










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Habanera de Ciencias Médicas
versión On-line ISSN 1729-519X
Rev haban cienc méd vol.12 no.1 Ciudad de La Habana ene.-mar. 2013
Universidad de Ciencias Médicas de La Habana. Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón". Departamento de Bioquímica
La vía ubiquitina-proteasoma ¿destruir o construir? ese es el dilema
Ubiquitin-proteasome pathway to built or to destroy? that is the question
Rolando A. Hernández Fernández
Profesor Titular. Especialista de Segundo Grado en Bioquímica. Universidad de Ciencias Médicas de La Habana. Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón". Departamento de Bioquímica. rolher@giron.sld.cu, lantigua@infomed.sld.cu
RESUMEN
Introducción: la construcción de nuevas células es un proceso complejo y para lograr que sea unidireccional e irreversible la célula utiliza el mecanismo de destruir proteínas que se oponen al paso de una etapa a la siguiente.
Objetivo: demostrar que la destrucción de proteínas contribuye a la reproducción celular.
Método: se analizaron más de 500 artículos de los últimos 5 años publicados en revistas nacionales y de circulación internacional, disponibles en las bases de datos HINARI, PubMed y Perii y localizados mediante el sitio infomed.
Desarrollo: primero, se hizo una exposición sobre la vía ubiquitina-proteasoma. Después, se analizó la participación en el ciclo celular de los dos grandes complejos con actividad de ubiquitina-ligasa que son los encargados de marcar las proteínas que deben ser destruidas. Estos complejos actúan consecutiva y coordinadamente, y sus acciones determinan el progreso en un solo sentido del ciclo de vida de la célula.
Conclusiones: la destrucción selectiva de proteínas mediante la vía ubiquitina proteasoma permite la formación de nuevas células y con ello la perpetuación de la vida.
Palabras clave: ubiquitina, ubiquitina-ligasa, complejo SCF, complejo APC, ciclo celular.
ABSTRACT
Introduction: the generation of new cells is an extremely complex process. To become this process unidirectional and irreversible cells destroy proteins which actions are oppose to the transition from one phase to the next one.
Objective: to show that protein destruction is essential for cell reproduction.
Material and Methods: more than 500 papers published during the last five years in national and international journals were analyzed. These articles are available in HINARI, PubMed, and Perii databases and were localized through infomed.
Development: first, the ubiquitin-proteasome pathway is presented. Next, the contribution of the complexes SCF an anaphase promoting complex to the progression of the cell cycle is analysed. These complexes act consecutively and coordinately and their actions determine de progression of the cell´s life.
Conclusions: the selective destroy of specific proteins by mean of ubiquitin-proteasome pathway, allow new cells formation, and ensure the continuity of life.
Key words: ubiquitin, ubiquitin-ligase, SCF complex, APC complex, cell cycle.
INTRODUCCIÓN
La eliminación selectiva de proteínas es uno de los mecanismos más empleados en el control de procesos celulares complejos, pues permite limitar la actividad de estas moléculas a momentos específicos de la vida celular. Es la versión molecular del refrán popular cubano "Muerto el perro, se acabó la rabia."
Aún cuando las proteínas están sujetas a un recambio constante, la vida media de cada una es diferente, encontrándose las controladoras del ciclo celular entre las de vida más efímera. En este artículo se hará primero una somera exposición de la vía ubiquitina-proteasoma por estar implicada en el control del ciclo celular para por último dar los argumentos que demuestran la tesis de este trabajo. El objetivo de esta investigación es demostrar cómo la destrucción selectiva de proteínas es parte esencial del proceso de reproducción celular.
MATERIAL Y MÉTODOS
Se analizaron más de 500 artículos de los últimos 5 años, publicados en revistas nacionales y de circulación internacional disponibles en las bases de datos HINARI, PubMed y Perii y localizados mediante el sitio www.infomed.sld.cu
DESARROLLO
La vía ubiquitina-proteasoma está relacionada con múltiples funciones celulares, tales como el control del ciclo celular, la reparación del ADN dañado, la apoptosis, la respuesta inmune, etcétera. A continuación se describe esta vía.
