










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Habanera de Ciencias Médicas
versión On-line ISSN 1729-519X
Rev haban cienc méd vol.13 no.1 Ciudad de La Habana ene.-feb. 2014
CIENCIAS BÁSICAS BIOMÉDICAS
Universidad de Ciencias Médicas de La Habana
Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón"
La modificación covalente de proteínas. Un nivel cualitativamente superior de la información molecular
Covalent Modifications of Proteins. A qualitative higher level of molecular information
Rolando A. Hernández Fernández
Especialista Segundo Grado en Bioquímica. Profesor Titular y Consultante. E.mail: rolher@giron.sld.cu; lantigua@infomed.sld.cu
RESUMEN
Introducción: La información molecular es una propiedad principal de las biomacromoléculas, especialmente proteínas y ácidos nucleicos, que permite la realización de funciones con un alto grado de especificidad. Ella deriva de la variedad en los componentes de las macromoléculas.
Objetivo: Demostrar que la modificación covalente de proteínas constituye un nivel nuevo y superior de información molecular.
Método: Se analizaron artículos de los últimos 5 años, publicados en revistas nacionales y de circulación internacional, disponibles en las bases de datos HINARI, PubMed y Perii y localizados mediante el sitio www.infomed.sld.cu.
Desarrollo: Se exponen los mecanismos y características principales del proceso y posteriormente algunos de los efectos principales de la modificación covalente sobre las funciones y propiedades de las proteínas.
Conclusiones: La modificación covalente es un mecanismo que amplía el campo de acción de las proteínas permitiendo un rápido cambio en sus propiedades funcionales y, por lo tanto, constituye un nivel nuevo y cualitativamente superior de información molecular.
Palabras clave: Modificación covalente de proteínas, información molecular, interacciones interproteínas, recambio de proteínas.
ABSTRACT
Introduction: Molecular information is an essential property of biomacromolecules, especially proteins and nucleic acids. This property allows carrying out specific functions. It derives from variability of macromolecules components.
Objetive: To prove that covalent modification of proteins represents a new and higher level of molecular information.
Method: Papers published during the last five years in national and international journals were analyzed. These articles are available in HINARI, PubMed, and Perii databases and were localized through www.infomed.sld.cu.
Main text: First, the mechanism and features of covalent modifications of protein are presented. Next, the principal effects on protein functions and properties are analyzed.
Conclusions: We conclude that covalent modification of proteins represents a new and higher level of molecular information.
Key words: Covalent modification of protein, molecular information, protein interactions, protein turn over.
INTRODUCCIÓN
La información molecular es la propiedad que poseen las macromoléculas biológicas que les permite actuar con un alto grado de especificidad en variados procesos con disminución de la entropía y con ello minimizar los efectos del azar. Puede ser principalmente secuencial (como en los ácidos nucleicos) o conformacional (como en las proteínas). La existencia de esta propiedad está vinculada a la variedad y al orden de los precursores que forman parte de las macromoléculas.
Mientras la forma secuencial es la idónea para conservar y transmitir la información, la conformacional es la adecuada para la realización de funciones específicas que se realizan mediante el mecanismo de reconocimiento molecular. A partir del gran número de conformaciones posibles, las proteínas realizan las más variadas funciones, incluso la formación de otras moléculas a las cuales dotan de cierto grado de información molecular.
La modificación covalente es un mecanismo muy generalizado que modula las funciones y otras propiedades de las proteínas. Este mecanismo incrementa la variedad de los precursores de una forma muy selectiva, lo cual no solo modifica la información secuencial y la conformacional, sino que aporta propiedades nuevas a esas moléculas, modifica sus interacciones con otras moléculas, determina el lugar y el momento de su actividad, etcétera. Esto significa que el grupo añadido covalentemente a las proteínas modifica su contenido informativo momentáneamente lo que permite una adaptación a determinadas condiciones en un momento determinado. Esta interpretación del fenómeno no ha sido expuesta con anterioridad y por eso el objetivo de este trabajo es demostrar que la modificación covalente de proteínas constituye un nivel nuevo y superior de información molecular.
MATERIAL Y MÉTODO
Se recopilaron artículos publicados en revistas cubanas y de circulación internacional sobre el tema, utilizando las bases de datos HINARI, PubMed y Perii disponibles en el portal www.infomed.sld.cu. Se realizó una selección de los artículos de los últimos 5 años que tienen un valor más sobresaliente en la tesis que se pretende demostrar.
DESARROLLO
Primero se hará una exposición de los principales aspectos conceptuales del problema; después, se tratará el mecanismo y las modalidades de las modificaciones covalentes de proteínas y finalmente se ilustrará con ejemplos concretos las principales consecuencias de este mecanismo.
La información molecular y la modificación covalente de proteínas
La información genética contenida en el ADN es la fuente primaria de información en los organismos celulares y se transfiere dosificadamente a las proteínas mediante los procesos de transcripción y traducción. El ADN solo puede transferir a las proteínas información secuencial, con la cual estas construyen su estructura tridimensional como fue demostrado hace años por Anfinsen. Pero la información secuencial en una determinada proteína es permanente, mientras se mantenga la integridad de la estructura primaria. Este tipo de información da lugar a la conformacional, algo más flexible, pero de todas formas es también permanente. Se puede decir que el contenido de esta información es "lo que puede hacer esa proteína", es decir, su función específica.
