










 Custom services
Custom services
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroduction
Moringa oleifera Lam. is the best known species of the 13 identified in the Moringa genus (Font Quer 1975). It shows nutritional and pharmaceutical benefits, which make it highly promising for its promotion and use. One of the characteristics of this species is its rapid growth during the first year, where it can reach from 3 to 5 m high. For this reason, this plant is very demanding regarding mineral nutrition during the first year and its deficiencies are quickly observed in the foliage coloration (Pita-Hernández and García-Quiñones 2012).
The effect of beneficial rhizospheric bacteria on Moringa oleifera and other tree species not belonging to the Legume family has been studied by several authors (Zayed 2012 and Kannan and Rajendran 2015), but little is known about the effect of rhizobia about Moringa. However, results obtained by inoculating rhizobia in different crops not belonging to the Legume family (Bécquer et al. 2012, 2013, 2015), demonstrate its usefulness for the phenological development and productivity of different crops. This fact is presumably stated due to their potential for the emission of substances that promote plant growth (Machado et al. 2013), as well as the solubility of phosphates (López et al. 2013), mechanisms that are responsible to induce a favorable response in the plant.
Although the use of organic fertilizers is recommended in Moringa sowing (Oquendo 2014), biofertilizers can substantially improve their effect, by the production of phytohormones and by the high enzymatic activity of rhizobacteria, which positively affect germination and phenological development of the plant, as well as its yield (Singh et al. 2011).
For these reasons, the general objective of this study was to evaluate the effect of native isolates and reference strains of rhizobia on the germination and incipient development of Moringa plantlet and select the most efficient with a view to a subsequent evaluation under greenhouse or nursery conditions.
Materials and Methods
Plant variety. Moringa oleifera L., from Supergenius, provided by the Experimental Station of Pastures and Forages Sancti Spíritus.
Bacterial material. An amount of 2 reference strains and 8 native isolates was used, which were obtained from radical nodules belonging to forage legumes from cattle ecosystems of Cuba and previously characterized by Bécquer et al. (2016) (table 1).
Substrate characteristics. A non-sterile substrate was used, containing 40 % of organic matter, 40 % of a soft brown soil (Hernández et al. 2015), and 20 % of river sand. Its chemical characteristics are shown in table 2.

Experimental procedure. All reference strains and native isolates of rhizobia grew in solid yeast-mannitol medium (Vincent 1970). They were re-suspended in a yeast-mannitol liquid medium until reaching a concentration of 107-108 cfu/mL. Prior to inoculation, the inocula were dissolved in a sterile isotonic NaCl solution (0.9 %), in a 1:10 ratio.
Sowing and inoculation. The experiment was carried out in trays of Styrofoam, in glass house, where the plantlet grew under natural light conditions in chamber of 2500 to 3500 lux, with temperatures of ± 27 ºC. The relative humidity was 75/85 % and 2 seeds were planted in each pot containing 120 g of substrate. At the time of planting, 5 mL of inoculum was applied per pot. When germinating, it was thinned to leave 1 plantlet.
Evaluated variables. Percentage of germination (10 d, 15 d and 20 d), according to the formula (Ede et al. 2015):
% G: Number of germinated seeds/number of sown seeds x 100
Germination Speed Index (GSI), according to the formula (Terry et al. 2014):
Inoculation Efficiency Index (IEI, %) (Santillana et al. 2012), according to the formula:
In addition, at 20 days after sowing (DAS), stem length (SL, cm) was measured (with ruler graduated from the substrate level up to the apex of the apical branch), root length (RL, cm), number of leaves (NL), dry weight of the aerial part (DWAP, g) and dry weight of the root (DWR, g).
Experimental design, treatments and statistical analysis. In a completely randomized design, with twelve treatments (table 3) and 14 repetitions, an ANOVA analysis was performed. There was absolute control (AC), and the fertilized treatment (FT) consisted in an application of 23.84 mg of NH4NO3/pot, equivalent to 150 kgN/ha. Differences between means were determined by Fisher LSD test. The values with digit counting were transformed by √x, and the percentage data by arcsin√P. The correlation coefficient among variables was calculated (Sigarroa 1985). The statistical program Stat Graphics Centurion (Stat Point Technologies 2010) was used.

Results and Discussion
When evaluating the germination of seeds, it is observed that at 10 DAS, treatments inoculated with the isolates JK5 (64.3 %), and Ho9 (57.1 %) presented the highest values (P <0.0001) with relation to the rest of the treatments, which include absolute control (28.6 %) and fertilized control (35.7 %) (figure 1). However, at 15 DAS, JK5 shows lower values (85.7%) with respect to Ho5 (92.9 %) and Ho9 (92.9%), which in turn were higher (P <0.0001) to the rest of the inoculated treatments, and they equal the fertilized control (92.9 %).
