










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCuban Journal of Agricultural Science
versión On-line ISSN 2079-3480
Cuban J. Agric. Sci. vol.50 no.3 Mayabeque jul.-set. 2016
Cuban Journal of Agricultural Science, 50(3): 393-402, 2016, ISSN: 2079-3480
ORIGINAL ARTICLE
Effect of five inclusion levels of mulberry (Morus alba cv. cubana) on methanogens and some main cellulolytic populations within rumen liquor of water buffalos (Bubalus bubalis)
Efecto de cinco niveles de inclusión de morera (Morus alba vc cubana) en los metanógenos y algunas de las poblaciones celulolíticas principales presentes en el líquido ruminal de búfalos de río (Bubalus bubalis)
Niurca González,I Adibe L. Abdalla,II Juana Galindo,I María Regina Santos,II Patricia Louvandini,II Helder Louvandini,II
IInstituto de Ciencia Animal. Apartado Postal 24, San José de las Lajas, Mayabeque, Cuba.
IICentro de Energía Nuclear para la Agricultura. Laboratorio de Nutrición Animal. Universidad de São Paulo. Brasil.
ABSTRACT
In order to study the effect of five inclusion levels of mulberry (Morus alba cv. cubana) on methanogens and some main cellulolytic populations within rumen liquor of water buffalos (Bubalus bubalis), and contribute to the knowledge of the action mechanism by which the Cuban variety of mulberry reduces ruminal methanogenesis, an in vitro fermentation was carried out and five inclusion levels (0, 15, 20, 25 and 30%) of M. alba vc. cubana to a diet based on star grass (Cynodon nlemfuensis) were evaluated. Total, methanogenic and cellulolytic (Fibrobacter succinogenes, Ruminococcus albus) bacteria, and fungi were identified and quantified by polymerase chain reaction in real time. The percentage of cellulolytic and methanogenic bacteria, regarding total bacteria population was evaluated, as well as the delta delta Ct (ΔΔCt) and the expression of different ruminal microbial populations, in relation to control treatment. The inclusion of M. alba cv. cubana did not affect the studied ruminal microbial populations. The percentage of representation of F. succinogenes, R. albus and methanogens, regarding total bacteria was not affected with the inclusion of this variety of mulberry on the diet. Values of ΔΔCt and expression relative to control treatment of microbial populations evidenced that diets containing mulberry tree had the same difference and the same expression relative to the control treatment. It can be concluded that inclusion of M. alba cv. cubana on diet not affect the main microbial populations that degrade the fiber in the rumen, which could be used as a strategy for reducing methane production in the rumen. The mechanism by which this plant is able to reduce ruminal methanogenesis is not through direct effect on methanogens.
RESUMEN
Para estudiar el efecto de la inclusión de diferentes niveles de morera (Morus alba vc cubana) en los metanógenos y algunas de las principales poblaciones de microorganismos celulolíticos del rumen de búfalos de río (Bubalus bubalis), y contribuir al conocimiento del mecanismo de acción mediante el cual la variedad cubana de morera reduce la metanogénesis ruminal, se realizó una fermentación in vitro y se evaluaron cinco niveles de inclusión (0, 15, 20, 25 y 30 %) de M. alba vc cubana a una dieta base de pasto estrella (Cynodon nlemfuensis). Se identificaron y cuantificaron bacterias totales, metanogénicas, celulolíticas (Fibrobacter succinogenes, Ruminococcus albus) y hongos mediante la reacción en cadena de la polimerasa en tiempo real. Se determinó el porciento de bacterias celulolíticas y metanogénicas relativo a la población de bacterias totales, el delta delta Ct (∆∆Ct) y la expresión de las diferentes poblaciones microbianas ruminales relativo al tratamiento control. La inclusión de M. alba vc cubana no afectó las poblaciones microbianas ruminales estudiadas. El porcentaje de representación de F. succinogenes, R. albus y metanógenos con respecto a las bacterias totales tampoco se afectó con la inclusión de esta variedad de morera en la dieta. Los valores de ∆∆Ct y la expresión relativa al tratamiento control de las poblaciones microbianas evidenciaron que las dietas que contenían morera tuvieron la misma diferencia y la misma expresión relativa al tratamiento control. Se concluye que la inclusión de M. alba vc cubana en la dieta no afecta las principales poblaciones microbianas que degradan la fibra en el rumen, lo que podría utilizarse como estrategia para reducir la producción de metano en el rumen. El mecanismo por el que esta planta logra reducir la metanogénesis ruminal no es por efecto directo sobre los metanógenos.