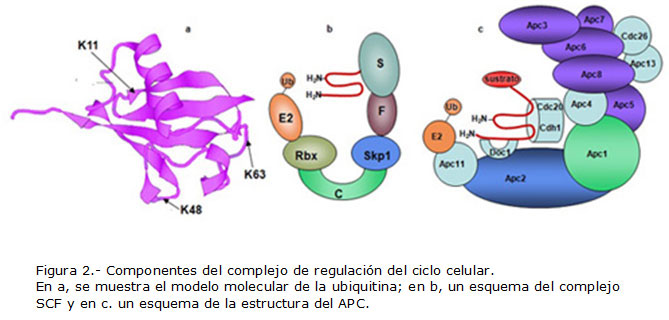
La ubiquitina. Es un polipéptido de 76 aminoácidos que se encuentra muy conservado desde las levaduras hasta el hombre. En su extremo C-terminal presenta dos glicinas consecutivas y la última se une a cadenas laterales de lisina mediante un enlace isopeptídico. También presenta varios residuos internos de lisina, de los cuales el más importante para lo que se pretende demostrar es el que ocupa la posición 48 (K48). Al grupo amino de esta lisina se puede unir una segunda molécula de ubiquitina y al de esta una tercera y así sucesivamente hasta formar una cadena de poliubiquitina. Cuatro ubiquitinas constituyen la señal para la destrucción por proteólisis. Los trabajos iniciales acerca de la ubiquitina y sus funciones celulares fueron descritos por Glickman y Ciechanover.1 Aaron Ciechanover, junto con Avram Hershko y Irwin A. Rose recibió el premio Nobel de Química en 2004 por sus trabajos en este campo.
En 1983, Hershko y sus colaboradores descubrieron tres actividades enzimáticas necesarias para la adición de ubiquitinas a proteínas sustrato en un proceso dependiente de ATP. La primera, unía la ubiquitina al centro activo de la enzima activadora de ubiquitina (E1). Esta unión requiere de ATP y se forma un enlace tioéster entre el carboxilo de la glicina última de la ubiquitina y un grupo sulfihidrilo del centro activo de la enzima. Las enzimas de este tipo son escasas de tal modo que en levaduras solamente existe una. La segunda, transfiere la ubiquitina desde la E1 hacia la enzima conjugante de ubiquitna (E2), también mediante un enlace tioéster. Al contrario de las E1, estas enzimas son más numerosas. Las E2 realizan una función central en este proceso. Investigaciones recientes apuntan que la E2 determina si el producto final contendrá mono-ubiquitina o poli-ubiquitina o la unión a una lisina específica.2,3
La tercera actividad es de ubiquitina ligasa (E3). Estas enzimas son muy numerosas y se estima que los humanos poseen alrededor de 1 000. Se distinguen dos tipos principales: la que posee motivo HECT (Homologue of E6-AP[E6-Associated Protein] Carboxy Terminal) 4 que presenta una secuencia de 350 aminoácidos, encontrada por primera vez en la región carboxilo-terminal de la proteína asociada a la E6 del papiloma virus (de ahí su nombre). En él se encuentra la cisteína conservada que actúa como aceptor de la ubiquitina desde la E2 y se transfiere al sustrato. El otro tipo presenta el motivo RING (Really Interesting New Gene) que se define por la presencia de cisteínas e histidinas conservadas que forman una estructura como de "grapa cruzada". Pueden ser simples o complejas. Las simples pueden ser monómeros u homodímeros, mientras que las complejas forman complejos multiproteínicos como el complejo promotor de la anafase (APC), el SCF y el VBC. 5 Estas enzimas transfieren la ubiquitina directamente desde la E2 hacia el sustrato. Por lo tanto, es la E3 la que determina la especificidad de sustrato, lo cual explica su elevado número.
Por último, se ha identificado una actividad que ha sido denominada E4. Estas enzimas poseen un motivo RING modificado que se nombra motivo U. Su función consiste en añadir ubiquitinas desde una E2 hacia otra ubiquitina ya unida al sustrato y, por lo tanto, formar las cadenas de poliubiquitinas que determinan la degradación del sustrato. Un resumen de este proceso se ilustra en la Figura 1.
