










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCuban Journal of Agricultural Science
versión On-line ISSN 2079-3480
Cuban J. Agric. Sci. vol.51 no.1 Mayabeque ene.-mar. 2017
ARTÍCULO ORIGINAL
Metabolitos secundarios de Leucaena leucocephala. Su relación con algunos elementos del clima, diferentes expresiones de digestibilidad y metabolitos primarios
Secondary metabolites of Leucaena leucochephala. Their relationship with some climate elements, different expressions of digestibility and primary metabolites
R. S. Herrera1, D. M. Verdecia2, J. L. Ramírez2, M. García1 and Ana M. Cruz1
1Instituto de Ciencia Animal, Apartado Postal 24, San José de las Lajas, Mayabeque, Cuba
2Universidad de Granma, Carretera de Manzanillo km 17½, Universidad de Granma, Bayamo, Cuba
RESUMEN
Se establecieron los coeficientes de correlación de Pearson entre los metabolitos secundarios de Leucaena leucocephala y algunos factores climáticos (temperatura máxima, mínima y media, lluvia y número de días con lluvia), los metabolitos primarios y las digestibilidades (materia orgánica, materia seca, proteína y FDN). Los metabolitos fueron A) secundarios: taninos totales, fenoles totales, taninos condensados totales, taninos condensados totales, taninos condensados libres, alcaloides, saponinas, triterpenos, esteroides y flavonoides) y B) primarios: glucosa, fructosa, sacarosa y N. Los metabolitos secundarios se correlacionaron con la temperatura máxima y el mayor coeficiente (r=0.78) fue para los esteroides, similar resultado se obtuvo para los taninos condensados ligados a la fibra, pero con la temperatura mínima (r= -0.90). El mayor coeficiente para el número de días con lluvias se presentó en los flavonoides (r=0.99), mientras que para la lluvia total fue para los fenoles totales (r= -0.90). Los coeficientes de correlación entre los metabolitos secundarios y la DMO variaron entre -0.57 y -0.74. Para la DIVMS fluctuaron entre -0.65 y -0.94; para FDN entre -0.91 y -0.98, mientras que para la DPB fueron los menores (-0.39 y 0.55). Al considerar los compuestos fenólicos como variables independientes y relacionarlos con los alcaloides, saponinas, triterpenos, esteroides y flavonoides, los mayores coeficientes (≥0.80) lo registraron lo taninos totales, fenoles totales, taninos condensados totales y taninos condensados ligados a la fibra. Al relacionar los metabolitos secundarios con los metabolitos primarios (glucosa, fructosa, sacarosa y N), los coeficientes fueron negativos y variaron entre -0.41 y -0.98. Se concluye que los elementos del clima influyeron en los metabolitos estudiados. Todas las expresiones de digestibilidad se afectaron negativamente por la presencia de los metabolitos secundarios. Los compuestos fenólicos se correlacionaron positivamente con los flavonoides, alcaloides, saponinas, triterpenos y esteroides, mientras que los metabolitos primarios se correlacionaron negativamente con los metabolitos secundarios. Se recomienda considerar los resultados de este estudio en Leucaena leucocephala para valorar el efecto del cambio climático en los metabolitos secundarios; contrarrestar el efecto negativo de estos compuestos en los sistemas de alimentación de rumiantes; realizar investigaciones futuras que permitan establecer su valor máximo permisible en la dieta y aplicar la simulación y modelación matemática para predecir el comportamiento de estas sustancias en esta y otras plantas.