INTRODUCTION
In most of tropical countries, ruminant production is limited due to the low quality of feed provided to them, which are nitrogen deficient and low digestibility. There are different researches that search for alternatives to improve feed quality. Mulberry is used for ruminant nutrition and its leaves are provided as a main source of feed for sheep and goats (Bakshi and Wadhwa 2007). This plant has also been used for substituting concentrates in diets cows, sheep, goats and buffalos (Anbarasu et al. 2004, Kandylis et al. 2009, Foiklang et al. 2011).
Mulberry grows under varies climatic conditions (Foiklang et al. 2011) and have a great capacity of biomass production. Its leaves have a great content of protein (from 15.0 % to 27.6 % of DM) and high in vivo digestibility of DM (from 75 % to 85 %) (Ba et al. 2005). Therefore, it has a high potential as protein-rich forage for animal production (Tan et al. 2012). This plant has also been evaluated as part of nutritional strategies, based on use of tree and shrub species to reduce methane production by ruminants.
Delgado et al. (2007) and González et al. (2010) studied the effect of inclusion of different levels of mulberry on control of ruminal methanogenesis. These authors found that 25 and 30 % of inclusion reduced the formation of this in vitro gas, which is very beneficial. It is also known the negative impact of methane emission to the environment, and losses from 2 to 12 % of energy from feed consumed by the animal, as a consequence of the formation of this gas within the rumen. In addition to this potential to reduce methane formation in the rumen, there are properties and potentialities of this plant for its use in ruminant feed.
Decrease of methane formation in the rumen can be obtained by many action mechanisms. Direct inhibition of methanogens is one of them (McAllister and Newbold 2008). However, it is not known the mechanism by which mulberry decreased ruminal methanogenesis, in studies conducted by Delgado et al. (2007) and González et al. (2010). In addition, it is also necessary to know the effect of this plant on some of the main ruminal populations that are involved in fibrous material degradation, because strategies for controlling ruminal methanogenesis are effective, only if they are able to reduce the formation of this gas without affecting microbial populations that degrade fiber (Soliva et al. 2003).
The objective of this study was to analyze the effect of different inclusion levels of mulberry (M. alba cv cubana) on methanogenic microorganisms and some of the main cellulolytic populations (Fibrobacter succinogenes, Ruminococcus flavefaciens and fungi) from the rumen of water buffalos (Bubalus bubalis), and contribute to the knowledge of the action mechanism by which mulberry tree reduces ruminal methanogenesis.
MATERIALS AND METHODS
The research was conducted in cooperation between the Institute of Animal Science (ICA, initials in Spanish), from Cuba, and the University of São Paulo (CENA/USP) from Brazil. In vitro fermentation took place at ICA, according to the technique described by Theodorou et al. (1994). Molecular techniques were implemented at CENA / USP.
Animals and diet. Two adult male water buffalos (Murrah breed), with a single cannula in rumen and average weight of 453 kg were used as donors of ruminal liquor. Animals were housed in individual cubicles, with shade and free access to water and feed. All received star grass (Cynodon nlemfuensis) forage.
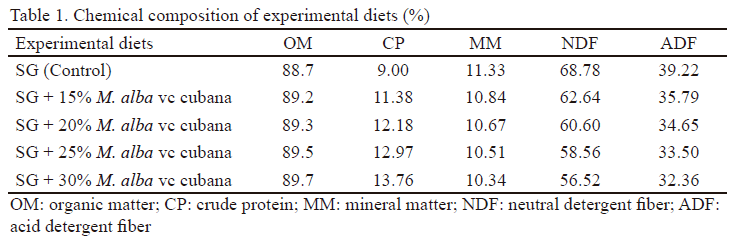
Experimental diets. Five diets were evaluated: 1) 100% star grass (SG) (control), 2) SG + 15% M. alba cv. cubana, 3) SG + 20% M. alba cv. cubana, 4) SG + 25% M. alba cv. cubana and 5) SG + 30 % M. alba cv. cubana.