El proteasoma. Es un complejo multiproteínico gigantesco de aproximadamente 25 MDa (26S) que posee actividad de endoproteasa (de ahí su nombre). Está compuesto por dos copias de al menos 32 subunidades diferentes muy conservadas en los eucariontes. El complejo tiene la forma de un cilindro hueco y en él se distinguen dos componentes: la partícula reguladora de 19S que ocupa los extremos del cilindro y la partícula central de 20S.
En la subunidad reguladora se distinguen dos estructuras: la base y la pestaña. La base está formada por 6 subunidades con actividad de ATPasa (Rpt1-Rpt6) pertenecientes a la superfamilia AAA+ y 3 sin actividad de ATPasa (Rpn1, Rpn2 y Rpn10). Se supone que las ATPasas forman una anillo hexagonal, común en este tipo de enzimas, que está en contacto con las subunidades á de la partícula central. La pestaña, de 400 kDa, está formada por subunidades sin actividad de ATPasa (Rpn3-9,11-12) y tiene la forma parecida a un disco que puede separarse y unirse al resto de la partícula.6
La partícula central está formada por dos anillos heptagonales de subunidades a dos de subunidades b. Las subunidades a están en la parte exterior en contacto con la partícula reguladora mientras las bocupan el centro del proteasoma.7 Las regiones N-terminal de las subunidades a ocluyen el canal central, por lo que es de suponer que funcionan como una compuerta. Tres subunidades b poseen actividad de treonin-proteasa, por lo que el proteasoma en total tiene seis. Esta combinación de centros activos produce péptidos de longitud entre 3 y 23 aminoácidos. Ambas subunidades forman una estructura en forma de cilindro irregular como se ha demostrado recientemente para el proteasoma humano.8
La proteólisis parece realizarse de la siguiente forma. La proteína sustrato debe tener al menos cuatro ubiquitinas unidas para ser reconocida. La Rpn10 se asocia con la poliubiquitina mientras las Rpn1-Rpn2 se unen a la proteína. Enzimas desubiquitinizantes separan las ubiquitinas y las subunidades con actividad de ATPasa, utilizando la energía del ATP, producen el desplegamiento de la proteína y la van haciendo pasar hacia la cámara interior de la partícula central mediante cambios de conformación de las subunidades á que obstruyen la entrada. A medida que la proteína va atravesando la cámara, se produce la hidrólisis de los enlaces peptídicos y los péptidos así formados son liberados a través de la partícula reguladora. Los péptidos formados tienen una vida media muy corta, pues son atacados rápidamente por proteasas y aminopeptidasas. Cada proteasoma procesa solamente un sustrato a la vez.
Dos complejos con actividad de ubiquitina ligasa (e3) regulan el ciclo celular
La maquinaria molecular del ciclo celular asegura que cada ronda de duplicación del ADN sea seguida de una ronda de división celular y que cada uno de estos eventos se complete antes de comenzar un nuevo ciclo. El ciclo progresa por la oscilación de la actividad de las kinasas dependientes de ciclinas que son activadas por las ciclinas y que son rápidamente inactivadas por la destrucción de las ciclinas. La función de las ubiquitinas ligasas que controla el ciclo es principalmente el marcado de las diferentes ciclinas en el momento adecuado y provoca esa oscilación en la actividad de las distintas Cdk.9
Dos complejos multiproteínicos con actividad de ubiquitina-ligasa (E3) intervienen en la regulación de la progresión del ciclo celular. Son ellos el complejo SCF y el complejo promotor de la anafase APC. La acción consecutiva e interrelacionada hace progresar el ciclo celular por sus diferentes etapas.
El complejo SCF
El complejo SCF así nombrado por tres de sus componentes, la proteína asociada a la kinasa de la fase S, Skp1 (del inglés, S-phase Kinase associated Protein 1), un miembro de la familia de las culinas (Culina-1, del inglés cull: seleccionar, separar) y otro, de la familia de proteínas con motivo F. Un componente obligado es una proteína de la familia RING que es la Rbx1 (del inglés, ring box protein).10
Las células humanas expresan siete diferentes culinas (CUL1, 2, 3, 4A, 4B, 5 y 7) y cada una sirve de núcleo estructural a ubiquitinas ligasas multiunitarias. Estas proteínas presentan una estructura extendida y rígida en forma de herradura. En este complejo, la culina 1 funciona como una plataforma molecular que interactúa simultáneamente con la proteína adaptadora Skp1 por su extremo N-terminal y con la proteína Rbx, por su extremo C-terminal y con una E2 específica, tales como Ubc3, Ubc4 o Ubc5. A su vez, Skp1 interactúa con la proteína de motivo F que es la que determina la especificidad de sustrato.
