










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Producción Animal
versión On-line ISSN 2224-7920
Rev. prod. anim. vol.29 no.3 Camagüey set.-dic. 2017
MANEJO Y ALIMENTACIÓN
Comportamiento agronómico de cultivares de Cenchrus purpureus tolerantes a la salinidad
Agronomic Behavior of Cenchrus purpureus Cultivars Tolerant to Salinity
José Leonardo Ledea Rodríguez, Diocles Guillermo Benítez Giménez, Ramón Crucito Arias Pérez y Adania Guerra Manso
Instituto de Investigaciones Agropecuarias Jorge Dimitrov, Estación Experimental de Pastos y Forrajes, Bayamo, Granma, Cuba. ledea@dimitrov.cu
RESUMEN
Con el objetivo de determinar el efecto de la edad de corte (60, 90 y 120 días) y del cultivar (CT-500 y CT-115), sobre algunos componentes del rendimiento en nuevas variedades de Cenchrus purpureus, se desarrolló un estudio en la Estación Experimental del Pastos y Forrajes del Instituto de Investigaciones Agropecuarias Jorge Dimitrov. Según diseño de bloques al azar y en arreglo factorial se controlaron los efectos edad de corte (3), el cultivar (2) y su interacción (3 x 2), sobre el rendimiento y algunas variables agronómicas que definen el mismo. Se observó que la interacción no afectó las variables morfológicas de hojas y tallos, pero si la altura de la planta, los cultivares mostraron diferencias significativas (P < 0,001) para las edades de 60 y 90 días, mientras que, para el número de hijos basales y el porcentaje de materia seca en la planta íntegra, el CT-115 fue significativamente superior en las edades de 60 y 120 días, respectivamente. La tasa de crecimiento absoluto y duración de la biomasa se desfasaron significativamente a los 90 días a favor del CT-500. Ambas variedades demostraron el potencial productivo de materia seca por hectárea y solo se diferenciaron (P < 0,001) para la edad de 120 días en condiciones de áreas forrajeras con limitantes de salinidad en el suelo. Se recomienda considerar los resultados del presente estudio como datos preliminares para el diseño de la estructura varietal de áreas forrajeras salinizadas.
Palabras clave: morfología, productividad, Cenchrus purpureus, Pennisetum purpureum.
ABSTRACT
To determine the effect of cutting age (60, 90, 120 days), and cultivar (CT-500 and CT-115) on some yield components in new varieties of Cenchrus purpureus, a study was made at the Experimental Station of Pastures and Forages, at the Jorge Dimitrov Agricultural Research Institute. The effects of cutting age (3), cultivar (2), and their interaction (3 x 2) on yields and some agronomic variables were controlled by a randomized block design. Interaction did not to affect the morphological variables of leaves and stems, though it did affect plant height. The cultivars had significant differences (P < 0.001) for the ages of 60 and 90 days; whereas for the number of basal shoots and percent of dry matter in the whole plant, CT-115 was significantly higher at 60 and 120 days, respectively. The total growth rate and biomass duration significantly phased out at 90 days, toward CT-500. Both varieties showed potential for high dry matter production per hectare, and only differed (P < 0.001) from the 120 days in the forage areas with saline soils. These results must be considered as preliminary data for the design of a variety structure in saline forage areas.
Keywords: Morphology, productivity, Cenchrus purpurus, Pennisetum purpureum.
INTRODUCCIÓN
En Latinoamérica la transformación de los ecosistemas por el efecto del cambio climático, y prácticas agrícolas inadecuadas, propician la degradación de los suelos de los sistemas pastoriles, que reducen la productividad y persistencia de los pastos y forrajes (Pérez-Infante, 2013).
Cuba presenta esta misma problemática, que se asocia al cambio climático y distribución anual de las precipitaciones, que en conjunto con otros factores como la temperatura, la radiación solar, las prolongadas sequías y la creciente salinización de los suelos hace que los rendimientos de los pastos no sean estables durante todo el año (Argentel, Garatuza, Yépez y de los Santos, 2016). Esta situación se acentúa mucho más en el oriente del país, donde las sequías son prolongadas, existiendo elevada evaporación y coeficientes de precipitación/evaporación menores de 0,45, característicos de zonas áridas (Benítez, Cisneros, Pérez-Infante y Nuviola, 2010).
