










 Serviços customizados
Serviços customizados Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCTION
Eggplant (Solanum melongena L.) is a species of the Solanaceae family and constitutes an important crop in several tropical and subtropical regions of the world (Kashyap et al., 2003). In addition to food use, eggplant is used in folk medicine for the treatment of diseases associated with high cholesterol levels (Aiso et al., 2014; Meyer et al., 2014). Among their proven activities are reported the cardio-protective action (Das et al., 2011), antioxidant and antidiabetic (Kwon et al., 2008; Kaur et al., 2014).
The application of in vitro culture methodologies has resulted in the availability of several techniques that complement conventional eggplant breeding, such as embryo rescue, in vitro selection, somatic hybridization and genetic transformation (Kashyap et al., 2003). Clones can be produced by distinct morphogenic pathways from different types of explants, genotypes and growth regulators (Magioli and Mansur, 2005). However, in the production of clones, genetic stability can be broken due to somaclonal variations induced during in vitro culture (Larkin and Scowcroft, 1981; Bednarek et al., 2007).
However, the genotype is considered one of the main factors predisposing to the occurrence of genetic variations (Bairu et al., 2011). Regarding culture, endogenous hormones should be considered, since it can influence the sensitivity of plant cells to the plant growth regulators added to the culture media, so that in addition to the type, the concentration also plays an important role in the generation of variants (Kaur et al., 2011a; Krishna et al., 2016). Another important factor to consider is the morphogenetic pathway induced for the production of the clones, as it is known that in indirect organogenesis or embryogenesis, callus development reduces the genetic stability of the clones, due to higher mutation rate as a consequence of a phase of differentiation followed by uncontrolled cell divisions (Krishna et al., 2016). Therefore, is a need to assess the genetic stability of these plant materials, regardless of the purpose of the work (Kaur et al., 2011a; Kaur et al., 2011b).
Although many genetic stability monitoring strategies are available, the use of molecular markers has been consolidated based on the practicality and comprehensiveness of genome analysis. Several types of molecular markers have been used for the evaluation of genetic stability (Bairu et al., 2011). Some are based on restriction and hybridization of DNA sequences, others are based on Polymerase Chain Reaction (Grover and Sharma, 2016), as Randomly Amplified Polymorphic DNA (RAPD). RAPD was developed two decades ago for identifying variants. Currently is the technique most used both in breeding programs (Mallaya and Ravishankar, 2013) and in germplasm conservation programs (Naseer and Mahmood, 2014).
Its use is mainly based on simplicity, speed and relatively low cost when compared to other molecular markers (Grove and Sharma, 2016). The reaction requires only a primer (decamer oligonucleotide) of arbitrary sequence and the polymorphisms result from the sequence variation at the annealing sites and / or variations in the target sequence length between the primer binding sites (Grover and Sharma, 2016). Amplification products are visualized as bands on the electrophoresis gel. Therefore, any change in the sequence is translated by the presence or absence of the bands when the samples are compared with each other or with the matrix. RAPD allows detecting single-base variations in point mutations of any DNA region (Sirvent and De Castro, 2012). Although it is a dominant marker, the requirement of minimal amounts of DNA represents a major advantage over other random markers in the analysis of cloned in vitro plants (Bairu et al., 2011).
In Solanum melongena, the development of the molecular marker RAPD was reported to determine the genetic stability of clones of some cultivars (cv. Nirrala and cv. Arka Shirish) in vitro produced in response to cytokinins alone or combined with auxins (Mallaya and Ravishankar, 2013; Naseer and Mahmood, 2014).
Solanum melongena cv. Florida Market made possible the production of the hybrid F-100 in Brazil. It replaced the old plantations due to its high productivity (Sousa et al., 1997). For this cultivar, although protocols for induction of clones by different morphogenetic pathways have been defined (Magioli and Mansur, 2005), the genetic stability of the clones has not been evaluated. Therefore, this paper aimed to evaluate the genetic stability of Solanum melongena cv. Florida market clones obtained in vitro in response to different types of growth regulators by primers capable of revealing polymorphisms.