Las proteínas con motivo F, FBP (del inglés, F-box protein) se definen por la presencia del motivo de unión a proteínas conocido como motivo F que permite su unión con el resto del complejo SCF. La primera proteína con este motivo fue la ciclina F y por eso se le dio ese nombre. Posteriormente, otras proteínas fueron encontradas que interactuaban con Skp1 mediante motivos F y así nació esta familia de proteínas.11,12 Estas proteínas son las que determinan la especificidad de sustrato de los complejos SCF.
Las FBP de los mamíferos se nombran de acuerdo con las características estructurales del dominio de unión al sustrato. Este dominio ocupa invariablemente una posición C-terminal con respecto al motivo F y no se ha encontrado ningún miembro de la familia con más de un motivo F. La clase FBW (FB por F-box y W por poseer un dominio de unión a proteínas formado por repeticiones WD40). La FBL (L por presentar repeticiones ricas en leucina) con una estructura en forma de arco que aparece en otras muchas proteínas. La tercera clase está formada por proteínas que contienen un motivo diferente a WD40 y repeticiones de leucina y son denominadas FBX, las cuales forman complejos que actúan sobre otros sustratos como carbohidratos y proteínas hidroxiladas.
Los complejos SCF utilizan diferentes proteínas con motivo F para su funcionamiento, por lo cual se acostumbra a indicarla como un superíndice al mencionar el complejo. Tres son las proteínas F identificadas en los complejos SCF humanos que participan en el control del ciclo celular. Ellas son Skp2, â-TrCP (del inglés, â-Transducin repeat containing protein) y Cdc4 (Fbw7). Así cuando se quiere hacer referencia al complejo con Skp2, se escribe SCF.Skp2 En todos los casos el sustrato debe ser fosforilado para ser reconocido por las proteínas F del complejo.
La estructura cristalina del complejo SCFSkp2 muestra un bolsón cóncavo compuesto por repeticiones ricas en leucina, al cual se une el sustrato en proximidad al centro activo. SKP2 requiere además de una proteína de unión a las Cdk, Csk1, como cofactor para unir al menos algunos sustratos.
En líneas generales, el complejo puede ser descrito como formado por una plataforma en forma de herradura (culina) con la subunidad catalítica (Rbx en un extremo), unida a la E2 y la proteína adaptadora en el otro (Skp1), unida a la proteína con motivo F que es la que une el sustrato. Lo más importante es la rigidez de la zona central de la herradura que permite que los extremos con sus proteínas unidas se mantengan a una distancia constante de 59 Å que al parecer es la óptima para la catálisis.
El complejo promotor de la anafase
El complejo promotor de la anafase APC (del inglés, Anaphase-Promoting Complex), también conocido como ciclosoma, está compuesto por 15 a 17 subunidades, según la especie y muchas de ellas están muy conservadas. Es tal vez la más compleja de las ubiquitinas ligasas que se conocen, pues posee una masa molecular de aproximadamente 1 MDa.
El APC puede ser dividido en cuatro módulos: 1) catalítico, 2) de repeticiones tetratricopetídicas, TPR, 3) de plataforma y 4) activador. El módulo catalítico incluye las subunidades Apc11 una proteína de la familia RING y Apc2 de la familia de las culinas, de modo que APC y SCF pertenecen a la misma familia. El TPR interactúa con los activadores.13 La plataforma proporciona un puente entre el módulo catalítico y el TPR tal vez optimizando el espacio entre estas regiones para una catálisis eficiente. Los activadores son proteínas de la familia Cdc20 (Cdc20 y Cdh1) con repeticiones triptófano aspártico (WD) necesarios para la eficiente unión y ubiquitinación del sustrato.14
Estudios estructurales con reconstrucciones tridimensionales muestran que APC presenta una forma aproximadamente triangular con una cavidad central.15 Parece que la cavidad constituye el centro catalítico, pues Cdh1 y un sustrato modelo se unen en la cavidad.16 Esta unión se hace mediante APC10 que participa en la especificación del sustrato. Otros datos sugieren que Cdc20 se une de una forma similar.17
A diferencia del complejo SCF cuyos sustratos deben ser fosforilados previamente, el APC tiene la habilidad de reconocer diferentes sustratos no modificados en diferentes momentos del ciclo celular. Las proteínas sustratos de APC presentan una secuencia de aminoácidos, degradón, que permite su reconocimiento por los co-activadores. La mayoría de los degradón puede ser dividida en dos grupos, los motivos D (llamado así por destrucción) con la secuencia consenso RxxLxxxxN y KEN con KENxxxN. El primero es reconocido por Cdh1 en unión con APC10. Esta situación a veces se utiliza para impedir, al menos momentáneamente, la degradación de algunos sustratos, como la Cdc6 y la separasa, 18 mediante la fosforilación en, o cerca de estos motivos.
