










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkTecnología Química
versión On-line ISSN 2224-6185
RTQ vol.37 no.1 Santiago de Cuba ene.-abr. 2017
ARTICULO ORIGINAL
Modelo dinámico de un fotobiorreactor de capa fina, utilizado para el cultivo de la microalga Chlorella sp. y bacterias en aguas residuales de alta carga orgánica
Dynamic model of a thin layer photobioreactor, used for the cultivation of the microalga Chlorella sp. and bacteria in wastewater of high organic load
MSc. Orlando Gines Alfaro-VivesI, Ing. Roger Anner Proenza-YeroI, Dr. Rafael Matos-DurandII, Ing. Ajani Rupert-JacobsII
I: Centro de Investigaciones de Energía Solar. Santiago de Cuba, Cuba
II: Universidad de Oriente. Santiago de Cuba, Cuba
Resumen
Se presenta un modelo matemático dinámico para describir el crecimiento simbiótico de la microalga Chlorella sp. y bacterias en un fotobiorreactor de capa fina, utilizado en el tratamiento de aguas residuales de alta carga orgánica. Se demuestra una buena correlación con los resultados experimentales disponibles, ya que las variaciones de los parámetros de proceso (pH, concentración de oxígeno disuelto, concentración de dióxido de carbono disuelto y concentración de substrato en el medio de cultivo) fueron comparados con los resultados experimentales y en el 95 % de los casos coinciden con un error de +-3 %. Además, se evalúa la influencia de los parámetros de operación sobre el rendimiento de algas obtenidas, utilizando el modelo se obtuvo la productividad neta total por unidad de área con un error máximo de +-2,5 % con respecto a los valores experimentales.
Palabras clave: microalgas, tratamiento de aguas residuales, fotobiorreactor.
Abstract
A dynamic mathematical model is presented to describe the symbiotic growth of the microalgae Chlorella sp. and bacteria in a photobioreactor thin film used in the wastewater treatment of high organic load. A good correlation is shown by the experimental results, since the variations of the process parameters (pH, dissolved oxygen concentration, concentration of dissolved carbon dioxide and substrate concentration in the culture medium) were compared with the experimental results and in 95 % of cases coincide with an error of + -3%. Furthermore, the influence of the operating parameters on the performance of algae obtained is evaluated, using the model, the total net productivity per unit area was obtained with a maximum error of + -2, 5 % with respect to the experimental values.
Keywords: microalgae, wastewater treatment, photobioreactor.
INTRODUCCION
El uso de las microalgas en sistemas de tratamiento de aguas residuales, tiene sus orígenes en los trabajos de Oswald y sus colaboradores en la Universidad de California, quienes reconocieron las ventajas de la promoción de la simbiosis entre algas y micro-organismos aeróbicos en aguas residuales domésticas. Las algas producen el oxígeno requerido para la descomposición de la materia orgánica por las bacterias, mientras que las bacterias generan dióxido de carbono que es utilizado como sustrato por las algas.
El trabajo inicial en las lagunas de estabilización (Oswald et. al., 1963); condujo al desarrollo en la década de 1960 de las lagunas de algas de alta velocidad de crecimiento (HRAP), las cuales combinan la función del tratamiento de agua residual con la recuperación de algas (Oswald et al., 1988).
En las décadas de 1980 y 1990 se desarrolló el cultivo en fotobiorreactores de capa fina en Cuba, con la cooperación de la Academia de Ciencia Checa, este sistema permite alcanzar concentraciones de algas-bacterias diez veces superiores a las obtenidas en las HRAP, lo que facilita la separación de los microorganismos y reduce el tiempo de residencia del sustrato orgánico a un día, (Rubio, D., et al., 1992) y (Armas, E., et al., 1993). En la actualidad el interés mundial en el cultivo de la biomasa de las algas como fuente de producción de biofertilizantes y biocombustibles estimula el uso de los fotobiorreactores de capa fina para el crecimiento de algas y bacterias.
El fotobiorreactor de capa fina convencional consiste en dos áreas de 1 750 m2 cada una, con 2 % de pendiente longitudinal separadas entre sí por un pasillo; la diferencia entre ambas áreas radica en que sus pendientes son inversas y forman dos vertientes consecutivas. La superficie del fotobiorreactor será lo más lisa posible y su pendiente transversal será nula; dicha superficie se encuentra dividida a lo largo por muretes que forman canales de un metro de ancho.