Mulberry plants came from the Estación Experimental de Pastos y Forrajes “Indio Hatuey”, Matanzas province, Cuba. These plants grew in a red ferrallitic soil and were fertilized with chicken manure. Their leaves with petioles and young stems were manually cut, simulating animal selection. The SG was cut by hand in grazing areas from the Institute of Animal Science, Cuba. The plant material was dried in an oven at 60 °C and ground in hammer mill up to reach a particle size of 1mm.
The experimental diets underwent the analysis of chemical composition according to AOAC (2016) and fibrous fractions were analyzed according to Goering and van Soest (1970). Table 1 shows the chemical composition of the experimental diets.
Experimental Procedure. An amount of 0.5 g from each of the treatments were weighed and added to the corresponding glass bottles of 100 ml.
Ruminal liquor of fastening animals was extracted through the cannula, with the help of a vacuum pump. The liquor was stored in a thermos to ensure adequate temperature conditions (39 °C) and anaerobiosis during transport to the laboratory.
Ruminal content of both animals was mixed and filtered through muslin. The resulting solid was added a small portion of buffer solution of Menke and Steingass (1988). This solution was agitated for a few seconds in a domestic blender to release microorganisms adhered to fiber. Later, filtrate of this portion was added to liquid fraction. Ruminal liquor was maintained in a CO2 atmosphere.
Each bottle was added 50 mL of a mixture of rumen liquor and buffer solution of Menke and Steingass (1988), in a 1:3 (v/v) proportion, and they were sealed with a butyl and agrafe stopper. Bottles without substrate were included as targets for correcting the effect of ruminal liquor in volumes of produced gas. All bottles were placed in a temperature controlled bath of 39 °C.
At 0 and 12 h of incubation, samples of each treatment were taken for identification and quantification by RT-PCR of populations of Fibrobacter succinogenes, Ruminococcus flavefaciens and total methanogens. Subsequently, they were placed in a freezer at -20 °C for further determination.
Values of Ct (threshold cycle), which is the cycle at which fluorescence is considered detectable in the exponential phase of PCR reaction, were obtained to determine the ΔCt, ΔΔCt, expression of these microbial populations related to control treatment and the percentage of cellulolytic and methanogenic bacteria in relation to total bacteria population.
Counting of protozoa, preserved in 10% of formaldehyde, was performed. They were counted with Neubauer chamber using an optical microscopy, after dyeing with a solution of gentian violet at 0.01% in glacial acetic acid.
DNA extraction. Cell disruption was achieved by freezing-thawing with liquid nitrogen. DNA was obtained using a matrix (glassmilk), joined to it and later released from it, when dissolved in double-distilled water (Makkar and McSweeney 2005). DNA concentration and sample purity (OD 260 nm/280 nm OD) were determined by absorbance at 260 nm and 280 nm in a Nanodrop 1000 spectrophotometer.
RT- PCR. Extracted DNA was amplified with pairs of primers for bacteria, fungi, R. flavefaciens, F. succinogenes and ruminal methanogens, according to the description of Denman and McSweeney (2006) and Denman et al. (2007).
DNA samples were taken to a final concentration of 10ng•µL-1 and amplification reaction was carried out with each of the specific primers (total bacteria, fungi, F. succinogenes, R. flavefaciens and methanogens) was performed. Primers were used at a concentration of 10mM and the SYBR Green 490 was also used. Final reaction volume was 25 µL.
DNA amplification was performed in an Applied Biosystem thermocycler, according to the following program: 1 cycle of 50 °C 2 min. and 95 °C 2 min., 40 cycles of 95 °C 15 sec. and 60 °C 1 min; 1 cycle of 95 °C 2 min., 60 °C 15 sec and 95 ° C 15 sec.
The amounts of studied microbial populations were expressed as proportion of total bacteria (ΔCt). These ΔCt values were calculated by the difference between the Ct value of target gene and the reference gene (16S rRNA of bacteria). The ΔΔCt was determined by the difference between the ΔCt of target groups from experimental diets and ΔCt of target groups from control diet. Percentage of F. succinogenes and R. flavefaciens, in relation to total bacterial population, was calculated from ΔCt values as 100 × (2ΔCt)-1 and the expression of target groups related to control treatment as 2-ΔΔCt (Denman and McSweeney 2006).
