










My SciELO
 Custom services
Custom servicesServices on Demand
Journal

Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCuban Journal of Agricultural Science
Print version ISSN 0864-0408On-line version ISSN 2079-3480
Cuban J. Agric. Sci. vol.52 no.3 Mayabeque July.-Sept. 2018 Epub Sep 01, 2018
Artículo de Revisión
Comportamiento de la producción lechera, reproducción y longevidad en el Holstein y sus cruces con Cebú
1Instituto de Ciencia Animal, Apartado Postal 24, San José de Las Lajas, Mayabeque, Cuba.
Se utilizaron los registros de vacas lecheras Mambí de Cuba (3/4 Holstein 1/4 Cebú), Siboney de Cuba (5/8 Holstein 3/8 Cebú) y Holstein, de tres ganaderías, para determinar el efecto del grupo genético (GG) y diversos factores no genéticos, para rasgos de producción lechera (131 658 lactancias de 63 066 vacas), reproducción (117 794 eventos reproductivos de 36 331 vacas), longevidad (18 741 vacas) y producción de leche por vida (56 536 vacas). Se estudiaron los rasgos producción de leche acumulada hasta 305 días (L305), duración de la lactancia (DL), edad al primer parto (Epar1), intervalo parto primer servicio (IPS), intervalo parto gestación (IPG), intervalo entre partos (IPP), vida productiva (VProd), número de partos por vida (NP) y producción de leche por vida (LTVida). Se aplicó un modelo lineal mixto, con el uso del procedimiento MIXED del SAS. Este modelo incluyó los efectos fijos del genético, rebaño, número de lactancia, año, época de parto, interacciones grupo genético x número de lactancia, grupo genético x año de parto y grupo genético x época de parto. Como aleatorios se incluyeron la vaca anidada en el rebaño y el error. Para los rasgos Epar1, VProd, NP y LTVida, el modelo no incluyó el número de lactancia ni su interacción. Los efectos fijos fueron significativos, a excepción de la época de parto y su interacción con el grupo genético en los rasgos reproductivos y de longevidad. Se concluye que el Mambí de Cuba presentó mejores producciones lecheras acumuladas durante la vida y mayor longevidad.
Palabras clave: bovinos lecheros; efectos ambientales; grupos genéticos
La mayoría de las vacas de los rebaños genéticos destinados a la producción lechera en Cuba corresponden a cruces de la raza Holstein con el Cebú. Predomina el uso del cruce 5/8 Holstein 3/8 Cebú, denominado Siboney de Cuba, y el 3/4 Holstein 1/4 Cebú, conocido como Mambí de Cuba. Una menor proporción representa el Holstein puro.
Si bien en los inicios de la formación de las razas Siboney de Cuba y Mambí de Cuba se realizaron experimentos para comparar el comportamiento del cruce 5/8 Holstein 3/8 Cebú, el 3/4 Holstein 1/4 Cebú y el Holstein puro (Ponce de León et al. 1988), una vez conformadas las razas, no se dispone de estudios que evalúen de conjunto los animales de las tres razas en condiciones de producción para comparar las ventajas del uso de una u otra. Por ello, el objetivo de este estudio fue evaluar los factores genéticos y ambientales que influyen en la producción de leche, la reproducción y la longevidad de vacas Siboney de Cuba, Mambí de Cuba y Holstein.
Materiales y Métodos
Se utilizaron los registros de 1984 a 2016 de la producción lechera, la reproducción y la longevidad de tres grupos genéticos: Mambí de Cuba (3/4 Holstein ¼ Cebú), Siboney de Cuba (5/8 Holstein 3/8 Cebú) y Holstein, localizados en el Centro Nacional de Control Pecuario (CENCOP). Dichos registros pertenecen a tres empresas genéticas (Empresa Pecuaria Genética de Matanzas, Camilo Cienfuegos y Los Naranjos) de la región occidental de la República de Cuba.
El sistema de alimentación de las vacas estuvo basado en pastoreo. Las vacas lactantes consumieron, fundamentalmente, los pastos Estrella (Cynodon nlemfuensis), Pangola (Digitaria decumbens), Guinea (Megathyrsus maximus), algunas especies de pasto naturales, y Cenchrus purpureus vc. Cuba CT-115. Este último como reserva estratégica de los períodos de sequía. También se les suministró granos secos deshidratados con solubles (DDGS) a partir del 2001, a razón de 0.46 kg/L, desde el segundo litro producido, aproximadamente.