Ahora bien, durante la vida de las células, trátese de organismos monocelulares o de las que forman parte de los pluricelulares, las condiciones ambientales cambian, a veces con gran rapidez, y las células necesitan adaptarse a esos cambios. Un nuevo ambiente puede requerir un aumento o disminución de la actividad de determinadas proteínas, un cambio en la localización, la separación o unión de unas proteínas con otras, la síntesis o proteólisis de algunas proteínas, en fin, la supervivencia o la muerte celular. Pero en el contenido informativo, tanto secuencial como conformacional, de las proteínas no está inscrito ¿con cuál intensidad actuar?, ¿cuándo actuar?, ¿dónde actuar?, ¿con cuál molécula interactuar?, ¿hasta cuándo actuar? o ¿en cuáles situaciones actuar?
Ese tipo de información se incorpora a las proteínas mediante la adición o sustracción de grupos químicos específicos que se unen a aminoácidos específicos en posiciones definidas de la cadena polipeptídica mediante el mecanismo de modificación covalente. Resulta asombroso que la adición de un grupo tan pequeño como el hidroxilo a un aminoácido específico pueda dar como resultado un cambio tan intenso en las propiedades de una proteína. Dotar a las proteínas de ese nuevo contenido informativo que determina la intensidad, la localización, el momento, la situación, los acompañantes, etcétera de su actividad es la función esencial del mecanismo de modificación covalente.
Claro está que, todas las posibilidades enunciadas en el párrafo anterior se encuentran como posibilidad en la proteína original, pero para la supervivencia del organismo es trascendental la transición de la posibilidad a la realidad que se logra mediante el mecanismo que se analiza en este trabajo.
Como quiera que la adaptación al ambiente es un proceso de supervivencia para las células y los organismos, la modificación covalente dota a las proteínas de un contenido informacional cualitativamente superior y como esta interpretación no se ha dado anteriormente puede considerarse totalmente nueva.
Diversidad y especificidad de la modificación covalente de proteínas
La información molecular está relacionada con la variedad y la posición de precursores a lo largo del polímero. Mientras mayor sea la variedad, mayor será el contenido informativo de una macromolécula.
El número de grupos químicos reportados que pueden unirse covalentemente a las proteínas está cercano a los veinte (ver más adelante), mientras que el número de aminoácidos modificables apenas alcanzan los diez, sin tener en cuenta que en ocasiones la modificación covalente ocurre en el aminoácido N-terminal sin una gran especificidad.
Se pudiera pensar entonces en un total de 200 modificaciones teniendo en cuenta que son 20 donantes y 10 aceptores. Sin embargo, la modificación covalente no solo es específica para el grupo modificador y el aceptor, sino también para la posición de este último en el polímero. Todo lo anterior indica que la modificación covalente introduce un alto grado de diversidad en la estructura de las proteínas y con alto grado de especificidad. Añadir grupos químicos a una proteína es como añadir letras a un alfabeto, con lo cual seguramente se pueden elaborar mensajes más complejos.
Para ilustrar lo anterior, tomemos el ejemplo de la lisina. Este aminoácido puede ser monometilado, dimetilado, trimetilado, acetilado o modificado por los péptidos de la familia de la ubiquitina (al menos seis). Así, que por arte de la modificación covalente un aminoácido se convierte en 10. A esto hay que añadir la posición de la lisina en la proteína.
Por lo tanto la variación en el contenido informativo de una proteína es un fenómeno complejo y que puede dar lugar a diferentes resultados. Una relación, seguramente incompleta de los aminoácidos modificados y sus modificadores se muestra en la Tabla 1.
Uno puede preguntarse ¿cuál es la fuente de esa información? Seguramente, la información genética contenida en el ADN. Tanto la proteína sustrato como las enzimas modificadoras están codificadas en el ADN y por eso la modificación covalente puede interpretarse como un mecanismo de interacción entre genes mediante sus productos primarios. Lo cual lleva a pensar que la modificación covalente es un mecanismo codificado genéticamente. Este fenómeno se ilustra en la Figura 1.
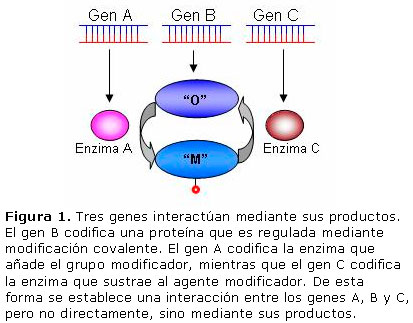
Pero un par de enzimas modificadoras puede tener numerosos sustratos, lo cual significa un ahorro considerable de material genético. De modo que la modificación covalente de proteínas puede ser considerada un ejemplo del principio de máxima economía típico de los seres vivos.
Todas las razones expuestas anteriormente nos llevan a considerar a la modificación covalente de proteínas como un nivel nuevo y cualitativamente superior de la información molecular.