El Valle del Cauto es una llanura dominada por la cuenca del río Cauto en la región oriental cubana. Tiene una extensión de 4.5 miles de km2 y ocupa áreas de cuatro provincias orientales (Holguín, Las Tunas, Santiago de Cuba y Granma), y se desarrolla el 90 % de la ganadería de la región oriental (Fajardo, 2008). El territorio del valle ocupado por la provincia Granma se extiende desde los límites de la Sierra Maestra al sur de la provincia, al Golfo de Guacanayabo y los límites de la llanura costera al oeste, hasta la provincia Santiago de Cuba al este y las Tunas y Holguín al norte (Benítez et al., 2007).
La variabilidad de los cambios atmosféricos y meteorológicos, propician dinámicos cambios en la dirección de los vientos cargados de humedad (ONEI, 2015) provocando que las precipitaciones descarguen mayormente en la costa norte de la región en cuestión, lo que supone una prolongada sequía estacional para el Valle, donde coincide este fenómeno con afectaciones edáficas, entre las que se destacan la erosión (13,5 %), baja fertilidad (11 %), mal drenaje (22,8 %) y pérdida del manto freático (25 %), cuyas afectaciones se solapan (Benítez, Cisneros, Pérez-Infante y Nuviola, 2010), la afectaciones por salinidad alcanzan el 38 % de su área (Álvarez, 2009).
Con el objetivo de estabilizar la producción de forrajes en Cuba bajo estas extremas condiciones edafoclimáticas, se comienza en la década del 2000 el programa de obtención de cultivares de Pennisetum purpureus con tolerancia a la sequía a partir del CT-115 (Herrera et al., 2003). Como resultado se obtuvieron clones con tolerancia a la salinidad. Herrera (2000) al realizar la caracterización molecular de estas plantas mediante la determinación electroforética de más de cinco sistemas isoenzimáticos, demostró que eran variedades y se reafirmó al mantenerse sus características iniciales por más de cinco generaciones.
Posteriormente en 2011, fueron aportadas por el Instituto de Ciencia Animal, al Instituto de Investigaciones Agropecuarias Jorge Dimitrov, con el objetivo de ampliar el germoplasma existente, y fortalecer la base alimentaria en los ecosistemas frágiles y degradados del Valle del Cauto.
El objetivo de este estudio, consistió en evaluar el comportamiento agronómico de las variedades CT-500 y CT-115 bajo la influencia de diferentes las edades de corte en la estación lluviosa, en las condiciones edafoclimáticas del Valle del Cauto.
MATERIALES Y MÉTODOS
Localización y comportamiento de los factores climáticos del área experimental
El estudio abarcó la estación lluviosa (mayo-octubre) del año 2014. Los experimentos se desarrollaron en la Estación Experimental de Pastos y Forrajes perteneciente al Instituto de Investigaciones Agropecuarias Jorge Dimitrov en la provincia Granma, Cuba. Está situada en los 20º 18´13" de latitud norte y los 76º 39´ 48" de longitud oeste.
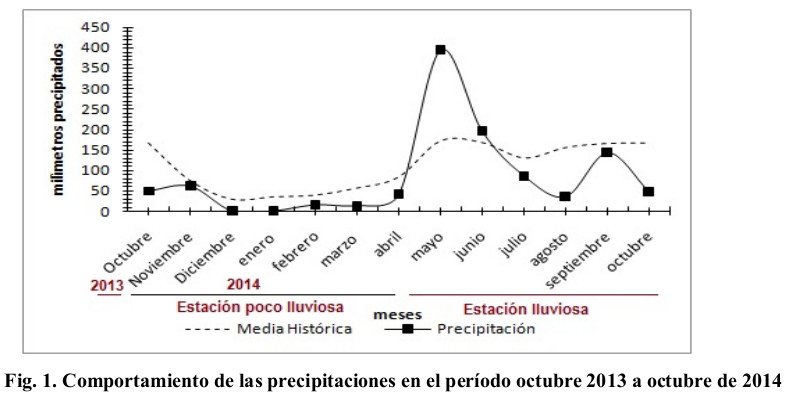
El clima de la región, donde está ubicada la estación, se clasifica como tropical relativamente húmedo (Barranco y Díaz, 1989). En la región donde se encuentra la estación, se alcanzan temperaturas máximas, medias y mínimas de 35,4; 27,1 y 20,3 ºC en la estación lluviosa, y en la poco lluviosa 34,9; 24,8 y 14,8 ºC, respectivamente, con humedades relativas para ambos períodos climáticos que oscilan entre 77,8 y 72,7 % en orden, y altos valores de evaporación con 216,2 mm en lluvias y 191,9 mm en el período poco lluvioso (Ledea, 2016). El comportamiento de las precipitaciones durante la investigación se muestra en la Fig. 1.