Durante el período poco lluvioso se suministró suplementación de caña de azúcar (Saccharum officinarum), King Grass (Cenchrus purpureus) en forma de forraje y hollejo de cítrico, según disponibilidad. La urea y las sales minerales formaban parte de los alimentos complementarios, según lo requería la dieta.
Las hembras no lactantes se mantuvieron en las mismas condiciones de pastoreo. A los treinta días preparto recibieron suplementación con concentrado, según disponibilidad. No se sobrepasaron de 2-3 kg/vaca/d. Durante el período de 1991 a 2000 no se ofreció ningún suplemento a las vacas preparto. Se realizó el ordeño mecánico dos veces al día. Los intervalos entre ordeños fueron de 10 y 14 h. Los horarios de pastoreo en las principales empresas estudiadas fueron los denominados normales (mañana, tarde y noche).
Se eliminaron aquellas lactancias con menos de 30 d de duración. Las edades al parto, inferiores a los 24 meses y las lactancias superiores a la décima lactancia, se agruparon en esta última. La época de parto se agrupó en lluviosa (mayo a octubre) y poco lluviosa (noviembre a abril).
Se emplearon diferentes conjuntos de datos. El fichero de datos de la producción lechera presentó un total de 131 658 lactancias, provenientes de 63 066 vacas, con años de parto de 1987 al 2016. De ellas, 50 022 lactancias correspondieron al Mambí de Cuba (16 128 vacas), 51 911 al Siboney de Cuba (20 769 vacas) y 29 725 al Holstein (26 169 vacas). Se estudiaron los rasgos producción de leche acumulada hasta 305 d (L305) y duración de la lactancia (DL). Para el estudio de la edad al primer parto (Epar1), se consideró la información de la primera lactancia para 52 342 vacas en total. De ellas, 21 899 fueron Holstein, 12 637 Mambí de Cuba y 17 806 Siboney de Cuba.
El fichero de datos de la reproducción presentó 117 794 eventos reproductivos, de 36 331 vacas, con años de parto de 1985 a 2014. De ellos, 56 807 eventos correspondieron al Mambí de Cuba (16 934 vacas), 8 861 al Siboney de Cuba (15 654 vacas) y 12 126 eventos al Holstein (3 743 vacas). Se estudiaron los rasgos intervalo parto primer servicio (IPS), intervalo parto gestación (IPG), e intervalo entre partos (IPP).
A partir del fichero de bajas de la producción de leche, se calcularon dos rasgos de longevidad: la vida productiva (VProd), como los meses del primer al último parto, y el número de partos por vida (NP). El fichero presentó un total de 18 741 vacas, con años de parto de 1984 a 2014. De ellas, 8 302 vacas fueron Mambí de Cuba, 8 514 Siboney de Cuba y 1 925 Holstein. A partir de este fichero también se determinó la producción de leche por vida (LTVida). El fichero presentó 56 536 vacas en total, con años de parto de 1984 a 2016. De ellas, 11 210 correspondieron a vacas Mambí de Cuba, 13 863 Siboney de Cuba y 31 463 Holstein.