MATERIALS AND METHODS
Production of in vitro clones
Plant material and culture conditions
Seeds of Solanum melongena (L.), cultivar ‘Florida Market’, were obtained commercially (Isla S.A.®). For establishing of in vitro tissue culture, seeds were soaked in a commercial detergent enriched tap water for 5 min, surface disinfected with 2% (w/v) sodium hypochlorite solution (Tricloro-S-Triazina Triona) and Tween 20 (0.001%) for 20 min and then, rinsed three times in sterile distilled water. Following these seeds were aseptically germinated on Murashige and Skoog (MS) medium (Murashige and Skoog, 1962) containing half-strength MS elements, 1.5% sucrose (w/v) (Sigma Chemical, St. Louis). This medium was solidified with 0.7% (w/v) agar (Merck®), for induction of the germination. Seeds were cultured and, after 21 days, the seedlings were used as explants source for tissue culture (Magioli et al., 1998). All the cultures were grown in a culture room at 25 ± 2 °C, 16 h photoperiod with light at 46 µM m-2 s-1 (Phillips "Cool white" fluorescent tubes).The seedlings visually more vigorous were selected for the regeneration in presence of growth regulators.
In vitro culture
Leaves segments (5.0 cm2) of the in vitro germinated seeds were inoculated in MS medium supplemented with 3% sucrose (w/v) and different growth regulators in defined concentrations of benzilaminopurine (11 µM BAP) (Sharma and Rajam, 1995; Muthusamy et al., 2014) or thidiazuron (0.2 µM TDZ) (Magioli et al., 1998), aiming to obtain shoots by organogenesis (Magioli and Mansur, 2005). Besides, it were inoculated I similar culture medium with naphthaleneacetic acid (54 µM NAA) (Tarré et al., 2004), for obtaining plants by somatic embryogenesis (Magioli et al., 2001). The somatic embryos at cotyledonary stage were transferred to MS0 solidified with 1% Phytagel in order to regenerate plants. Each growth regulator was tested on clones obtained from the same seed. The pH of the medium was adjusted to 5.8 prior to autoclaving at 120 °C for 15min.
The calluses developed from explants cultured in the presence of these growth regulators (BAP or TDZ) were fragmented (about 0.5 cm2), transferred to a basal medium MS free of growth regulators (MS0) and cultured for 15 days. Developed shoots (1.5-2.0 cm in length) were excised and transferred to MS medium containing 3% (w / v) sucrose and 0.6 mM indoleacetic acid (IAA) for rooting.
Aiming to increase the amount of plant material for DNA extraction, all the complete plants obtained in response to different plant growth regulators, independently of the regeneration pathway, were cloned from the multiplication of nodal segments (0.5 cm length) cultured in MS0 medium (Magioli et al., 1998).
Evaluation of genetic stability
RAPD technique
DNA isolation: the donor plant that provided the largest number of clones (about 120) were selected for genetic stability evaluation. Total genomic DNA was isolated from 250 mg of leaves of 30-45 randomly chosen clones of each tested plant growth regulators, and also from the donor plant, using the standard CTAB method. The concentration and the quality of genomic DNA was determined by electrophoresis in 0.8% (w/v) agarose gel containing ethidium bromide (0.5 μg ml-1) in 1× TBE buffer.
DNA amplification: a total of 10 decamers oligonucleotide primer synthesized at Invitrogen Custon Primers and available in laboratory stock (Table 1) were tested for RAPD analysis. The primers used had GC content ranged from 60 to 80%. Nine primers were selected using DNA from germinated seeds as matrices, resulting in simple and reproducible bands for the RAPD analysis of the clones produced by different culture conditions. RAPD reaction was performed in 25 µl reaction mixture containing Taq DNA polymerase (Biotools®) (2.0 U), AmpliTaq buffer (1x) with 4.0 mM MgCl2, random decamer primer (1.0 pmol/reaction) 0.25 mM, deoxynucleotide triphosphate (dNTPs) (Biotools®) and 5 ng of genomic DNA. Amplification was performed in a PTC-100 MJ Thermal Cycler (MJ Research, Inc.) programmed for 45 cycles as follows: preliminary denaturation of DNA at 94 °C for 4 min, followed by 45 cycles at 94 °C for 1 min, 35 °C for 2 min and 72 °C for 2 min, which is followed by a final extension cycle of 5 min at 72 °C.