Palabras clave: factores antinutricionales, digestibilidad, leguminosas, temperatura, lluvia
ABSTRACT
Pearson correlation coefficients were established among secondary metabolites of Leucaena leucocephala and some climatic factors (maximum, minimum and mean temperature, rainfall and number of rainy days), primary metabolites and digestibility (organic matter, dry matter, protein and NDF). The metabolites were A) secondary: total tannins, total phenols, total condensed tannins, free condensed tannins, alkaloids, saponins, triterpenes, steroids and flavonoids and B) primary: glucose, fructose, sucrose and N. Secondary metabolites were correlated with the maximum temperature and the highest coefficient (r = 0.78) was for steroids. Similar result was obtained for condensed tannins bound to the fiber, but with the minimum temperature (r = -0.90). The highest coefficient for the number of rainy days was presented in flavonoids (r = 0.99), while for total rainfall coefficient, it was for total phenols (r = -0.90). Correlation coefficients between the secondary metabolites and OMD ranged from -0.57 to -0.74. For the in vitro of dry matter digestibility, they ranged between -0.65 and -0.94 and for NDF between -0.91 and -0.98. For PBD, the values were the lowest (-0.39 and 0.55). Considering the phenolic compounds as independent variables and related to alkaloids, saponins, triterpenes, steroids and flavonoids, the highest coefficients (≥0.80) were recorded by total tannins, total phenols, total condensed tannins and condensed tannins bound to fiber. When the secondary metabolites were related to the primary metabolites (glucose, fructose, sucrose and N), coefficients were negative and varied between -0.41 and -0.98. It can be concluded that the elements of the climate influenced the studied metabolites. All digestibility expressions were negatively affected by the presence of the secondary metabolites. Phenolic compounds were positively correlated with flavonoids, alkaloids, saponins, triterpenes and steroids, while primary metabolites correlated negatively with secondary metabolites. It is recommended to consider the results of this study in Leucaena leucocephala to assess the effect of climate change on the secondary metabolites, counteract the negative effect of these compounds on ruminant feeding systems, conduct future research to establish its maximum allowable value in the diet and apply the simulation and mathematical modeling to predict the performance of these substances in this and other plants.
Key words: antinutritional factors, digestibility, legumes, temperature, rain
INTRODUCCIÓN
Los pastos de gramíneas, leguminosas, árboles y arbustos desempeñan una función protagónica en la alimentación animal por la calidad de la biomasa así como alternativa para reducir el uso de los alimentos concentrados debido a sus altos precios (Díaz 2014 y Friedrich 2014). No obstante, es necesario dominar su composición química y sus fuentes de variación (Boufennara et al. 2012).
Entre las leguminosas arbóreas, Leucaena leucocephala tiene gran difusión en el trópico, debido a las múltiples ventajas de su utilización. Esto permitió establecer sistemas de utilización que favorecen la producción de leche y carne, económicamente viable y amigable con el medio ambiente (Ruiz et al. 2008). Sin embargo, la mayoría de los estudios han centrado su atención en el contenido de proteína, fibra, minerales y digestibilidad (Wencomo y Ortiz 2012), mientras que son escasos los relacionados con los metabolitos secundarios (Pedraza 2008).
Estos metabolitos secundarios están integrados por una amplia gama de compuestos químicos (taninos, fenoles, alcaloides y saponinas, entre otros) que pueden producir variados efectos en el crecimiento y productividad del animal, en dependencia de su concentración y del tiempo de ingestión, entre otros factores (Semmar et al. 2011).
Amplia es la literatura que señala la variación de la composición química, de las gramíneas y leguminosas, con respecto a los componentes del manejo y es frecuente que se presenten estos resultados de acuerdo con la estación climática (período lluvioso y poco lluvioso), edad de rebrote y fertilización, entre otros aspectos. Sin embargo, esta información no es vasta en los metabolitos secundarios de las leguminosas y, en especial, en su relación con los factores que integran el clima.
Sobre la base de estos elementos antes citados, el objetivo de este trabajo fue establecer el coeficiente de correlación de Pearson entre: a) los metabolitos secundarios de Leucaena leucocephala y algunos factores climáticos, b) las diferentes expresiones de digestibilidad y c) los metabolitos primarios, glucosa, fructosa, sacarosa y N.
MATERIALES Y MÉTODOS
Localización, suelo y clima. La investigación se realizó en el área experimental de la Universidad de Granma, Cuba, en suelo pardo con carbonato (Hernández et al. 1999), pH 6.2, contenidos de P2O5, K2O y N de 2.4, 33.4 y 3.0 mg/100g de suelo, respectivamente, y 3.6 % de materia orgánica. El experimento tuvo dos años de duración y se cuantificaron las temperaturas máxima, mínima y media, lluvia y número de día con lluvia.
Procedimiento y muestreo. Se empleó la Leucaena leucocephala con dos años de establecida, en parcelas de 0.5 ha. Los muestreos se realizaron cada 60, 120 y 180 d de rebrote en 10 plantas al azar. En cada planta se simuló el ramoneo del animal, considerando las hojas, los pecíolos y los tallos con diámetro inferior a dos centímetros. Todo el material muestreado en cada planta se mezcló hasta obtener una muestra homogénea, se secó en estufa de circulación de aire forzado durante 72 horas a 65 ºC. Las muestras se molinaron a tamaño de partícula de 1 mm.