Se aplicó un modelo lineal mixto, con el uso del procedimiento MIXED del SAS, versión 9.3 (SAS 2013) según el tipo de datos. El modelo I utilizado para los rasgos de la producción lechera (L305, DL) fue:
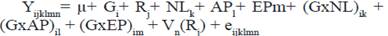
Donde:
Yijklmn |
- vector de las observaciones de las vacas |
µ |
- Media común para todas las observaciones |
Gi |
- Efecto fijo del i-ésimo grupo genético de la vaca (i= 1, 2,3) |
Rj |
- Efecto fijo del j-ésimo rebaño (j= 1, …, 474) |
NLk |
- Efecto fijo del k-ésimo número de lactancia (k=1,.., 10) |
APl |
- Efecto fijo del l-ésimo año de parto (l= 1987,…, 2016) |
EPm |
- Efecto fijo de la m-ésima época de parto (m=1, 2) 1=lluviosa, 2=poco lluviosa |
(GxNL)ik |
- Efecto de la interacción del i-ésimo grupo genético de la vaca con el k-ésimo número de lactancia (ik=1,..,30) |
(GxAP)il |
- Efecto de la interacción del i-ésimo grupo genético de la vaca con el l-ésimo año de parto (il=1,..,87) |
(GxEP)im |
- Efecto de la interacción del i-ésimo grupo genético de la vaca con la m-ésima época de parto (im=1,..,6) |
Vn(Rj) |
- Efecto aleatorio de la n-ésima vaca (n=1, …,63 066) anidada en el j-ésimo rebaño (j= 1, …,474) ~ NID (0, σ2 v) |
eijklmn |
- error aleatorio asociado con cada observación ~ NID (0, σ2 e) |
El modelo II utilizado para el rasgo de la producción lechera LTVida fue:
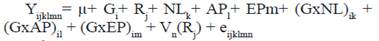
Donde:
Yijklmn |
- vector de las observaciones de las vacas |
µ |
- Media común para todas las observaciones |
Gi |
- Efecto fijo del i-ésimo grupo genético de la vaca (i= 1, 2,3) |
Rj |
- Efecto fijo del j-ésimo rebaño (j= 1, …, 424) |
APl |
- Efecto fijo del l-ésimo año de parto (l= 1984,…, 2016) |
EPm |
- Efecto fijo de la m-ésima época de parto (m=1, 2) 1=lluviosa, 2=poco lluviosa |
(GxAP)il |
- Efecto de la interacción del i-ésimo grupo genético de la vaca con el l-ésimo año de parto(il=1,..,96) |
(GxEP)im |
- Efecto de la interacción del i-ésimo grupo genético de la vaca con la m-ésima época de parto(im=1,..,6) |
Vn(Rj) |
- efecto aleatorio de la n-ésima vaca (n=1, …,56 536) anidada en el j-ésimo rebaño (j= 1, …,424) ~ NID (0, σ2 v) |
eijklmn |
- error aleatorio asociado con cada observación ~ NID (0, σ2 e) |
El modelo III utilizado para los rasgos de la reproducción (IPS, IPG, IPP) fue:
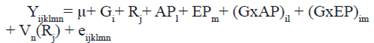
Donde:
Yijklmn |
- vector de las observaciones de las vacas, |
µ |
- Media común para todas las observaciones, |
Gi |
- Efecto fijo del i-ésimo grupo genético de la vaca (i= 1, 2,3) |
Rj |
- Efecto fijo del j-ésimo rebaño (j= 1, …, 473) |
NLk |
- Efecto fijo del k-ésimo número de lactancia (k=1,.., 10) |
APl |
- Efecto fijo del l-ésimo año de parto (l= 1985,…, 2014) |
EPm |
- Efecto fijo de la m-ésima época de parto (m=1, 2) 1=lluviosa, 2=poco lluviosa, |
(GxNL)ik |
- Efecto de la interacción del i-ésimo grupo genético de la vaca con el k-ésimo número de lactancia(ik=1,..,30) |
(GxAP)il |
- Efecto de la interacción del i-ésimo grupo genético de la vaca con el l-ésimo año de parto(il=1,..,90) |
(GxEP)im |
- Efecto de la interacción del i-ésimo grupo genético de la vaca con la m-ésima época de parto(im=1,..,6) |
Vn(Rj) |
- efecto aleatorio de la n-ésima vaca (n=1, …,36 331) anidada en el j-ésimo rebaño (j= 1, …,473) ~ NID (0, σ2v) |
eijklmn |