Experimental design and statistical analysis. A completely randomized design with 5x2 factorial arrangement (5 experimental diets and 2 h of sampling) was used. Four replications were carried out.
INFOSTAT statistical program was used for processing results, which was proposed by Di Rienzo et al. (2012). Multiple comparisons test of Duncan (1955) was applied on necessary cases.
RESULTS AND DISCUSSION
There were no significant interactions among experimental diets and hours of sampling for the studied variables, nor differences among individual factors (experimental diet and hours) for any of the selected indicators. Therefore, only the results of the effect of treatments on each variable are presented.
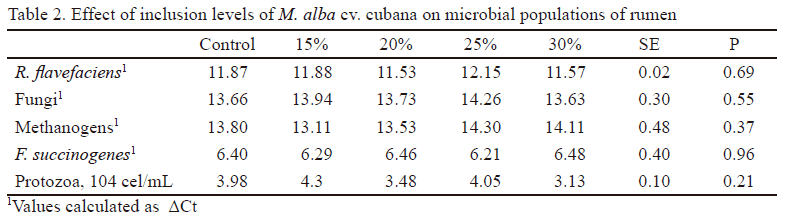
Currently, the characterization of complex microbial communities by molecular techniques is widely used and counting of microorganisms have been replaced by traditional culture methods (Saro et al. 2014). The study of ruminal microorganisms under the effect of different feeding strategies does not escape from this tendency. In this sense, real-time PCR has become a powerful tool that allows a fast quantification of these microorganisms. The design of specific primers has allowed their identification and quantification by this technique. In this experiment, identification and quantification of populations of microorganisms involved in fiber degradation in the rumen (F. succinogenes, R. flavefaciens and fungi) and total methanogens by RT-PCR (Table 2) indicated that the inclusion of M. alba cv cubana did not affect any of these ruminal populations. These results allowed to complement and corroborate those obtained by González et al. (2011), after evaluating the effect of four varieties of mulberry on the population of methanogens and cellulolytic microorganisms, quantified by culture techniques.
Secondary plant metabolites (saponins, flavonoids and tannins) have demonstrated the ability to manipulate ruminal fermentation in a favorable manner, which reduces the formation of methane in the rumen (Hu et al. 2005, Patra et al. 2006). Saponins reduce protozoa counts because the latter are more sensitive to changes caused by these compounds in their cell membranes (Moss et al. 2000). According to Goel et al. (2008), these compounds increase the proliferation of fiber-degrading bacteria and inhibit fungal population. Maldonado et al. (2000) and García (2003) determined saponin concentration in M. alba and found no presence of this compound in the plant. The absence of saponins in mulberry could then explain that no variations were found in populations of cellulolytic microorganisms (R. flavefaciens, F. succinogenes and fungi) and protozoa with the inclusion of this plant on the diet.
Condensed tannins present in some plants have demonstrated to be toxic to methanogens (Hess et al. 2003, Tavendale et al. 2005). In previous studies, González et al. (2010) determined the composition of condensed tannins of this mulberry variety and found no appreciable amounts of this compound. The fact that mulberry shows no saponins and tannins in its composition could explain that their inclusion on the diet had no negative effect on the population of methanogens.
Defaunation or reduction of protozoa is one of the methods used for reducing methanogens. It is stated that methanogens establish an ecto- and endosymbiotic relationship with rumen protozoa (Finlay et al. 1994, Ohene-Adjei et al. 2007). Many of methanogens are observed on the outer surface of ciliated protozoa from rumen. Therefore, when the number of protozoa decreases, generally methanogens are also reduced and, consequently, ruminal methane production diminishes (Kobayashi 2010). In this study, the inclusion of mulberry cv. cubana on the diet did not affect protozoa counts, so the amounts of methanogens were not affected either.