Mecanismo y modalidades
La modificación covalente consiste en la adición, mediante una unión covalente, de una especie química a la cadena lateral de alguno de los aminoácidos de la proteína salida del ribosoma. Como se trata de la formación y ruptura de enlaces covalentes es necesaria la participación de una enzima para catalizar la unión y otra para la separación que en la mayoría de los casos es una hidrolasa. De este modo, se originan dos formas de la proteína, diferentes en sus propiedades y que, a los efectos de este trabajo, se denominarán la original (no modificada) y la modificada.
La adición del grupo modificador provoca una perturbación de la estructura tridimensional que puede tener mayor o menor alcance. Debido a eso, las dos formas de la proteína pueden exhibir diferencias en la intensidad de su actividad, en sus interacciones con otras moléculas, en su localización celular, en sus mecanismos de control y regulación, etcétera.
Las modificaciones pueden ser reversibles o irreversibles. En el primer caso, pueden ser únicas o múltiples y estas últimas ser de un mismo grupo modificador o de grupos diferentes. Las modificaciones múltiples tienen efectos muy variados. Primero pueden producirse efectos sinérgicos o antagónicos. Por ejemplo, las fosforilaciones en la treonina-14 (T14) y la tirosina-15 (Y15) en las kinasas dependientes de ciclinas, Cdk (del inglés, Cyclin-dependet kinase) tienen un efecto sinérgico, pues ambas son inhibidoras, mientras que la fosforilación en treonina-160 (T160) tiene un efecto antagónico con las anteriores pues es activadora. Por lo general en estos casos, hay una que tiene carácter dominante, esto es, su efecto se expresa independientemente del estado de modificación de otros grupos, tal como sucede en las Cdk que son inactivas si están fosforiladas en T14 y Y15 independientemente de si T160 está fosforilada o no.1 Este es un ejemplo de cómo la modificación covalente influye en el estado de actividad de una enzima, sin modificar su acción específica.
Otro caso de antagonismo se evidencia en el control de la transcripción por la proteína de unión al elemento de respuesta al Adenosínmonofosfato cíclico. Cuando uno de sus coactivadores, CRTC2 (del inglés Cyclic AMP-Regulated TranscriptionalCo-activators) es fosforilado en las serinas 70 y 171, esto proporciona el sitio de unión a proteínas de la familia 14-3-3 que lo mantienen retenido en el citosol. Sin embargo, la adición de N-acetilglucosamina en los mismos residuos tiene el efecto contrario.2 De esta manera, la modificación covalente modifica las interacciones entre proteínas o de estas con el ADN.
Cuando son modificadores diferentes pueden presentar un carácter cooperativo, que es positivo cuando una primera modificación favorece la realización de la siguiente y negativo en el caso contrario. Así, la fosforilación de la beta-catenina en la serina 45 por la caseína kinasa 1alfa permite la fosforilación en las serinas 41, 37 y 33 por la glucógeno sintasa kinasa 3beta, modificación necesaria para su eliminación vía ubiquitina.3 O sea, la modificación determina hasta cuando permanecerá activa la beta-catenina, algo que no forma parte del contenido informativo de esa proteína. Mientras la acetilación de la kinasa dependiente de AMP (AMPK) en las lisinas 31, 33 y 71 por p300 (proteína de 300 kDa), dificulta la fosforilación por la LKB1 (del inglés, Liver Kinase B) de la treonina 172 del lazo de activación de la kinasa manteniendo a AMPK en su forma inactiva.4 Otra vez la modificación covalente determina el grado de actividad de una enzima.
Por último, las modificaciones múltiples pueden crear un código combinatorio con múltiples lecturas dependiendo de los aminoácidos modificados, su localización en la cadena polipeptídica y los grupos modificadores. El más notorio de estos casos es el de los nucleosomas donde las histonas que los forman pueden presentar un amplio grado de modificaciones que determina fundamentalmente el acceso al ADN necesario para procesos tales como la duplicación, transcripción y reparación del ADN. 5 Las modificaciones de las histonas componentes de los nucleosomas se ilustra en la Figura 2.