- error aleatorio asociado con cada observación ~ NID (0, σ2e) El modelo IV utilizado para el rasgo de la reproducción Epar1 fue: |

Donde:
Yijklmn |
- vector de las observaciones de las vacas |
µ |
- Media común para todas las observaciones |
Gi |
- Efecto fijo del i-ésimo grupo genético de la vaca (i= 1, 2,3) |
Rj |
- Efecto fijo del j-ésimo rebaño (j= 1, …, 269) |
APl |
- Efecto fijo del l-ésimo año de primer parto (l= 1984,…, 2014) |
EPm |
- Efecto fijo de la m-ésima época de primer parto (m=1, 2) 1=lluviosa, 2=poco lluviosa. |
(GxAP)il |
- Efecto de la interacción del i-ésimo grupo genético de la vaca con el l-ésimo año de primer parto(il=1,..,90) |
(GxEP)im |
- Efecto de la interacción del i-ésimo grupo genético de la vaca con la m-ésima época de primer parto(im=1, 6) |
Vn(Rj) |
- efecto aleatorio de la n-ésima vaca (n=1, …,52 342) anidada en el j-ésimo rebaño (j= 1, …,269) ~ NID (0, σ2 v) |
eijklmn |
- error aleatorio asociado con cada observación ~ NID (0, σ2 e) |
El modelo V utilizado para los rasgos de longevidad (VProd, NP) fue:
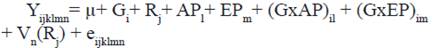
Donde:
Yijklmn |
- vector de las observaciones de las vacas |
µ |
- Media común para todas las observaciones |
Gi |
- Efecto fijo del i-ésimo grupo genético de la vaca (i= 1, 2,3) |
Rj |
- Efecto fijo del j-ésimo rebaño (j= 1, …, 257) |
APl |
- Efecto fijo del l-ésimo año de primer parto (l= 1984,…, 2014) |
EPm |
- Efecto fijo de la m-ésima época de primer parto (m=1, 2) 1=lluviosa, 2=poco lluviosa, |
(GxAP)il |
- Efecto de la interacción del i-ésimo grupo genético de la vaca con el l-ésimo año de primer parto(il=1,..,90) |
(GxEP)im |
- Efecto de la interacción del i-ésimo grupo genético de la vaca con la m-ésima época de primer parto(im=1, 6) |
Vn(Rj) |
- efecto aleatorio de la n-ésima vaca (n=1, …,18 741) anidada en el j-ésimo rebaño (j= 1, …,257) ~ NID (0, σ2 v) |
eijklmn |
- error aleatorio asociado con cada observación ~ NID (0, σ2 e) |
Se aplicó la dócima de Tukey-Kramer para la comparación múltiple de las medias de los mínimos cuadrados, según Kramer (1956).
Resultados y Discusión
El análisis de varianza de los rasgos de la producción de leche mostró que todos los efectos fijos estudiados fueron significativos (tabla 1).
Table 1 Analysis of variance of the fixed effects on milk production traits in Mambí de Cuba, Siboney de Cuba and Holstein cows
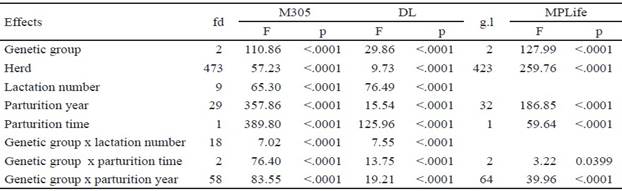
Para los rasgos reproductivos, los efectos de época de parto y de la interacción grupo genético x época de parto no influyeron en el IPS, IPG, IPP y Epar1 (tabla 2). Estos efectos tampoco influyeron en los rasgos de longevidad VProd y NP (tabla 3).
Table 2 Analysis of variance of the fixed effects on reproductive traits in Mambí de Cuba, Siboney de Cuba and Holstein cows
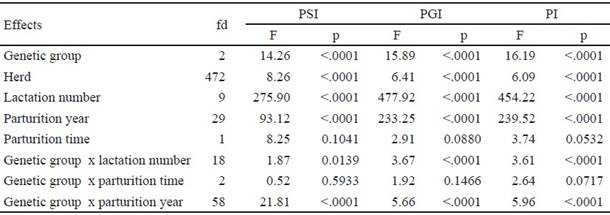
Table 3 Variance analysis of fixed effects on longevity and reproductive traits in Mambí de Cuba, Siboney de Cuba and Holstein cows
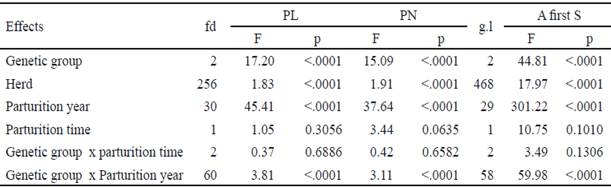
Con relación a los grupos genéticos estudiados, hay que analizar las diferencias entre ellos teniendo en cuenta la época de parto, el número de lactancia y los años de estudio, ya que la interacción genotipo x época de parto, la interacción genotipo x número de lactancia y la interacción genotipo x año de parto afectó los rasgos.