Análisis químico de las muestras. Se determinó el contenido de proteína bruta (ADAC 2016); fenoles y taninos totales por el método de Folin-Ciocalteu, antes y después del tratamiento de los extractos con polivinilpolipirrolidona (Makkar et al. 1988); taninos condesados, libres y ligados a la fibra con la utilización de nbutanol/HCl/Fe3+ (Porter et al. 1985); flavonoides según Bohm y Koupai-Abyazani (1994); saponinas, de acuerdo con Obadoni y Ochuko (2002); alcaloides como lo señalan Muzquiz et al. (1994); triterpenos según Jie-Ping y Chao-Hong (2006) y esteroides mediante Galindo et al. (1989).
Para la cuantificación de los azúcares, se utilizó un HPLC (WATERS) de 2410 detector de índice de refracción, con la utilización del programa Empower Pro 2002. El método cromatográfico adoptado fue el isocrático (flujo constante), de 1 mL/min con fase móvil de agua + ácido sulfúrico 0.01 N y 50 ºC de temperatura de columna. La columna utilizada fue exclusión iónica BioRad Amino HPX-87M de 300 mm x 7.8 mm.
La digestibilidad in vitro se cuantificó en el incubador DaisyII® (ANKOM 2000). Para la degradabilidad in situ, se utilizó el método de Mehrez y Ørskov (1977) con la utilización de tres ovejas provistas de cánula ruminal permanente. Se utilizaron bolsas (15 cm x 10 cm) para las incubaciones con 45 µm de tamaño de poro. A las 72 h, las bolsas se extrajeron, se lavaron con agua fría, se secaron en estufa de aire forzado a 53 ºC hasta alcanzar peso constante y se pesaron. Los residuos se procesaron en el analizador de fibra ANKOM (2000) para determinar la digestibilidad de la FDN y FDA. Se determinó el N (método Kjeldhal) para establecer la digestibilidad de la PB y la de la materia orgánica se estableció según Aumont et al. (1995).
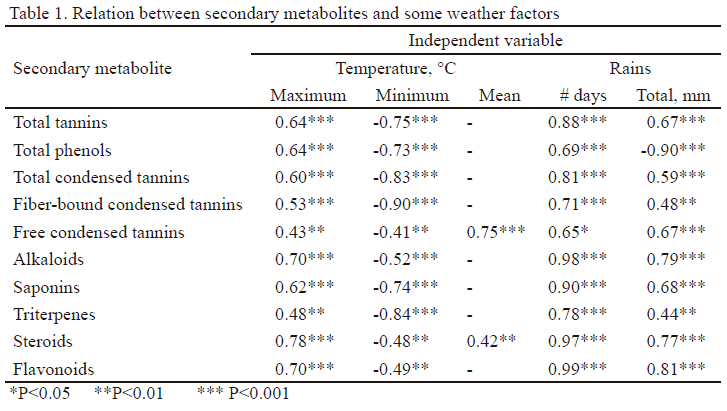
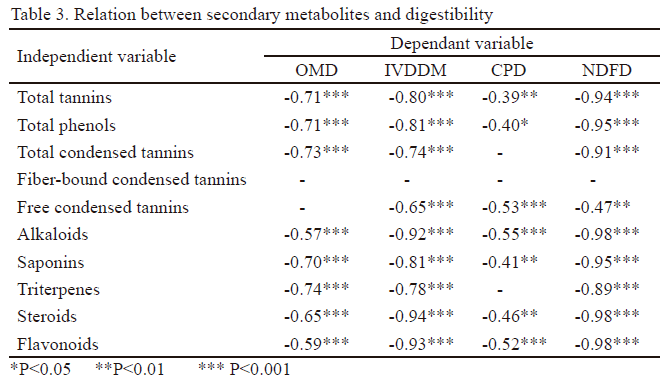
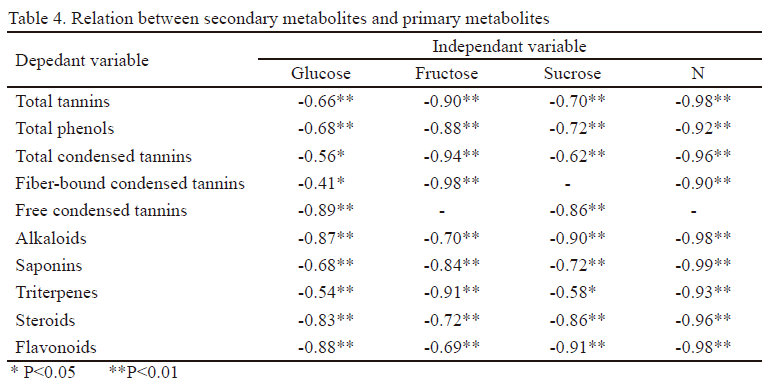
Análisis estadístico. Se determinaron los coeficientes de correlación de Pearson (Visauta 2007) entre los metabolitos secundarios (fenoles totales, taninos totales, taninos condensados ligados a la fibra, taninos condensados libres) y: a) algunos factores climáticos (temperatura máxima, mínima y media, número de días con lluvias y lluvia total) b) las digestibilidades de la MO, MS, PB y FDN y c) los metabolitos primarios (glucosa, fructosa, sacarosa y N). Además, se establecieron las relaciones entre los fenoles totales, taninos totales, taninos condensados ligados a la fibra, taninos condensados libres y los alcaloides, saponinas, triterpenos, esteroides y flavonoides. Solo se informan los coeficientes mayores de ± 0.40, con probabilidad de 95 % (Andrade et al. 2015).