El APC utiliza dos E2 que funcionan en tándem, la UBCH5 que inicia la modificación del sustrato y la UBD2S que extiende la cadena de ubiquitinas.19 Al contrario de otras especies, el APC humano transfiere las ubiquitinas adicionales tanto a la lisina-11 (K11) 20 como a la lisina-48 (K48) 21 Se ha visto que las ubiquitinas unidas a K11 permiten la formación de poliubiquitinas más largas que son reconocidas más fácilmente por el proteasoma y degradadas con más rapidez.20 Por otra parte, las cadenas unidas a K11 permiten distinguir las proteínas marcadas por el APC de las marcadas por otras E3 y, de esta forma. influir en la regulación de las enzimas específicas que eliminan las ubiquitinas.
El hecho de que APC debe marcar un conjunto de proteínas en determinados momentos del ciclo celular podría proporcionar la razón de la complejidad de su estructura. La Figura 2 muestra los principales componentes del sistema de regulación por proteólisis selectiva del ciclo celular.
La necesidad de que cada etapa del ciclo celular se complete antes de comenzar la siguiente requiere de eficientes mecanismos de control, tanto espaciales como temporales. Durante la evolución, se han creado los llamados puntos de control que verifican el completamiento de cada etapa y permiten o no, el inicio de la siguiente. El comportamiento de sistemas complejos como el ciclo celular entraña una paradoja fundamental: un proceso cíclico presenta dificultades para producir puntos de control y un sistema que produce puntos de control tiene serias dificultades para mantener ciclos repetitivos.
Los complejos E3 que regulan el ciclo celular utilizan mecanismos diferentes para realizar su función. En unos casos, se activa la ligasa (APC), mientras que en el otro se activa el sustrato (SCF). La activación de la ligasa tiene la ventaja de poder coordinar el marcado de un conjunto de proteínas simultáneamente y ser regulado de forma muy simple, pero carece de flexibilidad temporal y situacional. La activación del sustrato permite a una sola ubiquitina ligasa satisfacer multiplicidad de necesidades del ciclo celular en momentos diferentes. En ambos casos, la activación es por fosforilación.
A los efectos de la regulación del ciclo celular es más conveniente comenzar por la mitosis. Para este estudio, hemos decidido dividir la mitosis en dos grandes etapas: la de las kinasas y la de las fosfatasas, atendiendo a la actividad enzimática predominante en cada una. La actividad de kinasas predomina desde el comienzo de la mitosis hasta la transición hacia la anafase, las fosfatasas predominan en el resto. El principal complejo regulador de una fase a la otra es APC. El papel de kinasas y fosfatasas durante el ciclo de reproducción celular ha sido examinado recientemente en un artículo publicado en esta revista.22
Entra en acción el APC
En las células humanas, el complejo Cdk1/ciclina B activo entra al núcleo ~20 minutos antes de la desaparición de la envoltura nuclear 23 y poco después puede detectarse el APC fosforilado. Se ha demostrado que Cdk1/ciclina B fosforila algunas de las subunidades del APC favoreciendo su actividad aumentando su afinidad por los co-activadores, pero también fosforila a sus dos co-activadores Cdc20 y Cdh1. Cdc20 fosforilado está listo para unirse a APC pero es secuestrado en dos complejos: el punto de control del ensamblaje del huso SAC (del inglés, Spindle Assembly Checkpoint) y el complejo de control de la mitosis MCC (del inglés, Mitosis Checkpoint Complex). Estos complejos monitorizan la unión apropiada y bipolar de los kinetoros de los cromosomas a los extremos del huso, proceso que se desarrolla durante la pro-metafase. De no producirse una unión adecuada se detiene la progresión de la mitosis.