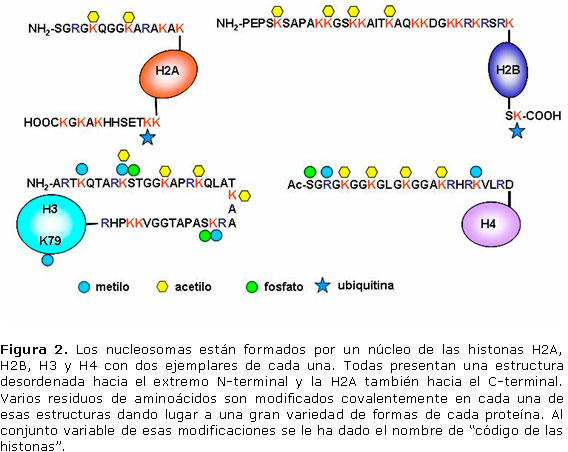
Modificaciones reversibles de proteínas
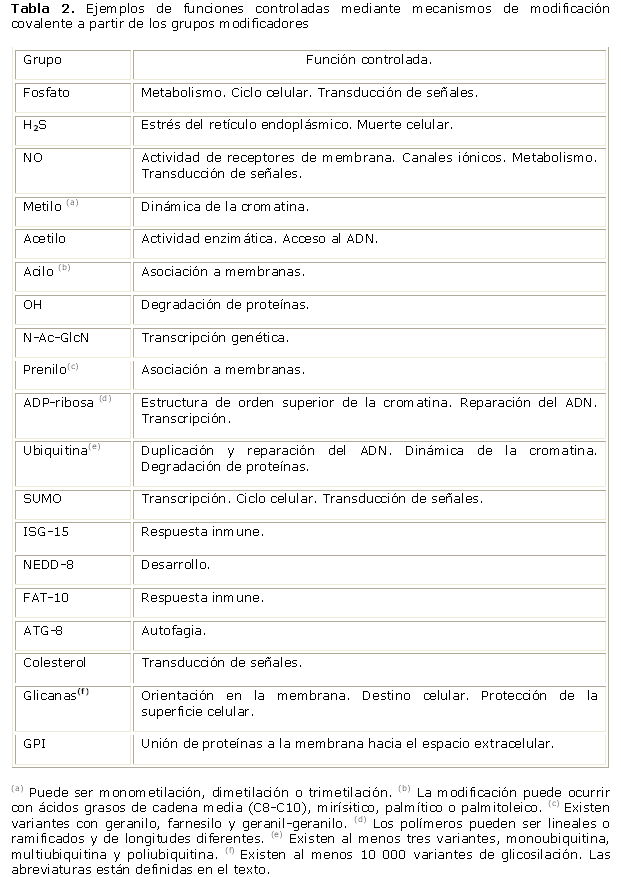
La modificación reversible es uno de los principales mecanismos de regulación y control de numerosas funciones celulares. Los principales grupos modificadores covalentes pueden ser moléculas inorgánicas como fosfato, sulfihidrilo (H2S), óxido nítrico (NO); grupos funcionales orgánicos como metilo, acetilo, acilos, hidroxilo, N-acetil-glucosamina, prenilo, ADP-ribosa; polipéptidos como la ubiquitina y sus similares SUMO (del inglés, Small Ubiquitin-likeMOdifier), ISG15 (del inglés, Interferon-StimulatedGenes 15), NEED8 (del inglés, NEural precursor cell Expressed Developmentally downregulated protein 8), FAT10 (del inglés,HLA-F-Adjacent Transcript 10), ATG-8 (del inglés, AuTofaGia-related 8). En la Tabla 2, se muestran las funciones donde intervienen mecanismos de modificación covalente.
Las células son capaces de regular la cantidad relativa de cada forma de la proteína, y así controla la actividad de las enzimas que participan en el ciclo de modificación. Estos mecanismos proporcionan una respuesta rápida a las necesidades celulares, pues la modificación catalizada por enzimas puede presentar una alta velocidad a la vez que proporciona en muchos casos una amplificación de la señal inicial. También es rápido el final del efecto provocado por la forma predominante pues la modificación en sentido contrario también es catalizada por enzimas. A continuación se exponen los ejemplos más sobresalientes de modificación reversible que permiten demostrar la tesis que se plantea en este trabajo.
Las células son capaces de regular la cantidad relativa de cada forma de la proteína, y así controla la actividad de las enzimas que participan en el ciclo de modificación. Estos mecanismos proporcionan una respuesta rápida a las necesidades celulares, pues la modificación catalizada por enzimas puede presentar una alta velocidad a la vez que proporciona en muchos casos una amplificación de la señal inicial. También es rápido el final del efecto provocado por la forma predominante pues la modificación en sentido contrario también es catalizada por enzimas. A continuación se exponen los ejemplos más sobresalientes de modificación reversible que permiten demostrar la tesis que se plantea en este trabajo.(Tabla 2)
Modificación de la velocidad de reacción de las enzimas
El metabolismo es el área de actividad celular donde primero fueron descritos los mecanismos de modificación covalente, esencialmente el ciclo de fosforilación/desfosforilación. La importancia de este fenómeno fue destacada por el otorgamiento a Edmond H. Fischer y Edwin G. Krebs en 1992 del Premio Nobel de Medicina.
El caso más conocido es el del metabolismo del glucógeno. Dos enzimas controlan este proceso: la glucógeno sintasa (la glucogénesis) y la glucógeno fosforilasa (la glucógenolisis). Mientras la glucógeno sintasa es más activa en su estado no fosforilado, la glucógeno fosforilasa lo es en su estado fosforilado. Esto permite el control coordinado de los dos procesos en respuesta a diferentes estímulos. La hipoglicemia estimula la fosforilación y con ello la glucogenolisis, mientras que la hiperglicemia incrementa la desfosforilación y con ello la glucogénesis.6 De manera que la modificación dota a estas enzimas de una nueva información, es decir, en cuales condiciones actúa una o la otra.
En estas dos enzimas, las modificaciones tienen un efecto acumulativo. La glucógeno sintasa presenta ocho sitios de fosforilación y va perdiendo actividad en la misma medida que esos sitios son modificados. Por su parte la glucógeno fosforilasa está formada por cuatro subunidades (alfa-beta-gamma-delta). La fosforilación de la subunidad alfa aumenta la actividad que se incrementa aún más con la fosforilación de la subunidad beta. Se desprende que la información sobre la intensidad de la actividad depende de la cantidad de grupos modificados.
