lo cual la actividad de la CDK1 se hace prácticamente nula.19 Al comienzo de la anafase, empieza a formarse el anillo contráctil que separará las dos células hijas al final de la mitosis. Su localización depende del cuerpo central formado por la interdigitación de los extremos + del huso hacia el centro de la célula. Hacia esa zona se traslada el complejo cromosómico viajero (de ahí su nombre) y la Plk1 que son indispensables en la formación del anillo.20 Al final de la anafase, tanto Aurora B como Plk1 son marcados por el APC-Cdh1 y degradados por el proteasoma. Así, al inicio de la telofase ha desaparecido totalmente la actividad de las kinasas mitóticas.
La telofase se caracteriza por el predominio absoluto de la actividad de las fosfatasas.21 La PP2A con la subunidad reguladora específica B55á elimina las fosforilaciones realizadas por el MPF 22 en la lámina B y las nucleoporinas, y facilita de esta forma la reorganización de la envoltura nuclear.23 La PP1 con su subunidad reguladora PNUTS (Phosphatase 1 Nuclear Targeting Subunit) revierte las fosforilaciones de la cromatina catalizadas por Aurora B en G2 y contribuye a la descondensación de la cromatina.24
Simultáneamente, con las descondensación de la cromatina se produce la citokinesis, la separación de las dos células hijas y con ello el fin de la mitosis.
CONCLUSIONES
La existencia de tendencias opuestas en la génesis del desarrollo de los procesos en la Naturaleza es una Ley General del Desarrollo. Las kinasas y fosfatasas representan actividades enzimáticas opuestas, pues la una hace lo contrario de la otra. Aún más, en lo que a mecanismos de regulación se refiere, la existencia de una determina la existencia de la otra y ambas se oponen y colaboran. En ocasiones, la actividad de una predomina, pero es sustituida en el momento adecuado por la actividad de la otra. La mitosis, que es el ejemplo seleccionado, muestra cómo la acción coordinada y contrapuesta de estas actividades dirigen el proceso hacia su culminación.
Si el yin y el yan de los antiguos chinos son la fuente del desarrollo, en los fenómenos moleculares de la vida ellos pueden ser expresados como la unidad y la oposición de las actividades de kinasas y fosfatasas.
OBSERVACIÓN. (Por las características de este trabajo las referencias no son para ubicar datos primarios, sino para ampliar los conceptos y procesos mencionados en el artículo).
REFERENCIAS BIBLIOGRÁFICAS
1. Pearce lR, Komander D. y Alessi DR. The nuts and bolts of AGC protein Kinases. Nature Rev Mol Cell Biol. 2010; 11: 9-22.
2. Rose BA, Force T. y WangY. Mitogen-Activated Protein Kinase Signaling in the Heart: Angels Versus Demons in a Heart-Breaking Tale. Physiol Rev. 2010; 90: 1507-1546.
3. Östman A, Hellberg C. y Böhmer FD. Protein-tyrosine phosphatases and cancer. Nature Rev Cancer. 2006; 6: 307-320.
4. Shi Y. Serine/Threonine Phosphatases: Mechanism through Structure. Cell. 2009; 139: 468-484.
5. Patterson KI, Brummer T, O'Brien PM. y Daly RJ. Dual-specificity phosphatases: critical regulators with diverse cellular targets. Biochem. J. 2009; 418: 475-489.
6. Bollen M, Peti W, Ragusa, MJ y Beullens M. The extended PP1 toolkit: designed to create specificity. Trends Biochem Sc. 2010; 35: 450-458.
7. Janssens V, Longin S. y Goris J. PP2A holoenzyme assembly: in cauda venenum (the sting is in the tail). Trends Biochem Sc. 2008; 33: 113-121.
8. Moorhead GBG, Trinkle-Mulcahy L. y Ulke-Lemée A. Emerging roles of nuclear protein phosphatases. Nature Rev Mol Cell Biol. 2007; 8: 234-244.
9. Morgan DO. Cyclin-dependent Kinases: engines, Clocks, and Microprocessors. Ann Rev Cell Dev Biol. 1997; 13: 261-291.
10. Carmena, M., Ruchaud, S. y Earnshaw, W. C.: Making the Auroras glow: regulation of Aurora A and B kinase function by interacting proteins. Curr Opin Cell Biol. 2009; 21: 796-805.
11. Takaki T, Trenz K, Costanzo V. y Petronczki M. Polo-like kinase 1 reaches beyond mitosis-cytokinesis, DNA damage response, and development. Curr Opin Cell Biol. 2008; 20: 650-660.
12. Boutros R, Dozier C. y Ducommun B. The when and wheres of CDC25 phosphatases. Curr Opin Cell Biol. 2006; 18:185-191.
13. Morris CA y Moazed D. Centromere Assembly and Propagation. Cell. 2007; 128: 647-650.
14. Walczak CE, Cai S. y Khodjakov A. Mechanisms of chromosome behaviour during mitosis. Nature Rev Mol Cell Biol. 2010; 11: 91-102.
15. Ruchaud S, Carmena M. y Earnshaw WC. The Chromosomal Passenger Complex: One for All and All for One. Cell. 2007; 131: 230-231.
16. McIntosh JR, Grishchuk EL. y West RR. Chromosome-Microtubule Interactions During Mitosis. Annu. Rev. Cell Dev. Biol. 2002; 18:193-21.
17. Musacchio A. y Salmon ED. The spindle-assembly checkpoint in space and time. Nature Rev Mol Cell Biol. 2007; 8: 379-393.
18. Pines J. Cubism and the cell cycle: the many faces of the APC/C. Nature Rev Mol Cell Biol. 2011; 12: 427-438.
19. Wurzenberger C. y Gerlich DW. Phosphatases: providing safe passage through mitotic exit. Nature Rev Mol Cell Biol. 2011; 12: 469-482.
20. Barr FA. y Gruneberg U. Cytokinesis: Placing and Making the Final Cut. Cell. 2007; 131: 847-860.
21. De Wulf, P. Montani, F. y Visintin, R.: Protein phosphatases take the mitotic stage. Curr Opin Cell Biol 2009; 21: 806-815.
22. Queralt E. y Uhlmann F. Cdk-counteracting phosphatases unlock mitotic exit. Curr Opin Cell Biol. 2008; 20: 661-668.
23. Kutay U. y Hetzer MW. Reorganization of the nuclear envelope during open mitosis. Curr Opin Cell Biol. 2008; 20: 669-677.
24. Sullivan M. y Morgan DO. Finishing mitosis, one step at a time. Nature Rev Mol. Cell Biol. 2007; 8: 894-903.
Recibido: 15 de enero de 2012.
Aprobado: 2 de febrero de 2012.


{kind=link}