El efecto de la interacción genotipo x época de parto se muestra en la tabla 4. En general, la época de parto afectó los rasgos, de igual manera en las vacas de las tres razas, con mayores promedios en la época poco lluviosa. Al analizar las medias de las tres razas en cada época, hubo mayor promedio de L 305 en el Mambí de Cuba en la época lluviosa. Sin embargo, en la época poco lluviosa, no difirió el comportamiento del Mambí con el del Holstein. La DL fue mayor en el Holstein en ambas épocas; mientras que LTVida fue superior en el Mambí de Cuba en los dos períodos.
La mayor producción de leche a favor de la época poco lluviosa se atribuyó, fundamentalmente, a que en dicha época se presentó un ambiente más fresco. Las temperaturas fueron menores y favorecieron un bienestar metabólico apropiado para la producción lechera, lo que infiere un incremento posterior de la producción de leche, al desarrollar la lactancia en la época de mayor disponibilidad de pastos. Similares resultados se informaron en otros estudios en condiciones tropicales diversas (García 2005 y López et al. 2009).
El efecto de la interacción genotipo x número de lactancia en la producción de leche se muestra en la figura 1. En las tres primeras lactancias, las producciones fueron superiores en el Mambí de Cuba, intermedias en el Holstein e inferiores en el Siboney de Cuba. De la cuarta a la novena lactancia, no hubo diferencias en las producciones lecheras del Mambí de Cuba y el Holstein, las cuales superaron al Siboney de Cuba. En la décima lactancia, no hubo diferencias entre razas.
El intervalo entre partos (figura 2) solamente difirió entre genotipos del cuarto al séptimo parto, con menores promedios en el Siboney de Cuba. En todos los grupos raciales hubo disminución progresiva del intervalo parto-gestación y el intervalo entre partos, con el incremento del número de partos. Este comportamiento se puede atribuir a que las vacas con dificultades reproductivas, generalmente son eliminadas del rebaño. García et al. (2002, 2003) y López et al. (2010) también refieren reducción en el intervalo entre partos, en la medida que aumentó el número de partos.
Table 4 Effect of the interaction genotype x parturition time on milk production and reproduction traits in Mambí de Cuba, Siboney de Cuba and Holstein cows.
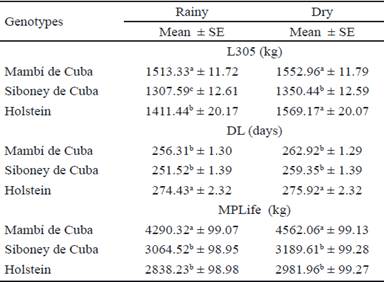
abcParameters with mismatched superscripts in the same column differ to P <0.05 (Kramer 1956).
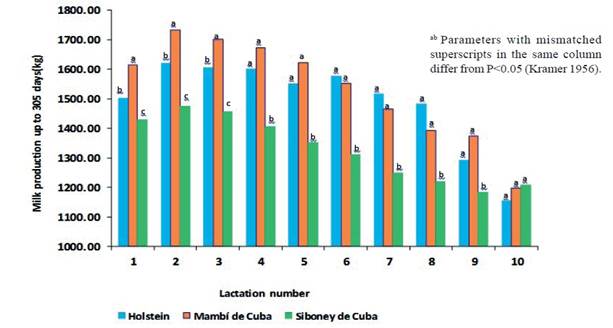
Figure 1 Effect of the interaction genotype x lactation number in milk production up to 305 days in Mambí de Cuba, Siboney de Cuba and Holstein cows
Las mayores producciones lácteas se obtuvieron en el período de 1987 a 1990 (figura 3), con mayores promedios en las vacas Holstein (3328.51, 3512.2, 3135.14 y 2881.15 kg, respectivamente) que superaron al Mambí de Cuba (2584.24 - 2115.55 kg) y al Siboney (1917.87 - 1721.1 kg). De los años 1991 al 2001, el comportamiento fue inestable. En 1991, 1993 y 1999 el Holstein superó al Mambí de Cuba y al Siboney de Cuba. En la mayoría de los años restantes de este período, hubo similares producciones en el Holstein y el Mambí, cuyos promedios superaron a los del Siboney de Cuba. Se observó disminución drástica en las producciones en general, lo que obedece a que este fue un período de limitaciones económicas en Cuba, en el que se redujeron los insumos y no se pudo mantener un suministro sistemático de pienso a las vacas lactantes. Por ello, la alimentación fue fundamentalmente a base de pastos, sin riego ni fertilización.