Estudios con células vivas han demostrado que la ciclina A es el primer sustrato en ser degradado inmediatamente después de la desaparición de la envoltura nuclear. Se supone que la Cdc20 formando parte del MCC puede unir el complejo Cdk2/ciclina A con Csk1 y mediante un mecanismo desconocido hacer que APC marque a la ciclina A.24 Así a mediados de la pro-metafase comienza a disminuir la actividad de kinasas.
Cuando todos los cromosomas se han unido correctamente a las fibras del huso y se disponen en la placa ecuatorial, Cdc20 se disocia de MCC y se une al APC (de ahora en adelante APCCdc20). El complejo APCCdc20 marca con ubiquitina a la proteína segurina que es un inhibidor de la separasa, una proteasa que hidroliza los complejos de cohesinas que mantienen unidas las dos cromátidas por el centrómero. La acción de la separasa permite que las cromátidas comiencen su desplazamiento hacia los polos de las células. El otro sustrato de APCCdc20 es la ciclina B.25 Cuando la ciclina B marcada con ubiquitina es degradada por el proteasoma la actividad de kinasa se reduce considerablemente y comienza el aumento relativo de las fosfatasas.
El predominio de las fosfatasas al inicio de la anafase lleva a la desfosrilación de Cdc20 que se separa de APC y permite la unión de Cdh1 (en lo adelante APCCdh1). El APCCdh1 marca la Cdc20 y continúa marcando las ciclinas A y B con lo cual la actividad de las Cdk se reduce prácticamente a cero. Al final de la mitosis durante la citokinesis APCCdh1 marca las kinasas Aurora y poloide 1 Plk1 (del inglés, Polo-like kinase), con lo cual toda la actividad de las kinasas mitóticas es eliminada.
La actividad de APCCdh1 continúa durante la fase G1, cuando comienza la formación del complejo pre-duplicativo y para ello es necesario una baja o más bien nula actividad de kinasa. En ese momento se incrementa la concentración de Skp2 que es marcado por APCCdh1.26 También es marcada la geminina una proteína que mantiene secuestrada a Cdt1, un componente del complejo pre-duplicativo. La eliminación de la geminina permite la formación del complejo a mitad de la fase G1. En esta fase, también se encuentran elevados los niveles de los inhibidores de las Cdk, CDI, tanto los de la familia Cip/Kip como de INK4. ¿Cómo interpretar estas acciones de APCCdh1? La proteína Skp2 forma parte del complejo SCFSkp2 que marca los inhibidores de la familia Cip/Kip que mantienen inactivas a Cdk2/ciclina E. La destrucción de Skp2 mantiene activo los inhibidores y garantiza la ausencia de actividad de kinasa durante los primeros momentos de la fase G1, lo cual es necesario para la formación del complejo pre-duplicativo a lo que también contribuye APCCdh1 con la destrucción de la geminina.
Entra en acción el SCF
A mediados de G1, los mitógenos activan la síntesis de las ciclinas B que, a su vez, activan las Cdk4 y Cdk6. Al final de G1, comienza la síntesis de la ciclina E que forma complejos con la Cdk2. Como la Cdk2 está inhibida por Cip/Kip es necesario un aumento considerable de los niveles de ciclina E para desplazarlos y activar la kinasa. Un vez activo el complejo Cdk2/ciclina E, fosforila a los inhibidores, p21Cip1, p27Kip1 y p57Kip2, que son marcados por el complejo SCFSkp2. También este complejo fosforila a Cdh1 y lo hace abandonar a APC. Sin embargo, este mecanismo no parece eliminar totalmente la actividad APCCdh1. Por otra parte, una proteína de la familia F (FBX5), conocida como EMI1 (del inglés, Entry Mitotic Inhibitor), inhibe a APC por un mecanismo independiente de la proteólisis de Cdh1.27 De esta forma, queda establecida una conexión entre los dos complejos ubiquitina ligasa que regulan el ciclo celular. El APC que comienza sus funciones en la pro-metafase, se mantiene durante G1 hasta que se ha completado la formación del complejo pre-duplicativo. Entonces, deja paso a la acción del complejo SCF que conducirá la célula hasta la entrada en la mitosis como se relatará a continuación.