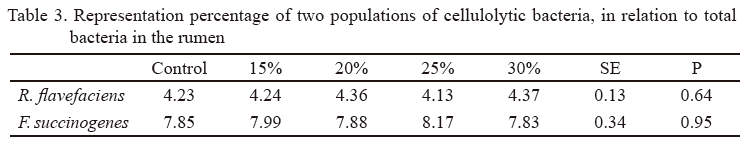
Table 3 shows that representation percentage of F. succinogenes and R. flavefaciens regarding total bacteria population was not affected by the inclusion of M. alba cv. cubana on diet. It is important to point out that in all treatments, F. succinogenes had a higher percentage of representation than R. flavefaciens. This result coincides with that obtained by McSweeney et al. (2007) and Hung et al. (2013), who listed some bacterial populations in the rumen of animals fed different diets and found that F. succinogenes was the predominant cellulolytic bacteria. The highest representation of F. succinogenes, found in this study, was favorable, taking into account that it is one of the cellulolytic bacteria with higher potentialities for fiber degradation (Kobayashi 2010). Therefore, animals could make better use of the fiber component of feeds.
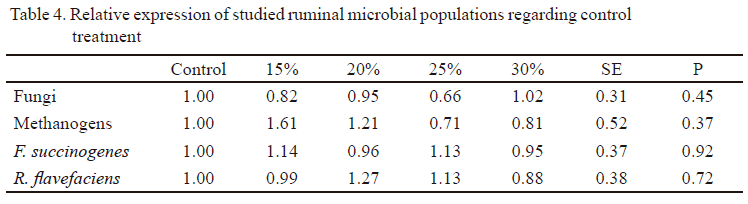
Values of ΔΔCt (figure 1) and the expression related to control treatment of ruminal microbial populations, identified and quantified (Table 4), demonstrated that all treatments including M. alba cv. cubana showed similar differences and expression related to control treatment for all microbial populations. These variables showed that inclusion levels of 15, 20, 25 and 30% of this Cuban variety in the ration did not affect populations of F. succinogenes, R. flavefaciens fungi and methanogens rumen.
Ruminal methanogens are a specialized group of microorganisms having the function of producing methane during the use of energy from food into the organ (Attwood et al. 2008, Liu et al. 2013, Chuntrakort et al. 2014, Khiaosa-ard & Zebeli 2014). In this study, the inclusion of this Cuban variety of mulberry did not reduce this microbial population numbers, so reductions of ruminal methane production should not be expected either. However, Gonzalez et al. (2012), when evaluating the same inclusion levels of this variety on the control of ruminal methanogenesis, found that all levels reduced methane production. Gonzalez et al. (2014), when evaluated the effect of these levels of mulberry on methanogen population by the molecular technique of denaturing gradient gel electrophoresis (DGGE), found no negative effects on these microorganisms. Therefore, it can be stated that reduction of methane production with the inclusion of 15, 20, 25 and 30% of M. alba cv. cubana is no because of direct effect of this plant on methanogens.
It is a favorable factor that studied cellulolytic populations within the rumen have not been affected by the inclusion of M. alba cv. cubana in the diet. Therefore, assuming the use of this variety as a strategy for controlling rumen methanogenesis, it would be carried out with the guarantee that it will not affect the main microbial populations degrading fiber nor their degradation process in the rumen.
Inclusion of M. alba cv. cubana on the diet does not affect methanogens or the main microbial populations that degrade the fiber in the rumen, which could be used as a strategy for reducing methane production in the rumen. The mechanism by which this plant reduces ruminal methanogenesis is not the direct effect on methanogens.
ACKNOWLEDGEMENTS
REFERENCES
Anbarasu, C., Dutta, N., Sharma, K. & Rawat, M. 2004. “Response of goats to partial replacement of dietary protein by a leaf meal mixture containing Leucaena leucocephala, Morus alba and Tectona grandis”. Small Ruminant Research, 51(1): 47–56, ISSN: 0921-4488, DOI: 10.1016/S0921-4488(03)00203-7.
AOAC. 2016. Official methods of analysis of AOAC International. 20th ed., Rockville, MD: AOAC International, ISBN: 978-0-935584-87-5, OCLC: 950056914, Available: <http://www.directtextbook.com/isbn/9780935584875>, [Consulted: September 22, 2016].
Attwood, G. T., Kelly, W. J., Altermann, E. H. & Leahy, S. C. 2008. “Analysis of the Methanobrevibacter ruminantium draft genome: understanding methanogen biology to inhibit their action in the rumen”. Australian Journal of Experimental Agriculture, 48(2): 83–88, ISSN: 0816-1089, DOI: 10.1071/EA07269.