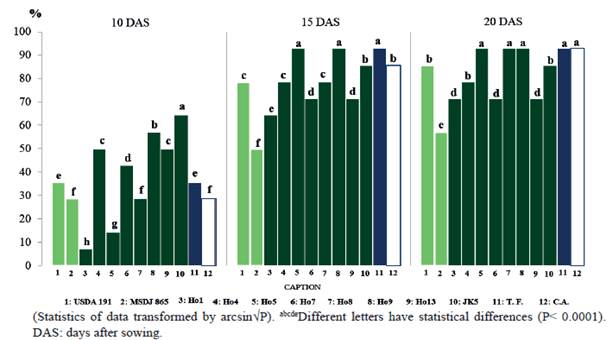
Figure 1 Percentage of germination of seeds of Moringa oleifera, at 10 DAS (SD: 0.0848, P<0.0001); 15 DAS (SD: 0.0699, P<0.0001) and 20 DAS (SD: 0.0661, P<0.0001)
On the other hand, at 20 DAS, JK5 showed lower values (85.7 %) than Ho5 (92.9%), Ho9 (92.9 %) and Ho8 (92.9 %), which were higher (P <0.0001) than the rest of the inoculated treatments, although with similar values to the fertilized control (92.9 %) and absolute control (92.9 %).
None of the two reference strains significantly influenced germination throughout the evaluation period and showed low values at each time of evaluation, especially MSDJ 865.
These results indicate that the greatest impact of inoculation with rhizobia was performed at 15 DAS. Note that the JK5 isolate exerted a greater effect at 10 DAS, however, that treatment was lower than Ho5 and Ho8, as well as the fertilized control (15 and 20 DAS) and absolute control (20 DAS), however, the treatment inoculated with Ho9 maintained superiority in its values from 10 DAS to 20 DAS. The treatment inoculated with Ho8 was superior only at 20 DAS. Apparently, the hormonal substances emitted by different strains of these bacteria have a limited effect on the development of the embryo with respect to the time of application, which does not necessarily prevent its positive effect on other physiological variables of the plant. It is also possible that the effect on some seeds was delayed due to the impermeability of the seminal cortex, which causes a temporary dormancy and affects germination in a certain way (Kassa et al. 2010).
Santillana et al. (2005) observed that 47 % of the evaluated rhizobia strains had a stimulating effect on tomato seeds, which resulted in better germination, possibly due to the ability of rhizobia to produce hormones such as indoleacetic acid, gibberellic acid and cytokinins, substances that regulate plant growth. Cassán et al. (2009) found that a strain of Bradyrhizobium japonicum that stimulated the germination and incipient development of the aerial part of maize, produced abundant indoleacetic acid, zeatin and gibberellic acid, which supports the possibility that these properties of rhizobia have had a positive effect on germination, with better results in treatments inoculated with the previously mentioned strains.
Figure 2 shows that the highest germination speed indexes (GSI) corresponded to the treatments inoculated with the Ho9 isolate (2.32), similar to the fertilized control (2.52), as well as JK5 (2.30). These treatments were superior (P <0.0001) to absolute control and to the rest of the evaluated treatments.
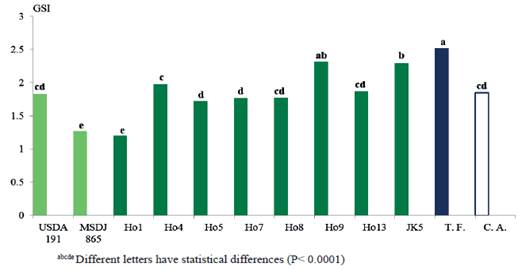
The GSI is an important variable related to vigor of seeds taking into account the number of these that germinate and the time they require to achieve it (Meot-Duros and Magné, 2008). According to Mia et al. (2012), the GSI determines the health state of the planlet and, finally, the productivity of the plant.
Evidently, treatments inoculated with the mentioned isolates had a significant influence on this variable. It is possible that these showed a higher activity of giberellic acid, which can break the latency of seeds and frequently replace the need of environmental stimulus, such as light and temperature (Hernández 2004).