RESULTADOS
Los metabolitos secundarios se correlacionaron positivamente con la temperatura máxima (0.43-0.78), mientras que esta fue negativa (0.41-0.90) con la temperatura mínima y no hubo correlación con la media para los fenoles totales, taninos totales, taninos condensados totales, taninos condensados ligados a la fibra, saponinas y triterpenos. El coeficiente de correlación fue positivo con el número de días con lluvias y varió entre 0.65 y 0.99, mientras que con la lluvia total fue variable (tabla 1).
Los taninos totales y fenoles totales mostraron altas y positivas correlaciones con los flavonoides, alcaloides, saponinas, triterpenos y esteroides, mientras que para los taninos condensados totales, taninos condensados ligados a la fibra y taninos condensados libres, las correlaciones tuvieron menor magnitud (tabla 2).
Los metabolitos secundarios se correlacionaron negativamente con las digestibilidades de la materia orgánica (DMO), in vivo de la materia seca (DIVMS), de la proteína (DPB) y de la FDN (DFDN), pero la mayor magnitud se registró en esta última con coeficientes de -0.98, con excepción de los taninos condensados libres, donde el coeficiente fue solo de -0.47 (tabla 3).
Se encontraron correlaciones negativas entre los metabolitos secundarios y los metabolitos primarios glucosa, fructosa, sacarosa y N. El mayor coeficiente se encontró para el N y, en general, el menor valor se obtuvo para la glucosa (tabla 4).
DISCUSIÓN
Se considera que existen, aproximadamente, 24000 compuestos químicos en el reino vegetal que tienen efectos tóxicos y antinutricionales, denominados metabolitos secundarios. Se encuentran ampliamente distribuidos y las leguminosas portan muchas de estas sustancias.
En la actualidad, sus funciones en el reino vegetal están mucho más esclarecidas que hace diez años. Se hallan referidas como un medio de defensa contra herbívoros, reguladores de la simbiosis, controladoras de la germinación de la semilla, efectos alelopáticos, intervienen directamente en la relación planta-animal y contribuyen a la adaptación de las plantas a las condiciones ambientales imperantes, entre otras múltiples funciones (Makkar et al. 2007).
Además, se ha informado que existe un grupo de sustancias generadas por el metabolismo natural de las especies vegetales y que, por diferentes mecanismos, ejercen efectos contrarios a la óptima nutrición de los animales cuando las consumen, debido a sus variados efectos en los procesos digestivos y metabólicos (Pérez et al. 2013). Sin embargo, no se debe perder de vista que la denominación que caracteriza su naturaleza antinutricional se adoptó a partir del enfoque de recurso para la alimentación de herbívoros, y no de las funciones que cumplen en los tejidos de los vegetales.
Todo parece indicar que el incremento de la temperatura máxima favorece la síntesis de los metabolitos secundarios, al igual que cuando disminuye la temperatura mínima. Esto pudiera estar relacionado con el papel que ellos desempeñan como agentes protectores de las plantas contra depredadores o condiciones climáticas adversas (Ryan et al. 2010). Este criterio se reafirma con los resultados de la presente investigación, pero se debe adicionar que mientras menores sean las temperaturas mínimas, mayor es la síntesis de estos compuestos.