El líquido es bombeado desde un tanque guarda, por debajo del nivel inferior de la segunda pendiente, hasta la parte más alta del cultivador, que es el inicio de la primera pendiente donde existe una canal distribuidora formada por tantas compuertas como canales de un metro de ancho tenga el fotobiorreactor, semejante canal distribuidora existirá al inicio de la segunda pendiente, donde llegará la suspensión a través de la primera canal recolectora que se encuentra al finalizar la primera pendiente, de esta forma se recolecta el líquido al terminar su recorrido.
Durante el día se evita la entrada de la suspensión al tanque cerrando la válvula que comunica el tanque con las bombas, de esta forma se establece la recirculación de la suspensión de cultivo por las canales garantizando un flujo de 4 L.s-1 por canal y una profundidad de 0,007 m, con el objetivo de aprovechar la luz solar como fuente energética para el proceso de fotosíntesis. El fotobiorreactor maneja 10 L de suspensión de cultivo por metro cuadrado de superficie de cultivo. Durante la noche se deja entrar la suspensión al tanque guarda donde es aireada para favorecer la respiración de los microorganismos.
Las características de operación en el fotobiorreactor, fueron determinadas por las interacciones detalladas entre varias especies químicas y biológicas presentes en el sistema, así como por las variaciones transitorias de los factores medioambientales. Los procesos de tratamiento de aguas residuales son generalmente sometidos a fluctuaciones de la carga orgánica, temperatura, etcétera, y en el caso de los cultivos de algas realizados a cielo abierto, hay una fuerte influencia ejercida por la irradiación solar incidente. Por tanto, una apreciación de las propiedades dinámicas en el fotobiorreactor de capa fina llega a ser un requisito importante en la operación y diseño exitoso de estos procesos.
El propósito de este trabajo es presentar un modelo matemático que ilustre las características operacionales del proceso en un fotobiorreactor de capa fina utilizado en el cultivo de algas-bacterias, que emplea residual líquido porcino como única fuente de carbono y nutrientes. La intención es validar el modelo, comparando los resultados con los datos experimentales obtenidos en un fotobiorreactor de capa fina de 3500 m2 existente en el cebadero porcino "El Brujo" en Santiago de Cuba.
MATERIALES Y METODOS
Un modelo dinámico de un sistema de crecimiento de algas-bacterias consiste en una serie de ecuaciones que describen las interrelaciones entre varias especies químicas y biológicas presentes. Para este propósito se considera un elemento de fluido pequeño, en el cual, las concentraciones de todas las especies presentes son uniformes. Es necesario examinar la cinética de crecimiento de las algas y bacterias, el equilibrio químico en el sistema y el balance de materias transportadas dentro y fuera del elemento de fluido escogido. Las ecuaciones resultantes se reflejan en la figura 1 y se discute con más detalles abajo.
En la tabla 1 están presentes los valores numéricos de varios parámetros empleados en el modelo, basados en datos de la literatura o de estimaciones como los discutidos por Alfaro O., Juantorena, A., (1997). La influencia de la temperatura en los parámetros de operación no se tiene en cuenta, ya que se utiliza un valor constante.

Fig. 1 Modelo matemático resumido para la descripción del sistema de algas y bacterias.
Tabla 1
Valores numéricos de los parámetros empleados
en el modelo matemático

El crecimiento de las algas
El término microalgas se refiere a un grupo diverso de microorganismos que contienen clorofila, capaces de sintetizar sus propios alimentos a partir del CO2 y el H2O. La microalga Chlorella sp., requiere los siguientes nutrientes para su crecimiento, C, H, O, N, P, K, Mg y otros micros elementos, metálicos. La ecuación estequiométrica (1) que describe su crecimiento fue propuesta por Buhr and Miller (1983):
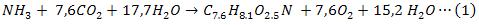
El requerimiento de fósforo es muy pequeño en comparación con el nitrógeno y el carbono, por tanto puede ser despreciado en la estequiometria, (Decostere, B. et al. 2013).
El proceso del crecimiento biológico es descrito normalmente por la función de Monod (1949), ecuación (2), que relaciona la velocidad del crecimiento específico y la concentración limitante de sustrato.

En condiciones donde exista la luz adecuada y las concentraciones necesarias de nitrógeno, el crecimiento de las algas estará limitado por la cantidad de carbono inorgánico disponible. Se conoce que la Chlorella sp., utiliza diferentes especies de carbono inorgánico como sustrato en función del pH, Descotere et al. (2013), Yan (2011) y Banks et al., (2003). Sin embargo, en este estudio se utiliza la concentración del dióxido de carbono disuelto como la especie química que determina la velocidad de crecimiento de las algas ya que demuestra, la disminución en el crecimiento de algas observado a pH alto, resultado del cambio en el equilibrio del ácido carbónico hacia más bicarbonato y carbonato y menor CO2.