El suelo presente en el área experimental es de tipo Fluvisol diferenciado (Hernández et al., 2015). Peña, Álvarez y Díaz (2002) al realizar estudios de perfiles de suelos en el Valle del Cauto, encontraron que 33,3 % de las afectaciones del área eran debido a la salinidad, y en esta región donde está ubicada la estación fue de 100 %, con valores de salinidad de débil a medianamente salinos. La profundidad efectiva tiene rangos de variación desde profundos (91-150 cm) hasta poco profundos (25-50 cm). Presenta un drenaje deficiente, la topografía es llana y generalmente su fertilidad natural se encuentra entre valores medios y bajos con un pH ácido Sus características químicas se señalan en la Tabla 1.
Características del material vegetal utilizado
El material vegetal para el establecimiento de los bancos de semilla fue proporcionado por el departamento de Pastos y Forrajes del Instituto de Ciencia Animal, de las cuales se utilizaron para el estudio el CT-500 obtenidas por cultivos de tejidos a partir del CT-115, este último fue utilizado como control, ambos cultivares se establecieron en parcelas de 10 x 20 m ![]() con 0,75 m entre planta y un metro entre surcos.
con 0,75 m entre planta y un metro entre surcos.
Tratamiento, diseño y análisis estadístico
En arreglo factorial según diseño de bloques al azar con cuatro repeticiones se desarrolló un estudio para evaluar el comportamiento de algunas variables agronómicas y el rendimiento de materia seca por hectárea. Se realizaron análisis de varianza usando modelos lineales en los que se controlaron los efectos de edad de rebrote (3), variedad (2), y su interacción (3 x 2) con un total de 120 observaciones. Para la distribución normal de los datos se empleó la prueba de Kolmogorov-Smirnov (Massey, 1951) y para la homogeneidad de varianza la prueba de Bartlett. Las medias se compararon mediante la dócima de Newman-Keuls (Keuls, 1952). Se utilizó el paquete StatSoft 10.0. El modelo matemático empleado en cada uno de los análisis fue el siguiente:

(EC x V)jk: efecto combinado de la j-ésima edad de corte en la k-ésima variedad
eijk: error aleatorio ![]()
Procedimiento experimental
En los bancos de semillas se utilizaron los surcos de forma intercalada coincidentes con cada edad de corte (60, 90 y 120 días) evaluada, dentro de estos surcos se formaron subparcelas de 4,50 m lineales, cada subparcela contó con cuatro repeticiones y se seleccionaron cinco plantas por cada edad de corte de cada variedad. Luego de realizar las mediciones se ejecutó el corte para estimar rendimiento de materia verde, para ello se eliminó el efecto de borde consistente en el surco precedente y los 50 cm al principio y final del surco en evaluación. Se efectuó el corte de uniformidad en marzo de 2014 para lograr homogeneidad en el material experimental y acto seguido se realizó un pase de arado entre surcos y se aplicó materia orgánica a razón de 20 t.ha-1.
Mediciones de la planta
En el momento del corte se seleccionaron cinco plantas por réplica a las que les fueron retiradas las distintas fracciones botánicas (hojas, tallos y material muerto) con tijera de acero inoxidable y, posteriormente se procedió según metodología propuesta por Herrera et al. (2003) para medir, altura de la planta determinada desde la base hasta el cono apical con la ayuda de una regla milimetrada, número de hojas verdes, porcentaje de ramas y número de hijos basales que surgen a partir de la macolla.
Se seleccionó la cuarta hoja para medir el largo y el ancho según la metodología propuesta por Herrera (2006). El ancho de la hoja se obtuvo a partir del promedio de las mediciones realizadas con una regla milimetrada a los segmentos base, medio y apical, mientras que el para el largo, se contempló desde la base hasta el ápice, medidos también con una regla milimetrada, para el grosor del tallo, medido en el cuarto entre nudo, también se siguieron las especificaciones de este autor medido con pie de rey marca ![]() con precisión de 0,05 mm .
con precisión de 0,05 mm .