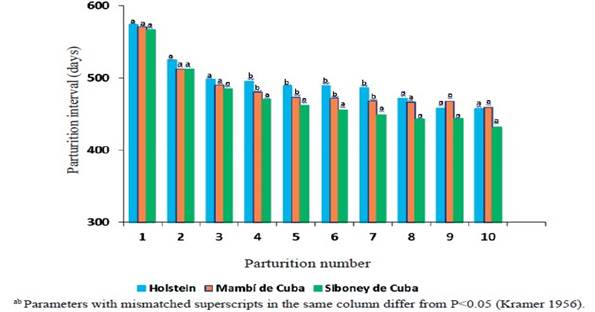
Figure 2 Effect of the interaction genotype x parturition number in the parturition interval in Mambí de Cuba, Siboney de Cuba and Holstein cows.

Figure 3 Effect of the interaction genotype x parturition year in milk production in Mambí de Cuba, Siboney de Cuba and Holstein.
Del 2002 al 2007, hubo ligero incremento de las producciones y se constataron diferencias entre los genotipos. Fueron superiores en el Mambí de Cuba, de los años 2002 al 2004, con promedios de 1850.30 ± 44.30, 1917.56 ± 50.65 y 1761.85 ± 35.41 kg, respectivamente. Mientras, de los años 2005 al 2007, y de 2009 al 2012, el Mambí de Cuba y el Siboney de Cuba presentaron producciones similares.
Los valores del intervalo entre partos (figura 4) de 1985 a 1988 fueron similares en los tres genotipos. De 1989 a 1991 fueron mayores en el Holstein. A partir de 1990, se incrementaron y alcanzaron los mayores promedios en 1991, con 643.49 d en el Holstein, 617.25 d en el Mambí de Cuba y 608.55 d en el Siboney de Cuba, para luego comenzar a disminuir. El incremento notable de los intervalos reproductivos en este período se debió, fundamentalmente, a las limitaciones en la alimentación antes referidas, lo que provocó dificultades para que las vacas quedaran gestantes. De 1992 a 1994, el Holstein y el Mambí de Cuba presentaron mayores IPP con respecto al Siboney de Cuba. Sin embargo, en la etapa posterior, los tres genotipos tuvieron un comportamiento similar, excepto en el 2001.
Los promedios de la vida productiva, según el año de primer parto (figura 5), mostraron variaciones, así de 1985 a 1990 el comportamiento fue similar en los tres genotipos. De 1991 a 1993, los promedios fueron superiores en el Mambí de Cuba y el Holstein. Mientras, de 1994 a 2005 (excepto en 1996) se mostraron iguales promedios de VProd para los tres genotipos. En la etapa del 2006 al 2012, el Holstein presentó menor duración de VProd.

Figure 4 Effect of the interaction genotype x parturition year in parturition interval in Mambí de Cuba, Siboney de Cuba and Holstein.
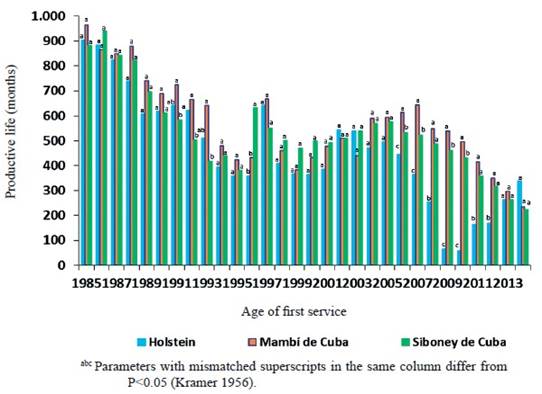
Figure 5 Effect of the interaction genotype x year of first parturition in the productive life in Mambí de Cuba, Siboney de Cuba and Holstein.