Dependiendo de la composición del agua residual alimentada, la disponibilidad de nitrógeno inorgánico en forma de ión amonio o nitrato puede o no ser un factor limitante del crecimiento. Para considerar el nitrógeno como el segundo nutriente limitante, la velocidad de crecimiento será expresada como el producto de dos tipos de funciones de Monod, una para cada uno de los nutrientes limitantes, ecuación (3).
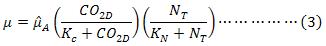
Limitación de Luz
Una restricción más para el crecimiento de las algas que debe ser considerada, es la disponibilidad de la luz. A medida que la irradiación solar penetra la superficie líquida, su intensidad se reduce exponencialmente con la profundidad, de acuerdo a la ley de Lambert -Beer. En un cultivo donde la velocidad de crecimiento no está restringida por la disponibilidad de nutrientes, tres regiones de crecimiento pueden ser distinguidas con la profundidad:
1. La zona superficial donde la intensidad de la luz excede la irradiación de saturación (IS) y la velocidad de crecimiento de las algas es una constante máxima. 
2. Una zona donde la irradiación cae por debajo de IS, y la velocidad de crecimiento disminuye con la profundidad hasta que se convierte efectivamente en cero.
3. Una zona de crecimiento cero donde la luz no penetra.
Como la irradiación incidente en cultivos a cielo abierto es mucho mayor que la IS, la mayor parte del cultivo ocurre en la región 1. Para propósitos de ingeniería se puede plantear que el crecimiento total es directamente proporcional a la penetración de la luz, donde el último se define como la profundidad donde la intensidad alcanza IS (región 1). Al mismo tiempo la naturaleza exponencial de la ley Lambert-Beer afirma que las variaciones sustanciales en irradiación incidente, ocasionarán muy poca diferencia en la profundidad de penetración de la luz y por tanto parece como si la velocidad del crecimiento fuese constante por unidad de superficie del fotobiorreactor. En un estudio sobre limitaciones a los productos fotosintéticos, [Goldman, 1979], también concluyó que el efecto de saturación de la luz resulta en un allanamiento de las curvas de producto, y que los grandes incrementos de la intensidad de la luz conducen solamente a pequeños incrementos de producto. Las grandes concentraciones microbianas dificultan la absorción de luz de las células que están por debajo y a pesar de que el movimiento de las mismas es grande, nunca llega a ser suficiente como para que puedan estar en contacto continuo con los rayos de luz incidente.
Basado en los argumentos anteriores se sugiere que el problema de la limitación de la luz, sea resuelto utilizando la ecuación (4), propuesta por Simmer (1979).

Con KI = 46 W.m-2 y es la constante de saturación de la luz. EZ = 0,7 l.g-1 es el producto del coeficiente de extinción y la profundidad del cultivo y A es la concentración de autótrofos (algas) en (g/L). Io, es la irradiación incidente sobre el cultivo (W.m-2), expresada como una función sinusoidal respecto al tiempo  , donde T es la duración del período de iluminación en (h), t es el tiempo en (h) e Imax es la irradiación máxima en (W.m-2). El crecimiento de las bacterias aeróbicas.
, donde T es la duración del período de iluminación en (h), t es el tiempo en (h) e Imax es la irradiación máxima en (W.m-2). El crecimiento de las bacterias aeróbicas.
La acción de las bacterias heterótrofas en un cultivo de algas-bacterias es similar a la de un proceso de lodos activados, en el cual los microorganismos descomponen la materia orgánica en presencia de oxígeno. Asumiendo una fórmula empírica de C5H7NO2 para protoplasma de bacteria (Buhr and Miller. 1983), y un rendimiento de bacteria de 0.4 g material celular por g de O2 consumido. La estequiometria del proceso aeróbico, está dada por la ecuación (5):
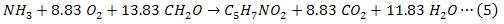
La velocidad de crecimiento de las bacterias, puede ser expresada como el producto de un número de expresiones de tipo Monod, representando limitaciones en sustrato orgánico (S), oxígeno (O2) y nitrógeno (NT).