En los momentos iniciales de S Cdk2/ciclina E activa al complejo pre-duplicativo y se pone en marcha la duplicación del ADN. Algunos de los componentes del complejo pre-duplicativo fosforilados durante la activación son marcados por SCFSkp2, mecanismo que garantiza que la duplicación solamente puede realizarse una vez durante la vida de la célula. Además SCFâTrCP degrada la Cdc25A necesaria para la activación total de la Cdk1.28 La kinasa que fosforila a Cdc25A es desconocida. De esta manera, se mantiene alta la actividad de Cdk2 y se atenúa la de Cdk1. Unos momentos después, la propia Cdk2 fosforila a la ciclina E que es marcada por SCFCdc4 y Cdk2 se une entonces a la ciclina A. Al inicio de la mitosis EMI1 es marcado por SCFâTrCP con lo cual se suprime la inhibición sobre APC. En la Figura 3, se muestran las principales proteínas destruidas por esta vía durante cada una de las etapas del ciclo celular.
Como se puede inferir de lo expuesto, los complejos APC y SCF no sólo regulan la progresión del ciclo celular, sino que además se regulan el uno al otro. La actividad destructiva de estos dos complejos actúa coordinadamente, para entre los dos construir las nuevas células que garantizan la perpetuidad de cada estirpe celular y con ello la vida del organismo.
CONCLUSIONES
Durante la vida de la célula, es necesario que algunas proteínas solamente estén activas durante un estrecho intervalo de modo que el proceso transcurra unidireccional e irreversiblemente. La mejor forma de lograr eliminar la actividad de una proteína es mediante la proteólisis. El sistema ubiquitina-proteasoma es altamente específico, pues en respuesta a señales moleculares solamente algunas proteínas son marcadas con ubiquitina y destruidas por el proteasoma. De esta forma se selecciona cual proteína y en qué momento será eliminada. Los dos grandes complejos que marcan esas proteínas, el APC y el SCF, se alternan durante el ciclo celular. Pero no se trata de una destrucción fortuita sino que contribuye a la formación de nuevas células, las estructuras más complejas que pueden formarse a ese nivel de organización de la vida. Cada proteína realiza su función en el momento preciso e inmediatamente es marcada con ubiquitina y degradada, de manera que su acción no se prolongue más del tiempo estrictamente necesario. De esta forma no existe un verdadero conflicto entre la destrucción y la construcción, sino que ambas son facetas opuestas, pero unidas indisolublemente en la realización de una función tan compleja como la reproducción celular.
REFERENCIAS BIBLIOGRÁFICAS
1. Glickman MH, Ciechanover A. The ubiquitin-proteasome proteolytic pathway: destruction for the sake of construction. Physiol Rev. 2002; 82: 373-428.
2. Burroughs AM, Jaffee M, Iyer LM, Aravind L. Anatomy of the E2 ligase fold: implications for enzymology and evolution of ubiquitin/Ub-like protein conjugation. J Struct Biol. 2008; 162: 205-218.
3. Windheim M, Peggie M, Cohen P. Two different classes of E2 ubiquitin-conjugating enzymes are required for the mono-ubiquitination of proteins and elongation by polyubiquitin chains with a specific topology. Biochem J. 2008; 409: 723-729.
4. Rotin D, Kumar S. Physiological functions of the HECT family of ubiquitin ligases. Nat Rev Mol Cell Biol. 2009; 10:398-409.
5. Budhidarmo R, Nakatani Y, Day CL. RINGs hold the key to ubiquitin Transfer. Trends Biochem Sci. 2012; 37: 58-65.
6. Rosenzweig R, Osmulski PA, Gaczynska M, Glickman MH. The central unit within the 19S regulatory particle of the proteasome. Nature Struct Mol Biol. 2008; 15: 573-580.
7. Kusmierczyk AR, Kunjappu MJ, Funakoshi M, Hochstrasser M. A multimeric assembly factor controls the formation of alternative 20S proteasomes. Nature Struct Mol Biol. 2008; 15: 237-244.