En general, los promedios estimados para los rasgos de la producción lechera, reproducción y longevidad en vacas Mambí de Cuba, Siboney de Cuba y Holstein (tabla 5) mostraron mayores producciones de leche acumulada hasta 305 d en el Mambí de Cuba y el Holstein, sin embargo, el Mambí de Cuba presentó mayores producciones acumuladas durante toda la vida superando en 1 299.13 kg al Siboney de Cuba y en 1 516.1 kg al Holstein. El Mambí de Cuba y el Holstein presentaron similar comportamiento reproductivo. No obstante, el Siboney de Cuba mostró mejor comportamiento con menor edad al primer parto y menores intervalos reproductivos. El Mambí de Cuba también tuvo mayor vida productiva, superando en 3.36 meses de duración al Siboney de Cuba y en 11.46 meses al Holstein.
Table 5 Minimum square means (± standard error) for traits of milk production, reproduction and longevity in Mambí de Cuba, Siboney de Cuba and Holstein cows

En las investigaciones realizadas por López et al. (2009), en tres hatos de bovinos cruzados (– Holstein (H) x ¼ Cebú (C), – Suizo (S) x ¼ C, 1/2H x 1/2C y 1/2S x 1/2C) en Veracruz, México, el grupo racial también tuvo efecto significativo en la producción de leche. Las vacas –H x ¼C mostraron mejor producción de leche por lactancia. Los autores argumentan que dicho comportamiento se debe al mejor ambiente, por la inclusión de complementos concentrados y el manejo del ordeño, sin el apoyo de los terneros.
Con respecto al comportamiento reproductivo, López et al. (2006, 2009) refirieron que el incremento en el IPG en animales con mayores proporciones de genes Bos taurus se explica, parcialmente, por un incremento en la producción de leche de dichos genotipos. Los resultados también se corresponden con los de López et al. (2010), en diferentes cruces Bos taurus x Bos indicus en México, en los que observaron que al incrementarse el porcentaje de genes Bos taurus también incrementaba el intervalo entre partos.
Los resultados de este estudio evidenciaron que los grupos genéticos con mayor potencial lechero (Holstein y el Mambí de Cuba) tienen comprometido su comportamiento reproductivo. En correspondencia, varios autores establecen antagonismo entre la producción láctea y la reproducción, especialmente al inicio de la lactancia, etapa en la que existe mayor demanda por los nutrientes (Ferguson 1991, McClure 1995, Pedroso y Bonachea 1995 e Inostroza y Sepúlveda 1999).
Ferguson et al. (1988) observaron descenso de la fertilidad al aumentar la producción lechera. Ellos sugirieron que la selección genética para obtener vacas de mayor producción ha resultado en una selección concomitante de un ganado con menor capacidad reproductiva.
De Luca (2017) explicó que las novillas Holstein tenían índices de concepción cercanos al 70 %. Este autor demostró que esta raza es genéticamente fértil, aunque sus madres de alta producción láctea presenten índices de aproximadamente 40 %. Esto confirma que los problemas de fertilidad no son genéticos, sino fisiológicos y de manejo. Según el autor, al incrementarse la producción de leche pasan a primer término las patologías metabólicas a nivel del útero, los ovarios y las mamas, denominadas “enfermedades de la producción”. Se considera, como causa etiológica de estas enfermedades, un desequilibrio entre los nutrientes ingeridos con los alimentos, y los aprovechados y/o eliminados. Este desequilibrio provoca el “síndrome de parto” de la vaca de alto rendimiento. Se entienden como tal todos los procesos patológicos relacionados con la tensión a que se ve sometido el animal, desde la gestación avanzada hasta el parto, la posterior producción láctea y la nueva concepción.
Los resultados de este trabajo mostraron que el Mambí de Cuba y el Holstein tienen similar comportamiento reproductivo y mayores producciones de leche que el Siboney de Cuba. Sin embargo, este último mostró mejor comportamiento reproductivo. El Mambí de Cuba presentó mayor vida productiva y mayores producciones lecheras acumuladas durante toda la vida.