Los procesos bioquímicos y fisiológicos, relacionados con la síntesis, transporte y degradación de sustancias en las plantas, están influenciados por el régimen térmico en el que se desarrollan, el grado de relación que estas poseen con la cinética de las reacciones bioquímicas y el mantenimiento de la integridad de las membranas (Murray y Keely 2009). Lo anterior se evidencia en la baja correlación o ausencia de ella entre los metabolitos secundarios y la temperatura media.
En Cuba, el régimen térmico se caracteriza por relativa homogeneidad en las llanuras y variaciones asociadas a la altitud. Sin embargo, las oscilaciones diarias superan a la anual y, por tanto, resulta de interés el comportamiento de la temperatura máxima y mínima, así como la diferenciación entre los períodos que caracterizan cada etapa estacional, con respecto al comportamiento de los compuestos químicos, en especial de los metabolitos secundarios de las especies y variedades.
No todas las especies forrajeras tienen el mismo valor óptimo de temperatura para el cumplimiento de estas funciones. Ramírez (2010) informó que el óptimo fotosintético para las leguminosas se encuentra entre 30 y 35 ºC, con alta sensibilidad a las bajas temperaturas, cuyos efectos negativos en el crecimiento ocurren entre 0 y 15 ºC, aunque en algunas especies comienza a partir de 20 ºC, si la humedad no es un factor limitante. Lo anterior se atribuye a la baja conversión de azúcares en los tejidos de las plantas, debido a la disminución de los procesos de biosíntesis y déficit energético por la reducción de la tasa respiratoria. Lo anterior se puede avalar con los resultados que se muestran en la tabla 4, donde las correlaciones fueron negativas entre los metabolitos secundarios y los primarios.
La conversión de energía solar en biomasa por las gramíneas y leguminosas es variable, y depende de las vías metabólicas mediante las que se efectúa la fotosíntesis, por lo que la tasa fotosintética es una función de la energía disponible. En las plantas C3, sometidas a altas intensidades de radiación, se afecta la tasa de asimilación neta y el estado de saturación lumínica se alcanza en sus hojas a 300-400 J/m2/seg en condiciones controladas. Sin embargo, en condiciones normales de explotación, las hojas superiores reducen los valores de radiación al resto del follaje y sólo una parte de ellas alcanza su potencial fotosintético.
Las leguminosas son portadoras del ciclo fotosintético C3 y esto determina que su etapa de crecimiento inicial sea lenta, si se compara con las gramíneas forrajeras de sendero C4. De modo que no se han realizado suficientes estudios que establezcan que las leguminosas arbustivas y las plantas arbóreas sean portadoras del referido ciclo fotosintético, a pesar de existir evidencias de que los factores climáticos influyen en su composición química ni aquellos que relacionan la vía fotosintética con los metabolitos secundarios.
El volumen de agua caída por las precipitaciones y su distribución en el año ejercen efectos en el crecimiento y calidad de los forrajes, debido a su relación con los factores bioquímicos y fisiológicos que regulan estos procesos biológicos de gran complejidad. El agua es un componente esencial de las células de las plantas, casi todos los procesos metabólicos dependen de su presencia. Además, se requiere para el mantenimiento de la presión de turgencia, la difusión de solutos en las células y suministra el hidrógeno y oxígeno que están involucrados durante el proceso fotosintético. De ahí, la importancia de considerar no solo el volumen de las precipitaciones sino su distribución a través del tiempo.
Los resultados de esta investigación señalaron que desde el punto de vista de los metabolitos secundarios es más importante la distribución de las lluvias (número de días con lluvia) que el volumen total de ellas. Esto puede estar relacionado con la existencia de cantidad de agua suficiente, que permite la correcta turgencia celular, adecuado balance hídrico y disponibilidad de nutrientes en la célula, lo que propicia la síntesis de metabolitos secundarios.
Si los metabolitos secundarios se encuentran en determinadas concentraciones en la MS, al ser ingeridos por los animales pueden causar efecto negativo en su fisiologismo, trastornos neurológicos, reproductivos y digestivos, papera, gangrena e incluso la muerte. Entre los metabolitos causantes de estos efectos, se encuentran los alcaloides, glicósidos cianogénicos, aminoácidos tóxicos y saponinas, entre otros. Si su concentración es superior al 2 % de la MS, producen trastornos digestivos y en órganos sensoriales, relacionados con la alimentación y disminución de la acción de los microorganismos ruminales en la digestibilidad de la pared celular, entre otros. Los metabolitos causantes de estos efectos son, principalmente, los taninos y los inhibidores de proteasas y amilasas (Makkar et al. 2007 y Newell-McGloughlin 2008).