8. Da Fonseca PCA, He J, Morris EP. Molecular Model of the Human 26S Proteasome. Mol Cell. 2012; 46: 54-66.
9. Coudreuse D, Nurse P. Driving the cell cycle with a minimal CDK control network. Nature. 2010; 468: 1074-1079.
10. Silverman JS. Skaar JR, Pagano M. SCF ubiquitin ligases in the maintenance of genome stability. Trends Biochem Sci. 2012;37: 66-73.
11. Skaar JR, Pagan JK, Pagano M. SnapShot: F Box Proteins I. Cell. 2009; 137: 1160-1160.e1.
12. Skaar JR, D'AngiolellaV, Pagan JK, Pagano M. SnapShot: F Box Proteins II. Cell. 2009; 137: 1358-1358.e1.
13. Matyskiela ME, Rodrigo-Brenni MC. Morgan DO. Mechanisms of ubiquitin transfer by the anaphase-promoting complex. J Biol. 2009; 8: 92.
14. Matyskiela ME, Morgan DO. Analysis of activator-binding sites on the APC/C supports a cooperative substrate-binding mechanism. Mol Cell. 2009; 34: 68-80.
15. Da Fonseca PC, Kong EH, Zhang Z, Schreiber A, Williams MA, Morris EP, Barford D. Structures of APC/CCdh1 with substrates identify Cdh1 and Apc10 as the Dbox co-receptor. Nature. 2011; 470: 274-278.
16. Buschhorn BA, Petzold G, Galova M, Dube P, Kraft C, Herzog F, Stark H, Peters JM. Substrate binding on the APC/C occurs between the coactivator Cdh1 and the processivity factor Doc1. Nature Struct Mol Biol. 2011; 18: 6-13.
17. Izawa D, Pines J. How APC/C_Cdc20 changes its substrate specificity in mitosis. Nature Cell Biol. 2011; 13: 223-233.
18. Holt LJ, Krutchinsky AN, Morgan DO. Positive feedback sharpens the anaphase switch. Nature. 2008; 454: 353-357.
19. Williamson A, Wickliffe KE, Mellone BG, Song L, Karpen GH, Rape M. Identification of a physiological E2 module for the human anaphase-promoting complex. Proc Natl Acad Sci USA. 2009; 106: 18213-18218.
20. Bremm A, Komander D. Emerging roles for Lys11-linked polyubiquitin in cellular regulation. Trends Biochem Sci. 2011; 36: 355-363.
21. Rodrigo-Brenni MC, Foster SA, Morgan DO. Catalysis of lysine 48specific ubiquitin chain assembly by residues in E2 and ubiquitin. Mol Cell. 2010; 39: 548-559.
22. Hernández Hernández RA. Kinasas y fosfatasas: el yin y el yan de la vida. Rev haban cienc méd [online]. 2012; 11(1): 15-24.
23. Gavet O, Pines J. Activation of Cyclin B1_Cdk1 synchronizes events in the nucleus and the cytoplasm at mitosis. J Cell Biol. 2010; 189: 247-259.
24. Di Fiore B, Pines J. How cyclin A destruction escapes the spindle assembly checkpoint. J Cell Biol. 2010; 190: 501-509.
25. Gorr IH, Boos D, Stemmann O. Mutual inhibition of separase and Cdk1 by two-step complex formation. Mol Cell. 2005; 19: 135-141.
26. Wei W, Ayad NG, Wan Y, Zhang GJ, Kirschner MW, Kaelin Jr WG. Degradation of the SCF component Skp2 in cell-cycle phase G1 by the anaphase-promoting complex. Nature. 2004; 428: 194-198.
27. Machida YJ, Dutta A. The APC/C inhibitor, Emi1, is essential for prevention of rereplication. Genes Dev. 2007; 21: 184-194.
28. Jin J, Shirogane T, Xu L, Nalepa G, Qin J. Elledge, SJ, Harper, JW. SCFâ-TRCP links Chk1 signaling to degradation of the Cdc25A protein phosphatase. Genes Dev. 2003; 17: 3062-3074.
Recibido: 3 de septiembre de 2012.
Aprobado: 20 de diciembre de 2012.

{kind=link}
{kind=link}
{kind=link}