La proteína kinasa B (PKB) participa en el mecanismo de acción de la insulina. La adición de un grupo NO (óxido nítrico) a un residuo específico de cisteína de la PKB inhibe la actividad catalítica de la enzima en respuesta a la insulina y se asocia con fenómenos relacionados con el envejecimiento. 7 De nuevo la intensidad de la actividad está regulada por una modificación covalente.
Los mecanismos de fosforilación/desfosforilación están muy asociados a los de nitrosilación tanto para seril-(treonil)-proteína kinasa como para tirosil-proteína kinasa.
La gliceraldehído-3-fosfato deshidrogenasa (GAPDH) constituye un buen ejemplo de la acción antagónica de los modificadores. La nitrosilación de la enzima en la cisteína-150 inhibe su función en la glicólisis, pero facilita su transporte al núcleo donde participa en varias funciones incluyendo la transcripción.8 Sin embargo, la sulfihidrilación en el mismo residuo incrementa su actividad ~ 700 veces.9 En una misma proteína modificaciones diferentes pueden determinar la localización y función o cambios en la intensidad de la actividad.
Modificación en la especificidad de sustrato, de acción o de ambas de las enzimas
Un caso aún más interesante es el de la enzima fosfofructokinasa/bisfosfofructo fosfatasa-2 (FFK/FFF-2). Esta enzima en uno de sus extremos presenta un sitio activo de kinasa mientras en el otro es de fosfatasa. En su forma no fosforilada se expresa la actividad de kinasa que cataliza la conversión de la fructosa-6-fosfato en fructosa-2,6-bisfosfato que es un potente activador de la glucólisis. En su estado fosforilado, se expresa la actividad de fosfatasa que cataliza la hidrolisis de la fructosa-2,6-bisfostado a fructosa-6-fosfato lo que propicia la intensificación de la gluconeogénesis. Este es un caso notorio de efectos de la modificación covalente pues se modifica al mismo tiempo tanto la especificidad de acción como la especificidad de sustrato.10 Pero esos cambios se producen en respuesta al estado de la glicemia y, por lo tanto, la modificación también determina cuando se expresa una actividad y cuando la otra.
Modificación de interacciones con otras macromoléculas
La formación de complejos no covalentes de macromoléculas es un fenómeno frecuente y en muchos casos decisivo para la progresión de un proceso. En ocasiones, esas interacciones están moduladas por la modificación covalente de una o varias proteínas que forman el complejo.
Los genes, cuyos productos intervienen en la duplicación del ADN, se encuentran bajo el control del factor de transcripción E2F (del inglés, Early promoter2 Factor). Al inicio de la fase G1, estos genes se encuentran silenciados pues la proteína del retinoblastoma (pRB) está asociada a E2F inhibiendo su función. Cuando, a mediados de G1, se induce la síntesis de la ciclina D, esta forma un complejo con las Cdk4 (del inglés, Cyclin-dependent kinase) o Cdk6 y fosforilan a pRB, que se disocia de E2F y permite la expresión de los genes bajo su control y la progresión de la célula hacia la fase S. Pero la ciclina D es inducida en respuesta a la acción de factores de crecimiento y, por lo tanto la actividad, de las Cdk depende de las condiciones celulares. Se publicó recientemente un estudio detallado del papel de los ciclos de fosforilación y desfosforilación durante el ciclo celular.1
La duplicación del ADN en ocasiones se ve interrumpida por la existencia de daños en esa molécula que provocan la detención del proceso. En esos casos el antígeno nuclear de células proliferantes PCNA (del inglés, Proliferating Cell Nuclear Antigen) es modificado covalentemente mediante la adición de ubiquitina a la lisina 164 (K164) de cada una de sus subunidades. Esta modificación permite la disociación de las polimerasas duplicativas, delta o epsilón, y el reclutamiento de ADN polimerasas de la familia Y que realizan copias menos fidedignas, capaces de continuar la síntesis sobre un molde dañado.11 Esto significa que las interacciones del PCNA cambian como resultado de la modificación. Aunque este modo de continuar es propenso a errores, permite la sobrevivencia de la célula que de otra manera activaría los procesos de muerte celular. Los errores cometidos pueden ser reparados mediante otros de los diversos mecanismos existentes.
Uno de los daños más graves que puede presentar el ADN es la rotura de una o ambas hebras. La proteína más eficiente en el reconocimiento de estos daños es la polimerasa de ADP-ribosa PARP-1 (del inglés, Poly[ADP-Ribosyl]-Polymerase) la cual una vez detectado el daño se automodifica (adición de ADP-ribosa a un residuo de aminoácido) y modifica a las histonas. Esta modificación permite reclutar hacia el sitio de la lesión proteínas que restauran la integridad del ADN.12
En el mecanismo de reparación por escisión de bases, la enzima timidina-ADN glicosilasa separa la base dañada pero se mantiene unida fuertemente al ADN interrumpiendo el proceso de reparación. La unión covalente de SUMO a la enzima disminuye su afinidad por el ADN y permite la separación de la enzima y que el proceso continúe hasta su culminación. 13 Estos dos últimos ejemplos muestran cómo la interacción de unas proteínas con otras puede ser alterada mediante la modificación covalente de una de ellas.