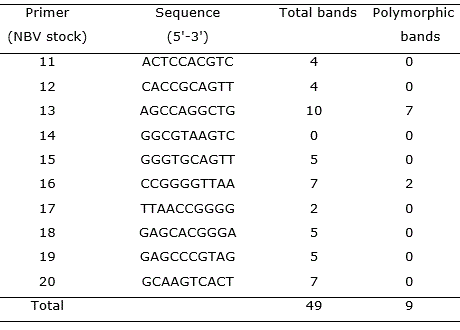
Table 1 RAPD primers used to analysis of Solanum melongena cv. Florida Market and products amplified from clones obtained by different regenerative process.
The amplification products of the plants DNA originated to the same explant were resolved by electrophoresis (3V/cm) on a 1.4% agarose gel prepared in TBE buffer containing ethidium bromide (0.5 mg l-1 ). The images were photographed with digital camera under UV light using red filter. The size of the amplification products was estimated from a 100 bp DNA ladder (Sigma). All the reactions were repeated at least twice and only those bands reproducible on all runs were considered for analysis.
RESULTS
Production of in vitro clones
The seeds used presented a high germination rate after 21 days, in the order of 90-100%. After 30 days of culture under the described conditions, the shoots were produced from leaves explants in response to BAP (Figure 1 A) and TDZ (Figure 1 B) by indirect organogenesis.
The subculture of organogenic calli in the presence of growth regulators (BAP or TDZ) and then transferred to a basal culture medium MS free of growth regulators (MS0) made it possible to obtain a large number of shoots/explants (18-20). Developed shoots produced roots about 2 weeks after.
Besides, after 30 days of culture, the explants cultivated in the presence of NAA developed embryogenic calli at high frequencies (95%) (Figure 1 C). The somatic embryos converted to whole plants, phenotypically normal, after 20-25 days.

Figure 1 In vitro regeneration of Solanum melongena cv. Florida Market plants from leaf explants. A. Indirect organogenesis in MS medium supplemented with 11 µM BAP, B. Indirect organogenesis in MS medium supplemented with 0.2 µM TDZ, C. Somatic embryogenesis on MS medium supplemented with 54 µM NAA. Bar = 1cm.
Evaluation of genetic stability
It was obtained between 75-100 ng the DNA of donor plant and from the clones of each plant growth regulators treatment.
Among the 10 primes used, nine revealed identifiable bands and produced 49 scorable bands with an average of 5.44 bands per primer. Out of the nine random primers used, only two, yielded nine polymorphic bands (Table 1).
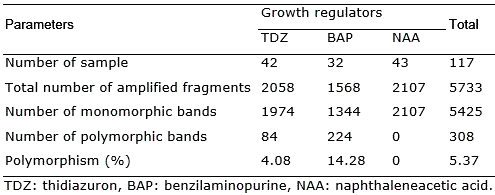
Considering all samples, a total of 308 polymorphic bands were produced among the 5733 amplified bands in total, which corresponds to a polymorphism rate of 5.37% (Table 2).
Table 2 RAPD analysis of Solanum melongena cv. Florida Market clones obtained in response to different plant growth regulators.
From TDZ-induced clones, the primer 13 originated polymorphic bands among fourteen clones evaluated (Figure 2 A). With this primer, the sample T24 showed a band of 750 pb. It was absent in the other samples, and it was also observed the absence of a band in sample T36 (550 pb) (Figure 2 A). No polymorphisms were observed with the primer 16 (Figure 2 C).
In the case of BAP-induced clones, the primer 13 allowed the identification of four variants among 14 clones obtained from explants. In B6 sample, five bands (1500, 1300, 800, 700, 550 pb) present in the control were absent. The clones B12, B13 and B14 also showed a similar pattern, with the absence of a band found in the control (550 pb) (Figure 2 B).
The clones obtained from explants in the presence of BAP also showed polymorphisms with the primer 16. Two bands were absent in the sample B6 (1500 and 800 pb), one band in the sample B12 (800 pb) and B13 (800 pb) (Figure 2 D).

Figure 2 Amplification pattern by RAPD in Solanum melongena cv. Florida Market plants from indirect organogenesis. The amplification products of the plant DNA originated to the same explant were resolved by electrophoresis (3 V/cm) on a 1.4% agarose gel, prepared in TBE buffer containing ethidium bromide (0.5 mg m l-1). A. Profile obtained with primer 13 in TDZ - induced clones; B. profile obtained with primer 13 in BAP - induced clones; C. Profile obtained with primer 16 in TDZ-induced clones; D. Profile obtained with primer 16 in BAP-induced clones. M: molecular marker (100 bp). C: control. Arrows indicate polymorphic bands.