En este estudio, los fenómenos de nitrificación y desnitrificación, que se espera ocurran en el fotobiorreactor de capa fina, han sido descartados. La inclusión de estos fenómenos en el estudio sería más realista y daría lugar al surgimiento de pequeñas diferencias en las condiciones de operación con respecto a las reportadas aquí, tales como el ligero incremento de la demanda del oxígeno de la nitrificación durante el día, y la reducción en sustrato durante la desnitrificación en la noche., al igual que las variaciones mínimas en pH. Los parámetros usados en el modelo matemático son los que se presentan en la tabla 1.
Solución del equilibrio
Las interrelaciones entre varias especies de carbono inorgánico, amoniaco y otros cationes no reaccionantes y los aniones son complejos, sin embargo son bien analizadas y documentadas por (Buhr and Miller, 1983). Aplicando constantes de equilibrio apropiadas, coeficientes de actividad, y el principio de neutralidad de carga, es posible determinar el pH de la solución y la distribución resultante de sub-especies individuales. Las relaciones requeridas para este propósito están recogidas en la figura 1 y la tabla 1. El valor de la concentración de ión H+ y por tanto el pH está evaluado por prueba y error usando el criterio que las especies carbónicas individuales deben sumar el total de carbono inorgánico presente, el que debe ser igual al obtenido en el balance de materiales.
Balance de masa
Un balance total dinámico de masa para un elemento de fluido puede ser desarrollado teniendo en cuenta que: la acumulación de material en el elemento está dada por la diferencia entre el flujo del material que entra y sale, el que se genera y consume por reacciones y el transferido hacia dentro o hacia fuera por difusión a través de los límites (notablemente la superficie libre). La figura 1 muestra las ecuaciones de balance de masa resultantes para las especies presentes en el sistema, sustrato orgánico, algas, bacterias, carbono inorgánico total, amoniaco total-N y oxígeno.
Simulación del Comportamiento del fotobiorreactor de capa fina
Las ecuaciones en la figura 1 representan las interacciones que ocurrirían en un pequeño elemento de fluido. Fue demostrado que una sección transversal ocurre un mezclado total en el régimen de flujo turbulento (Reynolds > 2100) y que el fotobiorreactor de capa fina, es un sistema de flujo pistón con una dispersión pequeña.
RESULTADOS Y DISCUSION DEL MODELO
Validación de Modelo
Las variaciones diurnas de productividad, remoción de materia orgánica, pH, el oxígeno disuelto y la temperatura en un fotobiorreactor de capa fina para el cultivo de algas bacterias en residuales líquidos porcinos han sido reportadas por Armas, E. y Alfaro O., et al., en 1993, quienes estudiaron un fotobiorreactor de capa fina construido en el cebadero porcino "El Brujo" en Santiago de Cuba. La calidad del modelo matemático puede ser evaluada, comparando sus predicciones teóricas con estos datos de comportamiento. En la tabla 2, se comparan los resultados del modelo con los reportados.
Pnt - es la productividad neta notal en (g/m2.d) y DBO es la DBO5 añadida por metro cuadrado de superficie de cultivo en (g/m2.d).
% R- Porcentaje de remoción de la materia orgánica en (%).
I - Irradiación incidente en (MJ.m-2).
La simulación dinámica de la operación del fotobiorreactor, fue realizada utilizando un tiempo de residencia de un día y un tiempo de recirculación de 10 h. La concentración de materia orgánica añadida se realizó durante las 10 h de operación diurna de forma continua, la temperatura se estimó igual a 33 0C.
La concentración inicial de oxígeno, siempre está en el orden de 0,125 mmol/L y los valores reportados fueron medidos antes de la estación de bombeo y después del punto de alimentación del residual, estos valores fueron estimados en 90 % con respecto a los experimentales.
Los valores predichos por el modelo de oxígeno disuelto y pH coinciden satisfactoriamente con los obtenidos experimentalmente. A medida que el CO2 es consumido por las algas, el pH aumenta debido a consideraciones de equilibrio y el oxígeno disuelto incrementa en las primeras horas y posteriormente se equilibra en un valor constante y más bajo. En contraste, cuando el substrato orgánico es consumido, el nivel de oxígeno disuelto baja y el pH va a disminuir debido a la producción de CO2. Por lo tanto, se anticipa que las dos variables se muevan en la misma dirección, si la tendencia es ascendente, las reacciones de las algas son dominantes, si es descendente, las reacciones bacterias son predominantes.