Ba, N. X., Giang, V. D. & Ngoan, L. D. 2005. “Ensiling of mulberry foliage (Morus alba) and the nutritive value of mulberry foliage silage for goats in central Vietnam”. Livestock Research for Rural Development, 17(2), ISSN: 0121-3784.
Bakshi, M. P. S. & Wadhwa, M. 2007. “Tree leaves as complete feed for goat bucks”. Small Ruminant Research, 69(1–3): 74–78, ISSN: 0921-4488, DOI: 10.1016/j.smallrumres.2005.12.009.
Chuntrakort, P., Otsuka, M., Hayashi, K., Takenaka, A., Udchachon, S. & Sommart, K. 2014. “The effect of dietary coconut kernels, whole cottonseeds and sunflower seeds on the intake, digestibility and enteric methane emissions of Zebu beef cattle fed rice straw based diets”. Livestock Science, 161: 80–89, ISSN: 1871-1413, DOI: 10.1016/j.livsci.2014.01.003.
Delgado, D. C., González, R., Galindo, J., Cairo, J. & Almeida, M. 2007. “Potencialidad de Trichantera gigantea y Morus alba para reducir la producción ruminal de metano in vitro”. Cuban Journal of Agricultural Science, 41(4): 339–342, ISSN: 2079-3480.
Denman, S. E. & McSweeney, C. S. 2006. “Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen: Real-time PCR assay of the rumen anaerobic fungal population”. FEMS Microbiology Ecology, 58(3): 572–582, ISSN: 0168-6496, 1574-6941, DOI: 10.1111/j.1574-6941.2006.00190.x.
Denman, S. E., Tomkins, N. W. & McSweeney, C. S. 2007. “Quantitation and diversity analysis of ruminal methanogenic populations in response to the antimethanogenic compound bromochloromethane: Monitoring of rumen methanogenic Archaea”. FEMS Microbiology Ecology, 62(3): 313–322, ISSN: 0168-6496, 1574-6941, DOI: 10.1111/j.1574-6941.2007.00394.x.
Di Rienzo, J. A., Casanoves, F., Balzarini, M. G., González, L., Tablada, M. & Robledo, C. W. 2012. InfoStat. version 2012, [Windows], Universidad Nacional de Córdoba, Argentina: Grupo InfoStat, Available: <http://www.infostat.com.ar/>.
Duncan, D. B. 1955. “Multiple Range and Multiple F Tests”. Biometrics, 11(1): 1–42, ISSN: 0006-341X, DOI: 10.2307/3001478.
Finlay, B. J., Esteban, G., Clarke, K. J., Williams, A. G., Embley, T. M. & Hirt, R. P. 1994. “Some rumen ciliates have endosymbiotic methanogens”. FEMS Microbiology Letters, 117(2): 157–161, ISSN: 0378-1097, 1574-6968, DOI: 10.1111/j.1574-6968.1994.tb06758.x.
Foiklang, S., Wanapat, M. & Toburan, W. 2011. “Effects of various plant protein sources in high-quality feed block on feed intake, rumen fermentation, and microbial population in swamp buffalo”. Tropical Animal Health and Production, 43(8): 1517–1524, ISSN: 0049-4747, 1573-7438, DOI: 10.1007/s11250-011-9836-y.
García, D. 2003. Efecto de los principales factores que influyen en la composición fitoquímica de Morus alba (Linn.). Master Thesis, Universidad de Matanzas ‘Camilo Cienfuegos’ - EEPF ‘Indio Hatuey’, Matanzas, Cuba.
Goel, G., Makkar, H. P. S. & Becker, K. 2008. “Changes in microbial community structure, methanogenesis and rumen fermentation in response to saponin-rich fractions from different plant materials”. Journal of Applied Microbiology, 105(3): 770–777, ISSN: 1364-5072, 1365-2672, DOI: 10.1111/j.1365-2672.2008.03818.x.
Goering, H. K. & Van Soest, P. J. 1970. Forage fiber analysis: apparatus, reagents, procedures, and some applications. (ser. Agriculture Handbook, no. ser. 379), Washington, D.C.: Agricultural Research Service, U.S. Dept. of Agriculture, 20 p., OCLC: 13295375.