Results of cloned samples from NAA-induced somatic embryogenesis showed no polymorphisms among clones obtained, regardless of the primer tested (data not show).
DISCUSSION
Although the genetic stability of some in vitro produced eggplant cultivars has been studied (Mallaya and Ravishankar, 2013; Naseer and Mahmood, 2014), this is the first report in the S. melongena cv. Florida Market, an important cultivar for the eggplant breeding (Sousa et al., 1997). Plants of this cultivar can be in vitro produced for hybridization, however, it should be considered that many factors may influence the rate of genetic variation in plant tissue culture. The genotype and the use of plant growth regulators have been considered the most important factors, affecting the genetic stability according to the type, concentrations, combinations and duration of the culture (Bairu et al., 2011; Kaur et al., 2011a; Sales and Butardo, 2014; Krishna et al., 2016). In this work, the genetic stability of clones induced by different growth regulators (BAP, TDZ and NAA) was evaluated in predefined concentrations as a function of the number of complete plants obtained.
The influence of plant regulators on the genetic stability of in vitro cloned eggplant has been studied in different genotypes or cultivars cultured in the presence of cytokinins alone or combined with auxins. Some papers evaluating the genetic stability of these in vitro plants by RAPD are available in the scientific literature. For instance, a study with four different genotypes of eggplant (Meizi, Xianfeng I, Heijuren and Jiuye) showed absence of polymorphism in organogenic shoots produced directly in response to zeatin and AIA (Xing et al., 2010). The absence of polymorphism has also been reported in clones of S. melongena cv. Arka Shirish produced in response to TDZ (Mallaya and Ravishankar, 2013). On the other hand, the evaluation of clones of cv. Nirrala, the variety of S. melongena, produced by indirect organogenesis in response to combinations of BAP, NAA and 2,4-Dichlorophenoxyacetic acid (2,4-D) showed a high polymorphism rate (26.99%) (Naseer and Mahmood, 2014).
In this work, only S. melongena cv. Florida Market was used with the clones being produced in response to each plant growth regulator, originating from the same seed. Among the clones produced by indirect organogenesis in response to BAP or TDZ, at standard concentrations for the species, 308 polymorphic bands were detected. The polymorphism rate (5.37%) was higher than the rate of spontaneous mutations (1/107), indicating a potential effect of variation induced by the cytokinins used in the culture. The concentrations used were previously standardized according to the number of shoots, but sub or supra-optimal levels to ensure clonal stability are likely.
In these sense, the evaluation in other species of the genus Solanum showed absence of polymorphism in clones produced in response to BAP and/or TDZ from S. surattense (Yadav et al., 2010) and S. aculeatissimum (Ghimire et al., 2012). On the other hand, genetic variation was high (20-53%) in callus cultures of S. tuberosum by the combination of zeatin 1.5 mg l-1 and BAP 1.0 mg l-1 (Munir et al., 2011).
Considering that polymorphism may be the expression of genetic variations of errors in the in vitro replication process that would alter DNA nucleotide sequences, plant growth regulators are considered mutagenic agents because of their direct action on cell division, with potential to induce disorganized growth and greater genetic instability (Bairu et al., 2011; Krishna et al., 2016). In addition, cytokinins increase indirectly, inducing multiplication of adventitious shoots and increased frequency of polyploid cells, whereas of auxins cause increased DNA methylation rate and chromosomal instability (Bairu et al., 2011). The polymorphism rate obtained in this research could have been influenced by the regeneration pathway, since the polymorphism was observed only in the clones obtained from organogenesis, whereas the clones from somatic embryogenesis showed a monomorphic pattern. There are no reports of genetic instability in clones derived from somatic embryogenesis in eggplant. However, there are several reports of somaclonal variation detected by RAPD from somatic embryogenesis in different species (Dey et al., 2015, Hazubska-Przybyt and Dering, 2017).
The two primers capable of detecting polymorphisms among the clones obtained in this research represent appropriate markers for the evaluation of genetic stability and identification of somaclonal variation, with possible application in the research and genetic improvement of eggplant cultures.
CONCLUSIONS
The response to auxins and cytokinins influences the appearance of genetic variation in eggplant clones produced and maintained in vitro. Indirect organogenesis induced for BAP or TDZ is genetically most unstable pathway than embryogenesis, with two primers identified for monitoring the genetic stability of eggplant.