La simulación demuestra que después de arrancar las bombas, las reacciones de las algas son claramente más rápidas y el pH aumenta, mientras que el CO2 en el sistema es agotado. Cuando el pH alcanza cerca de 9.5, la velocidad de crecimiento de las algas es severamente limitada debido a la no disponibilidad progresiva de CO2. La velocidad con la cual el pH aumenta se reduce y la producción de oxígeno se retarda. El pH, no obstante, se mantiene alto, porque el CO2 disponible es utilizado rápidamente por las algas.
Al medio día, el pH y oxígeno disuelto se mantienen altos, con algunas fluctuaciones. El pH alto causa pérdidas de NH3 a la atmósfera en ubicaciones distantes al punto de alimentación. Ocurren algunas pérdidas de oxígeno a la atmósfera, lo que provoca una disminución lenta de concentración de oxígeno disuelto sin caída del pH.
Las características generales de operación
Los resultados del modelo coinciden con los reportados, sin dudas con el modelo encontrado es posible simular un fotobiorreactor de capa fina para el cultivo de algas-bacterias y definir sus parámetros típicos de operación.
Los datos utilizados se muestran en la tabla 3. La luz fue considerada alternativamente, encendido para ciclos de 10 h de recirculación diurna y apagado para 14 h guardado en el tanque durante la noche.
Tabla 3
Condiciones de entrada empleadas en simulaciones generales

Una representación de las variaciones predichas de oxígeno disuelto, concentración de algas y bacterias, concentración de carbono inorgánico y nitrógeno inorgánico es dada en las figuras 2, 3 y 4 para las 10h de operación diurna.

Fig. 2 El comportamiento de las microalgas (a) y las bacterias (b).
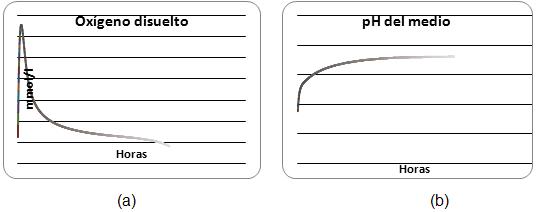
Fig. 3 El comportamiento del oxígeno (a) y del pH (b).

Fig. 4 Comportamiento del nitrógeno (a) y del Carbono (b).
El efecto del inicio de la actividad de las algas al encender las bombas a las 7:00 h es claramente evidente con el aumento rápido del oxígeno disuelto, mientras el substrato empieza a disminuir. El pH comienza aumentando, indicando que la actividad de las algas excede la de las bacterias y hay un agotamiento neto de CO2.
Después de las 7:00 h, el sustrato es consumido progresivamente y el pH aumenta, mientras que la actividad de las algas es dominante. Cuando la concentración del sustrato es alta en el punto de alimentación, es notable que el pH y oxígeno disuelto tiendan a disminuir.
El efecto del flujo del alimentado
No se presenta la influencia del flujo del alimentado con las variables operacionales del sistema, pero el efecto es previsto fácilmente. Esto se demuestra ya que a velocidades de alimentación bajas, el sistema está limitado en carbono; el pH aumenta rápidamente después del arranque del sistema y se queda en un valor alto muy temprano en la mañana. Cuando se emplea mayor flujo de sustrato, el pH aumenta gradualmente después del arranque y puede exhibir una decadencia inicial debido a las velocidades más altas de la generación de CO2. Cuando aumenta la velocidad de alimentación, el pH y el oxígeno disuelto aumentarán progresivamente, hasta que la velocidad de alimentación es tan alta que ocurra una sobrecarga.
El flujo de alimentación es determinado normalmente de acuerdo con el diseño del proceso y será fijo para un sistema particular. Tal decisión se toma teniendo en cuenta la influencia que tiene en el rendimiento obtenido de las algas.
CONCLUSIONES
1. El modelo empleado, describe el comportamiento de un fotobiorreactor de capa fina de 3500 m2, utilizado para el cultivo mixto de la microalga Chlorella sp en residuales líquidos de alta carga orgánica, el error máximo en la productividad neta total diaria por metro cuadrado de superficie de cultivo nunca excede +-2,5 % y las variaciones en las concentraciones de dióxido de carbono y nitrógeno total en la suspensión de cultivo se ajustan a las obtenidas experimentalmente.
2. El valor del pH obtenido en el modelo se ajusta a los datos experimentales y permite la distribución resultante de las sub-especies químicas individuales, lo que facilita utilizar la concentración de CO2 en la suspensión de cultivo como factor limitante del crecimiento de las algas y por tanto regular la velocidad de crecimiento en función del pH. A pH superiores a 10 la velocidad de crecimiento se ve limitada por la ausencia de CO2 en el medio, fenómeno que describe con precisión lo que sucede en la realidad.