González, N., Galindo, J., Aldana, A. I., Moreira, O. & Sarduy, L. R. 2011. “Effect of four mulberry (Morus alba Linn.) varieties on microbial population and fermentative products with rumen liquid from river buffaloes (Bubalus bubalis) under in vitro conditions”. Cuban Journal of Agricultural Science, 45(4): 399–404, ISSN: 2079-3480.
González, N., Galindo, J., Aldana, A. I., Moreira, O. & Sarduy, L. R. 2012. “Effect of different inclusion levels of Morus alba Linn cv. cubana on the methane fermentation and production under in vitro conditions with rumen liquor from river buffaloes (Bubalus bubalis)”. Cuban Journal of Agricultural Science, 46(2): 151–157, ISSN: 2079-3480.
González, N., Galindo, J., Aldana, A. I., Moreira, O., Sarduy, L. R., Abdalla, L. A. & Santos, M. R. 2010. “Evaluation of different varieties of mulberry (Morus alba) in the control of the methanogenesis in buffalo rumen liquid”. Cuban Journal of Agricultural Science, 44(1): 37–41, ISSN: 2079-3480.
González, N., Galindo, J., Navarrete, A. A., Abdalla, A. L. & Tsai, S. M. 2014. “Determination of the effect of Morus alba cv. cubana on the population of methanogens within the rumen liquor of water buffaloes, using the Denaturing Gradient Gel Electrophoresis (DGGE)”. Cuban Journal of Agricultural Science, 48(3): 253–257, ISSN: 2079-3480.
Hess, H. D., Kreuzer, M., Dı́az, T. E., Lascano, C. E., Carulla, J. E., Soliva, C. R. & Machmüller, A. 2003. “Saponin rich tropical fruits affect fermentation and methanogenesis in faunated and defaunated rumen fluid”. Animal Feed Science and Technology, 109(1–4): 79–94, ISSN: 0377-8401, DOI: 10.1016/S0377-8401(03)00212-8.
Hu, W. L., Liu, J. X., Ye, J. A., Wu, Y. M. & Guo, Y. Q. 2005. “Effect of tea saponin on rumen fermentation in vitro”. Animal Feed Science and Technology, 120(3–4): 333–339, ISSN: 0377-8401, DOI: 10.1016/j.anifeedsci.2005.02.029.
Hung, L. V., Wanapat, M. & Cherdthong, A. 2013. “Effects of Leucaena leaf pellet on bacterial diversity and microbial protein synthesis in swamp buffalo fed on rice straw”. Livestock Science, 151(2–3): 188–197, ISSN: 1871-1413, DOI: 10.1016/j.livsci.2012.11.011.
Kandylis, K., Hadjigeorgiou, I. & Harizanis, P. 2009. “The nutritive value of mulberry leaves (Morus alba) as a feed supplement for sheep”. Tropical Animal Health and Production, 41(1): 17–24, ISSN: 0049-4747, 1573-7438, DOI: 10.1007/s11250-008-9149-y.
Khiaosa-ard, R. & Zebeli, Q. 2014. “Cattle’s variation in rumen ecology and metabolism and its contributions to feed efficiency”. Livestock Science, 162: 66–75, ISSN: 1871-1413, DOI: 10.1016/j.livsci.2014.01.005.
Kobayashi, Y. 2010. “Abatement of Methane Production from Ruminants: Trends in the Manipulation of Rumen Fermentation”. Asian-Australasian Journal of Animal Sciences, 23(3): 410–416, ISSN: 1011-2367, 1976-5517, DOI: 10.5713/ajas.2010.r.01.
Liu, C., Zhu, Z. P., Shang, B., Chen, Y. X., Guo, T. J. & Luo, Y. M. 2013. “Long-term effects of ensiled cornstalk diet on methane emission, rumen fermentation, methanogenesis and weight gain in sheep”. Small Ruminant Research, 115(1–3): 15–20, ISSN: 0921-4488, DOI: 10.1016/j.smallrumres.2013.07.011.
Makkar, H. P. S. & McSweeney, C. S. (eds.). 2005. Methods in Gut Microbial Ecology for Ruminants. Berlin-Heidelberg: Springer-Verlag, ISBN: 978-1-4020-3790-0, Available: <http://link.springer.com/10.1007/1-4020-3791-0>, [Consulted: September 22, 2016].