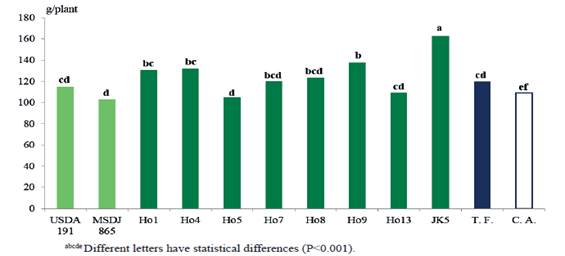
Figure 3 shows the results of dry weight of the aerial part (DWAP), where the superiority of the inoculated treatment with the native JK5 (162.5 g/plant) isolate is evident, regarding the rest of the treatments. It is followed by Ho9 (137.5 g/plant), which was superior (P<0.0001) to absolute control (109.0 g/plant), fertilized control (119.9 g/plant), Ho13 (109.5 g/plant), Ho5 (104.9 g/plant), as well as the reference strain USDA 191 (115.2 g/plant) and MSDJ 865 (103.1 g/plant).
On the other hand, calculating the Inoculation Efficiency Index (IEI) (figure 4), based on the DWAP, it was confirmed that treatments inoculated with isolates JK5 (49.1 %) and Ho9 (26.1 %) showed the highest increases regarding the non-inoculated control. In general, a better efficiency of isolated natives was confirmed in this variable, compared to reference strains.

These results are in contradiction with those found by Mazher et al. (2014), who observed that inoculation with rhizobia had no positive effect on variables of aerial biomass in Moringa oleifera. It is not rejected the possibility that, in this experiment, this variable is related to the increase of values of dry weight of the root due to a strong enzymatic activity of the previously mentioned isolates, and, that way, could favor a higher absorption of nutrients by the plant (McCully 2001).
Figure 5 demonstrates that, in the number of leaves, the treatment inoculated with the isolate Ho9 (29) was superior (P<0.0001) to absolute control (23), to treatments inoculated with MSDJ 865 (21), Ho13 (21), Ho1 (22), Ho8 (22) and USDA 191 (23). Likewise, showed values similar to the fertilized control (28), and to treatments inoculated with JK5 (27), Ho7 (26), and Ho4 (26).

Figure 5 Number of leaves per planlet of Moringa oleifera (Data transformed by √x: S. D.: 0.5702, P<0.001).
In this variable, there is a stimulating effect of one of the isolates (Ho9), which showed superior results in DWAP, and showed it in the amount of leaves.
According to Mia and Shamsudin (2010), auxin synthesis by rhizobia is widely demonstrated, so it is considered that there is a strong activity, in the isolate Ho9, of these enzymes, which are responsible for the apical dominance.
Although there is no abundant literature about the effect of rhizobia on this variable in Moringa, Mazher et al. (2014) found that the application of several commercial biofertilizers based on rhizobia and other plant growth stimulating bacteria had a favorable influence on the number of leaves/plant in Moringa, compared to the non-inoculated control.
Figure 6 shows, in the length of the stem (LT), that the treatment inoculated with the Ho7 native isolate (24.0 cm) had similar results to Ho8 (23.0 cm) and Ho9 (22.5 cm), as well as the fertilized control (22.4 cm) and was superior (P<0.0001) to the rest of the inoculated treatments, and to the absolute control (20.0 cm).
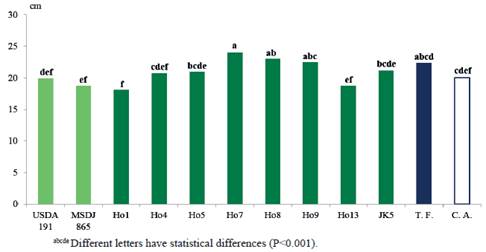
It is inferred that, according to these results, the production of phytohormones by the corresponding isolates, should have an impact not only on the root system, but through it could positively influence the development in the variables of the aerial part, like dry weight and stem length. In addition, Yanni et al. (2001) and Rosenblueth and Martínez-Romero (2006) considered that rhizobia and other microorganisms can penetrate the roots of non-legume species through the cracks or by the points of the lateral appearance of the root and settle in the xylem and in the intercellular spaces of the plants. Therefore, the radius of action of metabolites emitted by bacteria can reach far from the root system. According to reports by Mia et al. (2012), the elongation of the stem in rice seeds, when inoculated with different strains of rhizobia and dinitrofixer bacteria of free life, is due to the production of gibberellin by bacteria of both groups, so it is inferred that Ho7, Ho8 and Ho9, show high activity of this phytohormone
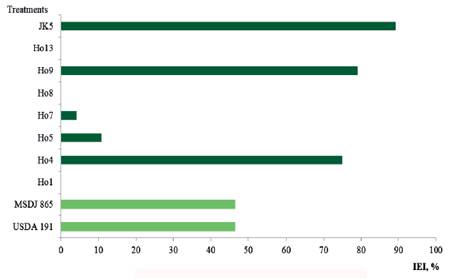
Also in the variable dry weight of the root (DWR) (figure 7), the treatment inoculated with the native isolate JK5 presented the highest value (28.0 g/plant) in comparison with most of the treatments, although its values coincide, in greater measure, with Ho9 (26.5 g/plant) and with Ho4 (25.9 g/plant). It was observed that 50 % of the inoculated treatments showed higher values (P <0.0001) than those of absolute control (14.8 g/plant) and the fertilized control (13.7 g/plant). When calculating the IEI based on this variable (figure 8), the superiority of treatments inoculated with JK5, Ho9 and Ho4 with respect to absolute control was verified, with indexes of 89.2 % (JK5) and 79.1 % (Ho9). The treatment inoculated with Ho4 (75.0 %) also showed higher increases.