3. Cuando no se emplean las cantidades de materia orgánica necesaria para satisfacer la demanda de los microorganismos, el pH crece por encima de 10 y ocurre el fenómeno descrito, es por tanto, indispensable añadir la cantidad de materia orgánica óptima, en función de la concentración de microorganismos presentes y de la irradiación solar incidente.
NOMENCLATURA

Símbolos griegos
| α = constante adimensional en la ecuación (6) |
|
|
γ = coeficiente de actividad | τF= tiempo de residencia hidráulico o del alimentado, h |
| ε = constante de dieléctrica | τR = tiempo de residencia de recirculación, h |
| θ = temperatura, 0C | ε = valencia iónica |
| μ =velocidad de crecimiento biológica específica |
|
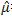 = velocidad de crecimiento especifica máxima
= velocidad de crecimiento especifica máxima
BIBLIOGRAFIA
1. ALFARO, O., JUANTORENA, A., "Tratamiento de residuales líquidos". Revista Tecnología Química. 1997. Vol 17, No.1. Santiago de Cuba.
2. ARMAS, E., ALFARO, O., JUANTORENA, A., "Cultivo de Chlorella sp. con Residual Porcino en un Cultivador de Película Descendente de 3500 m2". Revista Tecnología Química. 1993. Vol 14, No. 2. Universidad de Oriente. Santiago de Cuba, 1992.
3. BANKS, C.J., KOLOSKOV, G.B., LOCK, A.C. and HEAVEN, S., "A computer simulation of the oxygen balance in cold climate winter storage WSP during the critical spring warm-up period". Water science and technology: a journal of the International Association on Water Pollution Research, Vol 48, No. 2, 2003, p.189-196.
4. BUHR, H.O. and MILLER, S.B., "A Dynamic Model of the High-Rate Algal-Bacterial Wastewater Treatment Pond". Water Research, Vol. 17, No. 1, 1983, p. 29-37.
5. DECOSTERE, B., JANSSENS, N., ALVARADO, A., MAERE, T., GOETHALS, P., VAN HULLE, S.W.H. AND NOPENS, I., "A combined respirometer-titrimeter for the determination of microalgae kinetics: Experimental data collection and modelling". Chemical Engineering Journal, Vol. 222, 2013, p.85-93.
6. KOOPMAN, B., BENEMAN, J. y OSWALD, J., "Pond isolation and phase isolation for control of suspended solids concentration in sawege oxidation pond effluents. Algae Biomass". Elsevier. 1980, p. 135 - 161.
7. MONOD, J., Recherches sur la croissance des cultures bacteriennes. Herman & Cie. Paris, 1942.
8. MORENO-GRAU, S., GARCIA-SANCHEZ, A., MORENO-CLAVEL, J., SERRANO-ANIORTE, J. AND MORENO-GRAU, M.D., "A mathematical model for waste water stabilization ponds with macrophytes and microphytes". Ecological Modelling, Vol. 91, Nº(1-3), 1996, p.77-103.
9. OSWALD, W. J. AND GOLUEKE, C. G . "Biological transformations of solar energy". Adv. Appl. Microbiol. Vol. 2, p. 223. 1960.
10. OSWALD, W. J. "The high rate pond in waste disposal". Dev. Ind. Microbiol. Vol 4, p.112-119. 1963.
11. OSWALD, W. J., "Micro-algae and waste water treatment". Microalgal Biotechnology, Borowitzka and Borowitzka (Eds). Cambridge University Press, 1988, p.305-328.
12. RUBIO, D., ARMAS, E., ALFARO, O., JUANTORENA, A., LEAL, A. y ZAYAS, R., Cultivo Mixotrófico de Microalgas Chlorella sp. para la Alimentación Porcina. Informe Final del Resultado. Centro de Investigaciones de Energía Solar. Santiago de Cuba, 1992.
13. SIMMER, J. "Radiation Energy, Temperature and Algal Growth". p. 41-45. 1979.
14. YANG, A., "Modelling and Evaluation of CO2 Supply and Utilization in Algal Ponds". Industrial & Engineering Chemistry Research, Vol. 50, Nº 19. 2011.
Recibido: Septiembre 2016
Aprobado: Diciembre 2016
MSc. Orlando Gines Alfaro-VivesI. Centro de Investigaciones de Energía Solar. Santiago de Cuba, Cuba


{kind=link}