También la transcripción está asociada a mecanismos de modificación covalente. El dominio carboxilo terminal de la subunidad mayor de la ARN polimerasa II presenta una cadena no estructurada con repeticiones Tir1_Ser2_Pro3_Tre4_Ser5_Pro6_Ser7 (YSPTSPS). Los residuos de serina de las posiciones 2, 5 y 7 son fosforilados reversiblemente durante el curso de la transcripción lo que permite la unión de proteínas que participan en el procesamiento del transcrito primario. 14 En efecto, todas las etapas del proceso se realizan con la intervención del mecanismo de modificación covalente postraduccional reversible. Aquí se evidencia la importancia de la localización del residuo aminoacídico modificado, pues es la serina la que se modifica y siempre por fosforilación, pero la fosforilación en cada posición tiene un significado diferente para la etapa de la transcripción.
La siguiente etapa en la expresión de la información genética es la traducción. El proceso se inicia cuando el factor de iniciación eIF4E se une al extremo 5´modificado del ARNm. Sin embargo, en estado de reposo el eIF4E se encuentra unido a la proteína 4BP (del inglés, 4E Binding Protein). La fosforilación de 4BP provoca que esta se separe de eIF4E y este puede unirse al ARNm dando inicio así, a la síntesis de proteínas. 15 La información añadida por la modificación significa cuándo debe comenzar el proceso. Por lo tanto las dos etapas fundamentales del proceso global de expresión de la información genética están vinculadas a mecanismos de modificación covalente postraduccional reversible de proteínas.
Reclutamiento de proteínas
El fenómeno de reclutamiento se observa cuando una proteína se une a un sitio determinado y favorece la unión al mismo de otra proteína que por sí sola no puede hacerlo y que es en realidad la que realizará la función. En ocasiones para el reclutamiento es necesaria la modificación covalente de la proteína reclutadora.
Para la realización de la transcripción es necesario que la maquinaria molecular del proceso tenga acceso a las secuencias específicas del ADN donde debe comenzar el proceso (el promotor). El ADN se encuentra enrollado alrededor de un núcleo de histonas formando el nucleosoma. Cuando los grupos aminos de lisinas pertenecientes a las histonas son acetilados la unión entre las histonas y el ADN se debilita y permite el acceso al promotor. Las lisinas acetiladas sirven como reclutadoras para muchos factores de transcripción que poseen dominios de unión a lisinas acetiladas pero no se unen a las no acetiladas.16 La desacetilación en general tiene el efecto contrario. De nuevo la modificación determina cuándo y dónde.
Sin embargo, hay genes o conjunto de ellos que deben permanecer en estado silente durante la vida de la célula. Para lograr eso, la histona H3 es metilada en la lisina 9 (H3K9) y así recluta la proteína de la heterocromatina 1. La unión de HP-1 a un nucleosoma recluta a una metilasa que modifica al nucleosoma adyacente al cual nuevamente se une HP-1 y el proceso se repite.17 Este mecanismo propaga el estado de heterocromatina hasta arribar a señales que impiden la propagación más allá de ellas. De esta forma grupos de genes son silenciados permanentemente. La modificación covalente determina la inactividad transcripcional de los genes que forman parte de la heterocromatina. La formación de la heterocromatina se ilustra en la Figura 3.

El reclutamiento de proteínas es una fase primordial en el mecanismo de transducción de señales, especialmente en receptores con actividad de tirosil-proteína kinasa. Una vez unido al ligando, el receptor se autofosforila en residuos de tirosina. Estas tirosinas fosforiladas reclutan proteínas que poseen un dominio SH2 y que a su vez resultan fosforiladas por el receptor.18 Aunque la tirosina fosforilada es el elemento clave para el reclutamiento, los aminoácidos que están a su alrededor determinan cuál proteína con dominio SH2 se unirá al receptor. Este mecanismo es general para otras tirosil-proteínas kinasas que no son receptoras. De nuevo la modificación covalente influye en las interacciones entre proteínas.
Modificaciones irreversibles de proteínas
En ocasiones, el grupo modificador no es separado de la proteína pues la modificación cumple una función permanente. Las modificaciones irreversibles pueden influir en la velocidad de recambio de las proteínas, su localización subcelular o su actividad biológica específica.
Efecto sobre la actividad específica de proteínas
Los casos más notorios se refieren a proteínas o polipéptidos que actúan como mensajeros intercelulares. Al menos tres mensajeros moleculares intercelulares experimentan modificaciones covalentes imprescindibles para su actividad. La grelina una hormona digestiva que estimula el apetito requiere la adición de residuos de ácidos grasos de cadena media (C8-C19) en la serina 3 para su total actividad.19 Por su parte, la familia de ligandos WNT (del inglés wingless, sin alas) son acilados para mostrar su acción. Por último, los ligandos de la familia erizo (en inglés, hegdehog) son modificados por la adición de ácido palmítico en la cisteína-77, modificación necesaria para su secreción, y un residuo de ácido palmitoleico es esterificado a la serina-209, modificación necesaria para su actividad.20 En todos los casos anteriores, la modificación covalente influye sobre la actividad de las proteínas.