Los taninos hidrolizables pueden causar hemorragias gástricas y daños en los riñones. Los taninos condensados están relacionados con la mejor digestión de las proteínas en los rumiantes y los pueden proteger contra la helmintiasis (Makkar 2003). Además, Makkar et al. (2007) informaron que los metabolitos secundarios han despertado el interés en alimentos para consumo humano, debido a su potencial para prevenir cáncer, desórdenes circulatorios e infecciones virales, antioxidantes y efectos antibacterias, antivirus y antiparásitos, entre otras propiedades. Sin embargo, sus mecanismos de acción no están totalmente establecidos.
Las correlaciones negativas informadas en este trabajo entre los metabolitos secundarios y las diferentes expresiones de digestibilidad informaron que, mientras menor es el contenido de los primeros, mayores valores de digestibilidad se obtienen, en especial la digestibilidad de la FDN. Esto pudiera indicar mayor degradabilidad de esta fracción fibrosa y, por lo tanto, mayor energía disponible. Llamó la atención que la menos influida fue la digestibilidad de la PB, lo que se pudiera atribuir al efecto protector de los taninos.
Con anterioridad se evidenció la importancia de los metabolitos primarios. En este estudio, se encontraron correlaciones negativas de los metabolitos secundarios con la glucosa, fructosa, sacarosa y N. Esto pudiera indicar que los metabolitos primarios forman parte de la estructura de los metabolitos secundarios, como son las cadenas carbonadas, y están involucrados en la síntesis de los referidos metabolitos como fuentes de energía. Por ello, se debe atender a estos aspectos, ya que los metabolitos primarios desempeñan una función fundamental en el crecimiento, desarrollo y reproducción de las plantas, lo que implicaría alteraciones en los diferentes procesos fisiológicos, al ser empleados los metabolitos primarios en la síntesis de los metabolitos secundarios. En estudios futuros estos elementos necesitan ser confirmados.
En la bibliografía consultada no se encontraron artículos que hicieran referencia a los resultados aquí presentados, ya que en términos generales la composición química de la leucaena se ha informado por período estacional (período lluvioso y poco lluvioso). Igual situación se ha notificado en relación con los metabolitos secundarios (Verdecia 2014). Importante fue el efecto de la temperatura máxima y mínima, y la distribución de las precipitaciones en los metabolitos secundarios, así como la acción negativa de ellos en las diferentes expresiones de digestibilidad, pero la menor influencia fue para los taninos y fenoles en relación con la digestibilidad de la PB, así como su relación con los metabolitos primarios. Lo anterior necesita investigaciones específicas futuras para avalar y explicar con mayor profundidad los resultados obtenidos.
Se concluye que los elementos del clima influyeron en los metabolitos estudiados. Todas las expresiones de digestibilidad se afectaron negativamente por la presencia de los metabolitos secundarios. Los compuestos fenólicos se correlacionaron positivamente con los flavonoides, alcaloides, saponinas, triterpenos y esteroides, mientras que los metabolitos primarios se correlacionaron negativamente con los metabolitos secundarios. Se recomienda considerar los resultados de este estudio en Leucaena leucocephala para valorar el efecto del cambio climático en los metabolitos secundarios y contrarrestar el efecto negativo de estos compuestos en los sistemas de alimentación de rumiantes. Se sugiere además, realizar investigaciones futuras que permitan establecer su valor máximo permisible en la dieta y aplicar la simulación y modelación matemática para predecir el comportamiento de estas sustancias en esta y otras plantas.
AGRADECIMIENTOS
Los autores agradecen la Universidad de León, España por la colaboración en el análisis químico de las muestras
REFERENCIAS
Andrade, A. S., Santos, P. M., Pezzopane, J. R. M., Araujo, L. C., Pedreira, B. C., Pedreira, C. G. S., Marin, F. R. & Lara, M. a. S. 2015. “Simulating tropical forage growth and biomass accumulation: an overview of model development and application”. Grass and Forage Science, 71(1): 54–65, ISSN: 1365-2494, DOI: 10.1111/gfs.12177.
ANKOM. 2000. Procedures for fiber and in vitro analysis. Ankom Technology, Available: <http://www.ankom.com>, [Consulted: November 16, 2010].
AOAC. 2016. Official methods of analysis of AOAC International. 20th ed., Rockville, MD: AOAC International, ISBN: 978-0-935584-87-5, Available: <http://www.directtextbook.com/isbn/9780935584875>, [Consulted: September 22, 2016].