Maldonado, M., Grande, D., Aranda, E. & Pérez-Gil, F. 2000. “Evaluación de árboles forrajeros tropicales para la alimentación de rumiantes en Tabasco, México”. In: IV Taller Internacional Silvopastoril ‘Los árboles y arbustos en la ganadería tropical’, Matanzas, Cuba: EEPF ‘Indio Hatuey’, pp. 135–142, ISBN: 978-959-16-0285-5.
McAllister, T. A. & Newbold, C. J. 2008. “Redirecting rumen fermentation to reduce methanogenesis”. Australian Journal of Experimental Agriculture, 48(2): 7, ISSN: 0816-1089, DOI: 10.1071/EA07218.
McSweeney, C. S., Denman, S. E., Wright, A. D. G. & Yu, Z. 2007. “Application of Recent DNA/RNA-based Techniques in Rumen Ecology”. Asian-Australasian Journal of Animal Sciences, 20(2): 283–294, ISSN: 1011-2367, 1976-5517, DOI: 10.5713/ajas.2007.283.
Menke, K. H. & Steingass, H. 1988. “Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid”. Animal Research and Development, 28(1): 7–55, ISSN: 0340-3165.
Moss, A. R., Jouany, J. P. & Newbold, J. 2000. “Methane production by ruminants: its contribution to global warming”. Annales de Zootechnie, 49(3): 231–253, ISSN: 0003-424X, 1297-9651, DOI: 10.1051/animres:2000119.
Ohene-Adjei, S., Teather, R. M., Ivan, M. & Forster, R. J. 2007. “Postinoculation Protozoan Establishment and Association Patterns of Methanogenic Archaea in the Ovine Rumen”. Applied and Environmental Microbiology, 73(14): 4609–4618, ISSN: 0099-2240, DOI: 10.1128/AEM.02687-06.
Patra, A. K., Kamra, D. N. & Agarwal, N. 2006. “Effect of plant extracts on in vitro methanogenesis, enzyme activities and fermentation of feed in rumen liquor of buffalo”. Animal Feed Science and Technology, 128(3–4): 276–291, ISSN: 0377-8401, DOI: 10.1016/j.anifeedsci.2005.11.001.
Saro, C., Ranilla, M. J., Tejido, M. L. & Carro, M. D. 2014. “Influence of forage type in the diet of sheep on rumen microbiota and fermentation characteristics”. Livestock Science, 160: 52–59, ISSN: 1871-1413, DOI: 10.1016/j.livsci.2013.12.005.
Soliva, C. R., Hess, H. D., Meile, L., Kreuzer, M. & Machmüller, A. 2003. “Suppression of ruminal methanogenesis by dietary means: apparent inconsistency between methane formation and counts of microbes involved in methanogenesis”. Tropical and Subtropical Agroecosystems, 3: 209–213, ISSN: 1870-0462.
Tan, N. D., Wanapat, M., Uriyapongson, S., Cherdthong, A. & Pilajun, R. 2012. “Enhancing Mulberry Leaf Meal with Urea by Pelleting to Improve Rumen Fermentation in Cattle”. Asian-Australasian Journal of Animal Sciences, 25(4): 452–461, ISSN: 1011-2367, 1976-5517, DOI: 10.5713/ajas.2011.11270.
Tavendale, M. H., Meagher, L. P., Pacheco, D., Walker, N., Attwood, G. T. & Sivakumaran, S. 2005. “Methane production from in vitro rumen incubations with Lotus pedunculatus and Medicago sativa, and effects of extractable condensed tannin fractions on methanogenesis”. Animal Feed Science and Technology, 123–124: 403–419, ISSN: 0377-8401, DOI: 10.1016/j.anifeedsci.2005.04.037.
Theodorou, M. K., Williams, B. A., Dhanoa, M. S., McAllan, A. B. & France, J. 1994. “A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds”. Animal Feed Science and Technology, 48(3–4): 185–197, ISSN: 0377-8401, DOI: 10.1016/0377-8401(94)90171-6.
Received: 29/3/2016
Accepted: 16/9/2016
Niurca González, Instituto de Ciencia Animal. Apartado Postal 24, San José de las Lajas, Mayabeque, Cuba . Email: ngonzalez@ica.co.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}