References
De Luca, L. 2017. Vacas lecheras de alta producción, adaptación metabólica durante el período peripartal. Available: http://www.actualidadganadera.com/articulos/vacas-lecheras-de-alta-produccion-adaptacion- metabolica-durante-el-periodo-peripartal.html. [ Links ]
Ferguson, I. 1991. Nutrition and reproduction in dairy cows. Veterinary Clinics of North America Food Animal. p: 483-507. [ Links ]
Ferguson, J.D., Blanchard, T., Galligan, D.T., Hoshall, D.C. & Chalupa, W. 1988. Infertility in dairy cattle fed a high percentage of protein degradable in the rumen. JAVMA. 192: 659. [ Links ]
García, C. R. 2005. Producción de leche de vacas con diferente porcentaje de genes Bos taurus en el trópico mexicano. MSc. Thesis, Universidad Autónoma Chapingo, México. [ Links ]
García, G. A., Cárdenas, C. A., Monterrosa, V., Valencia, L. & Maldonado, J. G. 2002. Caracterización productiva y reproductiva de las explotaciones ganaderas del bajo cauca y el litoral atlántico antioqueños. I. Haciendas la Leyenda y la Candelaria. Revista Colombiana de Ciencias Pecuarias. 15: 293. ISSN: 0120-0690. [ Links ]
García, G. A., Maldonado, J. G. & García, J. G. 2003. Caracterización productiva y reproductiva de las explotaciones ganaderas del bajo cauca y el litoral atlántico antioqueños. II. Comportamiento de cuatro grupos raciales Bos indicus en un sistema de bosque seco tropical (bs-T). Revista Colombiana de Ciencias Pecuarias . 16: 117. ISSN: 0120-0690. [ Links ]
Inostroza, M.A. & Sepúlveda, N.G. 1999. Actividad reproductiva postparto en vacas lecheras frisonas. Arch. Zootec. 48: 429. [ Links ]
Kramer, C. Y. 1956. Extension of Multiple Range Tests to Group Means with Unequal Numbers of Replications. Biometrics. 12: 307. ISSN: 0006-341X, DOI: 10.2307/3001469. [ Links ]
López, O. R., Díaz, H. M., García, M. J. G., Núñez, R. & Martínez, H. P. A. 2010. Eventos reproductivos de vacas con diferente porcentaje de genes Bos taurus en el trópico mexicano. Revista Mexicana de Ciencias Pecuarias. 1: 325. ISSN: 2007-1124. [ Links ]
López, O. R., Vite, C. C., García, M. J. G. & Martínez, H. P. A. 2009. Reproductive and milk yield performance of crossbred cows with different proportions of Bos taurus genes. Archivos de Zootecnia. 58 (224): 683. ISSN: 0004-0592. [ Links ]
López, R., Thomas, M. G., Hallford, D. M., Keisler, D. H., Silver, G. A., Obeidat, B. S., García, M. D. & Krehbiel, C. R. 2006. Case study: Metabolic hormone profiles and evaluation of associations of metabolic hormones with body fat and reproductive characteristics of Angus, Brangus, and Brahman heifers. The Professional Animal Scientist. 22: 273. ISSN: 1080-7446. [ Links ]
Mc Clure, L. 1995. Infertilidad nutricional y metabólica de la vaca. Editorial Acribia S.A España. 507 p. [ Links ]
Pedroso, R. y S. Bonachea. 1995. Influencia de la condición corporal sobre el comportamiento reproductive del ganado bovino. Rev. Cubana Repr. Anim. 21: 1. [ Links ]
Ponce de León, R., de Bien, R. & Caran, N. 1988. A comparison between Holstein, –. ¼ and 5/8. 3/8 holstein-Zebu in their first two lactations. Cuban Journal of Agricultural Science. 22: 121. ISSN: 2079-3480. [ Links ]
SAS 2013. Statistical Analysis Software SAS/STAT®. version 9.3, Cary, N.C., USA: SAS Institute Inc, Available: http://www.sas.com/en_us/software/analytics/stat.html#. [ Links ]
Recibido: 18 de Diciembre de 2017; Aprobado: 08 de Agosto de 2018
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