Aumont, G., Caudron, I., Saminadin, G. & Xande´, A. 1995. “Sources of variation in nutritive values of tropical forages from the Caribbean”. Animal Feed Science and Technology, 51(1–2): 1–13, ISSN: 0377-8401, DOI: 10.1016/0377-8401(94)00688-6.
Bohm, B. A. & Koupai-Abyazani, M. R. 1994. “Flavonoids and Condensed Tannins from Leaves of Hawaiian Vaccinium reticulatum and V. calycinum (Ericaceae)”. Pacific Science, 48(4): 458–463, ISSN: 0030-8870.
Boufennara, S., López, S., Bousseboua, H., Bodas, R. & Bouazza, L. 2012. “Chemical composition and digestibility of some browse plant species collected from Algerian arid rangelands”. Spanish Journal of Agricultural Research, 10(1): 88–98, ISSN: 2171-9292, DOI: 10.5424/sjar/2012101-134-11.
Díaz, T. 2014. “Contribution of livestock production to food and nutritional security and to the reduction of poverty in Latin America and the Caribbean”. Cuban Journal of Agricultural Science, 48(1): 3–4, ISSN: 2079-3480.
Friedrich, T. 2014. “Production of animal origin feed. Current events and perspectives”. Cuban Journal of Agricultural Science, 48(1): 5–6, ISSN: 2079-3480.
Galindo, W., Rosales, M., Murgueitio, E. & Larrahondo, J. 1989. “Sustancias antinutricionales en las hojas de árboles forrajeros”. Livestock Research for Rural Development, 1(1): 36–47, ISSN: 0121-3784.
Hernández, J. A., Pérez, J. M., Bosch, D., Rivero, L., Camacho, E., Ruíz, J., Salgado, E. J., Marsán, R., Obregón, A., Torres, J. M., Gonzáles, J. E., Orellana, R., Paneque, J., Ruiz, J. M., Mesa, A., Fuentes, E., Durán, J. L., Pena, J., Cid, G., Ponce de León, D., Hernández, M., Frómeta, E., Fernández, L., Garcés, N., Morales, M., Suárez, E. & Martínez, E. 1999. Nueva versión de clasificación genética de los suelos de Cuba. La Habana, Cuba: AGROINFOR, 64 p., ISBN: 959-246-022-1.
Jie-Ping, F. & Chao-Hong, H. 2006. “Simultaneous quantification of three major bioactive triterpene acids in the leaves of Diospyros kaki by high-performance liquid chromatography method”. Journal of Pharmaceutical and Biomedical Analysis, 41(3): 950–956, ISSN: 0731-7085, DOI: 10.1016/j.jpba.2006.01.044.
Makkar, H. P. S. 2003. “Effects and fate of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effects of feeding tannin-rich feeds”. Small Ruminant Research, 49(3): 241–256, ISSN: 0921-4488, DOI: 10.1016/S0921-4488(03)00142-1.
Makkar, H. P. S., Dawra, R. K. & Singh, B. 1988. “Determination of both tannin and protein in a tannin-protein complex”. Journal of Agricultural and Food Chemistry, 36(3): 523–525, ISSN: 0021-8561.
Makkar, H. P. S., Sánchez, M. & Speedy, A. W. 2007. Feed supplementation blocks. (ser. FAO animal production and health paper, no. ser. 164), Rome, Italy: Food and Agriculture Organization of the United Nations, 248 p., ISBN: 978-92-5-105438-3.
Mehrez, A. Z. & Ørskov, E. R. 1977. “A study of artificial fibre bag technique for determining the dig estibility of feeds in the rumen”. The Journal of Agricultural Science, 88(3): 645–650, ISSN: 1469-5146, 0021-8596, DOI: 10.1017/S0021859600037321.
Murray, R. K. & Keely, F. W. 2009. “La matriz extracelular”. In: Murray, R. K., Bender, D. A., Botham, K. M., Kennelly, P. J., Rodwell, V. W. & Weil, P. A., Bioquímica ilustrada, McGraw-Hill Interamericana, p. 527, Available: <http://blogs.sld.cu/marionod/2010/08/12/robert-k-murray-bioquimica-ilustrada-de-harper-28e-mcgraw-hill-interamericana-editores-sa-de-cv-2009-capitulo-45-radicales-libres-y-nutrientes-antioxidantes/>, [Consulted: April 2, 2017].