Zayed (2012) found a significant increase of length and dry weight of the root of Moringa oleifera, when inoculated with Azotobacter chroococcum and Sacharomyces cerevisae, which, like rhizobia, produce substances that stimulate plant growth. In this regard, Perrine et al. (2004), as well as Kumari et al. (2009), maintain that growth-promoting molecules such as indoleacetic acid, gibberellins and cytokinins produced by the rhizobia present, either in the rhizosphere or in plant tissues, stimulate the highest radical development and increase the absorption ability of root nutrients to its benefit.
In the length of the main root (LMR) (figure 9), the treatment inoculated with Ho7 was superior (P<0.0001) to the rest of the treatments, with values of 9.2 cm, except for the absolute control (8.3cm) and Ho8 (8.1cm).
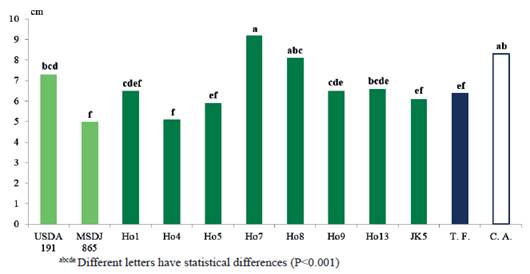
These results do not agree with authors like Mayak et al. (2004), who mention the ability of rhizobia to produce ACC (1-aminocyclopropane, 1-carboxylic acid) deaminase. This compound reduces the level of ethylene in the roots of plants, since ACC is its precursor, with the consequent increase of the length and growth of roots when decreasing this compound. It is inferred that, in spite of the treatments inoculated with Ho7 and Ho8, they excelled with respect to those inoculated with the rest of the strains and native isolates applied, in general, the production of substances that stimulate plant growth by rhizobia, did not favor the longitudinal development of the radicle. According to Lynch (1983), auxins, which are produced by rhizobia and other rhizobacteria, inhibit the elongation of plant roots, although they favor their thickening.
When calculating the correlation between DWAP and DWR, in the inoculated treatments, it was determined that the coefficient was 0.57 (P<0.0001, SE: 24.09). The correlation between DWAP and GSI was also calculated, where the coefficient was similar to the previous one: 0.60 (P <0.05, SE: 15.22). Both coefficients indicate a moderately strong relationship among variables (Sigarroa 1985).
Data suggest a close physiological relationship between the variables analyzed in those treatments where rhizobia strains were applied, where the correlation between the DWAP and GSI draws attention, which indicates a possible dependence on the development of aerial biomass with respect to vigor of seeds. The strong correlation between DWAP and DWR could explain the results obtained in both variables, where the treatments inoculated with Ho9 and JK5 presented superior values, indistinctly of the evaluated variable.
It is concluded that isolates JK5, Ho9 and Ho5 had a superior positive effect on germination at 10 and 15 D A S, as w ell as in mos t of the variables . H ow ever, the reference s trains showed an Inoculation Efficiency Index in DWAP, inferior to that of the native isolates. The 50 % of inoculated treatments showed higher values than absolute control in DWAP and DWR, as well as higher than the control fertilized in the latter variable. On the other hand, nitrogen fertilization did not show a positive effect on any of the evaluated variables. It is noteworthy that there was a moderately strong statistical correlation between DWR and DWAP, as well as between DWAP and GSI, which indicates a high physiological interrelation among these variables.
It is recommended to carry out assays in Green houses or in nursery phase with the isolates Ho4, JK5 and Ho9.