Determinación de la localización subcelular
Aunque las proteínas suelen tener una señal interna que indica su localización celular; en muchos casos, estas señales aparecen en estado críptico y solo se exponen a los mecanismos de transporte después de una modificación covalente.
Así algunas proteínas que realizan sus funciones asociadas a las membranas requieren el procesamiento del extremo carboxilo terminal. Mediante un mecanismo en tres etapas, un grupo farnesilo se añade al grupo sulfihidrilo de una cisteína. Después se eliminan los tres aminoácidos posteriores a la cisteína prenilada y finalmente el carboxilo del último aminoácido es metilado. De esta forma, la proteína es dotada de una cadena apolar (el grupo prenilo) que le permite asociarse con la membrana. Así sucede, por ejemplo, con la proteína Ras 21 que es un importante intermediario en la vía de transducción de señales de hormonas y factores de crecimiento.
En otros casos, se modifica el extremo N terminal mediante la adición de grupos acilos largos, como el miristilo o el palmitilo. El grupo acilo añadido permite la asociación de la proteína a la membrana como sucede con la subunidad alfa de las proteínas G triméricas que actúan acopladas a receptores de membrana.22
Las proteínas lisosomales tienen que ser procesadas para añadirle la señal de localización que consiste en un oligosacárido que contiene manosa-6-fosfato. La manosa-6-fosfato es reconocida por un receptor similar al del factor de crecimiento insulinoide 2 (IGF-2) y de esta forma se incorpora a los lisosomas nacientes en el aparato de Golgi.23
Otra modificación que determina la localización de proteínas es la adición de glucosil-fosfatidil-inositol al extremo C-terminal de proteínas. Esta modificación une a la capa externa de la membrana plasmática proteínas con importantes funciones en la transmisión de señales y la respuesta inmune.24 En todos los casos anteriores la localización definitiva y, por tanto, funcional de la proteína depende de la modificación covalente de alguno de sus grupos.
Modificación de la vida media de las proteínas
Existen proteínas que solamente pueden estar activas durante un período corto de tiempo (el llamado efecto de intervalo). Una vez concluido ese tiempo la forma más radical de eliminar su función es la degradación de la proteína. Para la realización de la proteólisis, las proteínas deben ser marcadas previamente mediante un mecanismo de modificación covalente. El sistema mejor conocido es la vía ubiquitina-proteasoma. Para más detalles sobre este sistema ver el artículo de reciente publicación "La vía ubiquitina-proteasoma. ¿Destruir o construir?" 25
Sin embargo, en ocasiones la adición de ubiquitina requiere de una modificación previa. Así, la proteína reclutadora de la glucógeno sintasa R5/PTG (del inglés, Protein Target of Glycogen) es fosforilada por la AMPK que la hace sustrato del complejo laforina-malina que la marcan con ubiquitina y entonces es degradada por el proteasoma.26 En estos casos la modificación covalente determina hasta cuándo una proteína debe actuar.
Otra modificación que determina la proteólisis es la hidroxilación. El caso más conocido es el del factor inducible por hipoxia HIF (del inglés, Hypoxia-Inducible Factor). Este factor está formado por dos subunidades, una de localización nuclear, HIF-beta y otra citoplasmática HIF-alfa. En condiciones de aporte normal de oxígeno HIF-alfa es hidroxilado en dos residuos de prolina. Esta modificación permite su reconocimiento por el complejo BC-VHL (formado por las elonguinas B y C y de la proteína de von Hippel Lindau) que lo marca con ubiquitina y es degradado por el proteasoma. Cuando los niveles de oxígeno disminuyen, la hidroxilación no es posible y HIF-alfa es transportado al núcleo donde en unión de HIF-beta induce la expresión de un gran número de genes relacionados con la respuesta a la hipoxia. 27 En este caso la modificación covalente determina dónde debe localizarse la proteína de acuerdo con las condiciones celulares.
CONCLUSIONES
Como se ha explicado el mecanismo de modificación covalente de proteínas es un evento crucial en múltiples funciones celulares, tales como el metabolismo, la localización de proteínas, la expresión de la información genética, la transducción de señales, la duplicación y reparación del ADN, entre otras.
La adición de grupos modificadores en residuos específicos introduce en la proteína una información nueva que modifica sus propiedades generalmente de forma transitoria y que permite la adaptación del organismo ya sea monocelular o pluricelular a los cambios ambientales. Por eso la modificación covalente de proteínas puede ser considerada como un nivel nuevo y cualitativamente superior de la información molecular.
REFERENCIAS BIBLIOGRÁFICAS
1. Hernández Fernández, RA. Kinasas y fosfatasas: el yin y el yan de la vida. Rev Haban Cien Med. 2012:11(1)15-24.
2. Dentin R, Hedrick S, Xie J, Yates J, Montminy M. Hepatic glucose sensing via the CREB coactivator CRTC 2. Science. 2008; 319: 1402-05.