Muzquiz, M., Cuadrado, C., Ayet, G., de la Cuadra, C., Burbano, C. & Osagie, A. 1994. “Variation of alkaloid components of lupin seeds in 49 genotypes of Lupinus albus L. from different countries and locations”. Journal of Agricultural and Food Chemistry, 42(7): 1447–1450, ISSN: 0021-8561.
Newell-McGloughlin, M. 2008. “Nutritionally Improved Agricultural Crops”. Plant Physiology, 147(3): 939–953, ISSN: 1532-2548, DOI: 10.1104/pp.108.121947.
Obadoni, B. O. & Ochuko, P. O. 2002. “Phytochemical Studies And Comparative Efficacy Of The Crude Extracts Of Some Haemostatic Plants In Edo And Delta States Of Nigeria”. Global Journal of Pure and Applied Sciences, 8(2): 203–208, ISSN: 1118-0579, DOI: 10.4314/gjpas.v8i2.16033.
Pedraza, O. R. M. 2008. “Metabolitos secundarios no fenólicos en el follaje de árboles y arbustos. Efecto en la fisiología digestiva de rumiantes”. Revista de Producción Animal, 20: 97–101, ISSN: 0258-6010, 2224-7920.
Pérez, A., Ibrahim, M., Villanueva, C., Skarpe, C. & Guerin, H. 2013. “Rasgos funcionales que determinan la calidad nutricional y preferencia de leñosas forrajeras en sistemas de alimentación ganadera en zonas seca”. In: VII Congreso Latinoamericano de Sistemas Agroflorestais para a Produção Pecuária Sustentável, Brasil, Available: <http://biblioteca.catie.ac.cr:5151/repositoriomap/bitstream/123456789/290/1/Pere z_et_al_Rasgos.pdf>, [Consulted: April 1, 2015].
Porter, L. J., Hrstich, L. N. & Chan, B. G. 1985. “The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin”. Phytochemistry, 25(1): 223–230, ISSN: 0031-9422, DOI: 10.1016/S0031-9422(00)94533-3.
Ramírez, J. L. 2010. Rendimiento y valor nutritivo de cinco gramíneas en el Valle del Cauto. Ph.D. Thesis, Universidad de Granma, Granma, Cuba.
Ruiz, T. E., Febles, G. J., Castillo, E., Jordán, H., Galindo, J. L., Chongo, B., Delgado, D., Mejias, R. A. & Crespo, G. J. 2008. Tecnología de producción animal mediante Leucaena leucocephala asociada con pastos en el 100% del área de la unidad ganadera. La Habana, Cuba: Instituto de Ciencia Animal, 6 p., Available: <http://www.produccion-animal.com.ar/produccion_y_manejo_pasturas/pasturas_cultivadas_megatermicas/112-leucaena.pdf>, [Consulted: April 2, 2017].
Ryan, G. D., Rasmussen, S. & Newman, J. A. 2010. “Global Atmospheric Change and Trophic Interactions: Are There Any General Responses?”. In: Baluška, F. & Ninkovic, V. (eds.), Plant Communication from an Ecological Perspective, (ser. Signaling and Communication in Plants), Springer Berlin Heidelberg, pp. 179–214, ISBN: 978-3-642-12161-6, DOI: 10.1007/978-3-642-12162-3_11, Available: <http://link.springer.com/chapter/10.1007/978-3-642-12162-3_11>, [Consulted: April 2, 2017].
Semmar, N., Nouira, S. & Farman, M. 2011. “Variability and ecological significances of secondary metabolites in terrestrial biosystems”. Environmental Research Journal, 5(2–3): 213–302, ISSN: 1994-5396.
Verdecia, D. M. 2014. Composición química y metabolitos secundarios en seis variedades de árboles, arbustos y leguminosas volubles en el Valle del Cauto. Ph.D. Thesis, Universidad de Granma, Granma, Cuba.
Visauta, V. B. 2007. Análisis estadístico con SPSS 14: estadística básica. Madrid, España: McGraw-Hilll Interamericana, 281 p., ISBN: 978-84-481-5670-1.
Wencomo, H. B. & Ortiz, R. 2012. “Comportamiento de la disponibilidad de biomasa y la composición química en 23 accesiones de Leucaena spp.”. Pastos y Forrajes, 35(1): 43–56, ISSN: 0864-0394.
Recibido: 3/1/2017
Aceptado: 3/3/2017
R. S. Herrera. Instituto de Ciencia Animal, Apartado Postal 24, San José de las Lajas, Mayabeque, Cuba. Email: rherrera@ica.co.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}