3. Valenta T, Hausmann G, Basler K. The many faces and functions of â-catenin. EMBO J. 2012; 31: 2714-36.
4. Lin YY, Kiihl S, Suhail Y, Liu SY, Chou YH, Kuang Z, et al. Functional dissection of lysine deacetylases reveals that HDAC1 and p300 regulate AMPK. Nature. 2012; 482: 251-5.
5. Suganuma T, Workman JL. Signals and combinatorial functions of histone modifications. Annu Rev Biochem. 2011; 80:473-499.
6. Roach PJ, Depaoli-Roach AA, Hurley TD, Tagliabracci VS. Glycogen and its metabolism: some new developments and old themes. Biochem J. 2012; 441: 763-87.
7. Wu M, Katta A, Gadde MK, Liu H, Kakarla SK, Fannin J, et al. Aging associated dysfunction of Akt/protein kinase B: S-nitrosylation and acetaminophen intervention. PLoS ONE. 2009; 4: e6430-e36.
8. Sen N, Hara MR, Kornberg MD, Cascio MB, Bae BI, Shahani N, et al. Nitric oxide-induced nuclear GAPDH activates p300/CBP and mediates apoptosis. Nature Cell Biol. 2008; 10: 866-73.
9. Mustafa AK, Gadalla MM, Sen N, Kim S, Mu W, Gazi SK, et al. H2S signals through protein Ssulfhydration. Sci Signal. 2009; 2(96): ra72.
10. Rider MH, Bertrand L, Vertommen D, Michels PA, Rousseau GG, Hue L. 6-Phosphofructo-2-kinase/fructose-2,6-bisphosphatase:head-to-head with a bifunctional enzyme that controls glycolysis. Biochem J. 2004; 381: 561-79.
11. Zhang W, Qin Z, Zhang X, Xiao W. Roles of sequential ubiquitination of PCNA in DNA- damage tolerance. FEBS Letters. 2011; 585: 2786-94.
12. Messner S, Hottiger MO. Histone ADP-ribosylation in DNA repair, replication and transcription. Trends Cell Biol. 2011; 21(9): 534-42.
13. Wilkinson KA, Henley JM. Mechanisms, regulation and consequences of protein SUMOylation. Biochem J. 2010; 428: 133-45.
14. Hsin JP, Manley JL. The RNA polymerase II CTD coordinates transcription and RNA processing. Genes Dev. 2012; 26: 2119-37.
15. Aitken CE, Lorsch JR. A mechanistic overview of translation initiation in eukaryotes. Nature Struc Mol Biol. 2012; 19: 568-76.
16. Hodawadekar SC, Marmorstein R. Chemistry of acetyl transfer by histone modifying enzymes: structure, mechanism and implications for effector design. Oncogene. 2007; 26: 5528-40.
17. Beisel C, Paro R. Silencing chromatin: comparing modes and mechanisms. Nature Rev Genetics. 2011; 12: 123-35.
18. Koch S, Tugues S, Li X, Gualandi LY, Claesson-Welsh L. Signal transduction by vascular endothelial growth factor receptors. Biochem J. 2011; 437: 169-83.
19. Dong CX, Brubaker PL. Ghrelin, the proglucagon-derived peptides and peptide YY in nutrient homeostasis. Nature Rev Gastroenterol Hepatol. 2012; 9:705-15.
20. Ryan KE, Chiang C. Hedgehog Secretion and Signal Transduction in Vertebrates. J Biol Chem. 2012; 287: 17905-13.
21. Ahearn IM, Haigis K, Bar-Sagi D, Philips MR. Regulating the regulator: post-translational modification of RAS. Nature Rev Mol Cell Biol. 2012; 13: 39-51.
22. Linder ME Deschenes RJ. Palmitoylation: policing protein stability and traffic. Nature Rev Mol Cell Biol. 2007; 8: 74-84.
23. Luzio JP, Pryor PR, Bright NA. Lysosomes: fusion and function. Nature Rev Mol Cell Biol. 2007; 8: 622-32.
24. Paulick MG, Bertozzi CR. The Glycosylphosphatidylinositol Anchor: A Complex Membrane-Anchoring Structure for Proteins. Biochemistry. 2008; 47: 6991-7000.
25. Hernández Fernández RA. La vía ubiquitina-proteasoma. ¿Destruir o construir? Ese es el dilema. Rev Haban Cien Med. 2013: 12(1):.
26. Vernia S, Solaz-Fuster MC, Gimeno-Alcañiz JV, Rubio T, García-Haro L, Foretz M, Rodríguez de Cordoba S, Sanz P. AMP-activated protein kinase phosphorylates R5/PTG, the glycogen targeting subunit of the R5/PTG-protein phosphatase 1 holoenzyme, and accelerates its down-regulation by the laforin-malin complex. J Biol Chem. 2009; 284: 8247-55.
27. Hernández Fernández RA. Fundamentos moleculares de la enfermedad de von HippelLindau. Rev Cub Invest Bioméd. 2010;29(2):262-273. ISSN 1561-3011.
Recibido: 1ro de Septiembre de 2013
Aprobado: 15 de Noviembre de 2013

{kind=link}
{kind=link}