Se consideró el porcentaje de hojas verdes, porcentaje de material muerto (% MM), también se tuvo en cuenta el número de nudos y entrenudos, porcentaje de tallos verdes, rendimiento de materia seca por hectárea, y contenido de materia seca de hojas, tallos y planta íntegra, para la relación hoja-tallo-material muerto se tomó una muestra de 200 g de masa verde, a lo que se determinó la materia seca en cada componente según la metodología descrita por Herrera (2006).
RESULTADOS Y DISCUSIÓN
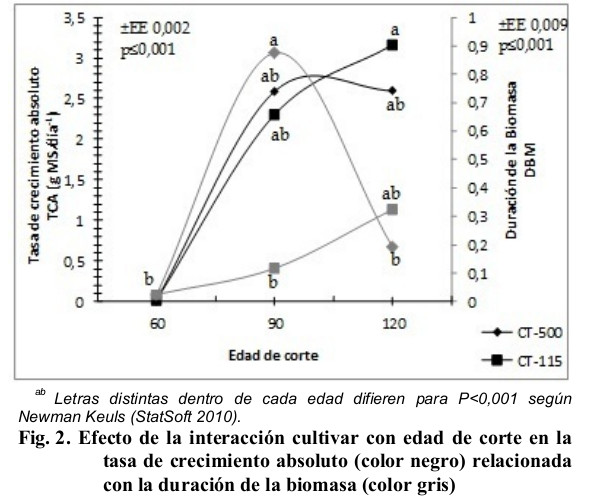
En la Fig. 2 se observa el efecto de la interacción cultivar con edad de corte en la Tasa de Crecimiento del Absoluto (TCA), relacionada con la Duración de la Biomasa (DBM). El mayor valor de TCA se manifestó para el CT-115 a los 120 días al corte, este promedio fue común para las edades de 90 días para ambas variedades y CT-500 a los 120 días, los valores significativamente inferiores se mostraron a la edad de 60 días para ambas variedades y difirieron del resto de los promedios de las edades evaluadas. La DBM fue significativamente superior a los 90 días para el CT-500 con respecto al CT-115, se diferenció significativamente con sus propios promedios en las edades de 60 y 120 días, y con los del CT-115 a los 60 y 90 días.
Este comportamiento demuestra que el CT-500 y CT-115 crecen a tasas absolutas diferentes y por tanto acumulan MS a diferentes ritmos, lo que indica que son las características intrínsecas de cada variedad las que diferenciaron las respuestas fenotípicas observadas en el período lluvioso, según Uvidia, Leonard, Benítez y Buestan (2013), esto se debe a que bajo diferentes estímulos, ambientales en este caso, se activan ciertos genes que estimulan una actividad determinada, que en concomitancia con los factores reguladores y enzimáticos se puede manifestar de forma expedita o no, efecto que pudo haber estado presente en el efecto analizado.
La TCA mostrada por el CT-115 y CT-500 es inferior a los obtenidos por Rincón, Ligarreto y Sanjuanelo (2007) cuando evaluaron de forma individual al maíz y en asociación con pastos naturalizados, y a los obtenidos por Uvidia, Leonard, Benítez y Buestan (2013) en condiciones de ecosistemas amazónicos en Pennisetum purpureum vc Maralfalfa, para la semana cinco (35 días), ocho (56 días) y 12 (84 días), con 4; 5,8 y 4 cm respectivamente, según estos autores en la semana de mayor crecimiento relativo, la planta obtuvo el mayor rendimiento de MS, y a partir de la semana 12 apreciaron acumulación de tallos y MM, sin embargo, en la Fig. 2 se observa que este fenómeno se prolonga hasta la edad de 120 días, aunque se deprime para el CT-500 pero sin mostrar diferencias significativas con la edad precedente ni con la mostrada por el CT-115 de igual forma para los 120 días.
Para la altura de la planta (Fig. 3), para ambas variedades se obtuvieron valores significativamente superiores a la edad de 120 días, los valores para el resto de las edades se mostraron estadísticamente semejantes para las variedades, pero difirieron de los promedios mostrados en cada edad de forma independiente para el CT-115 y CT-500. El promedio alcanzado a los 120 días superó el referido por Cordoví, Ray, Nhatumbo y Chimbalambala (2013) y Fernández, Viamonte, Fonseca y Ramírez (2015) en variedades de C. purpureus en ecosistemas degradados y con intensa sequía estacional, pero fueron inferiores a las obtenidas por Ramírez (2010a) en variedades tolerantes a la sequía en condiciones de pre-montaña, donde los cultivares se vieron favorecidos por el clima imperante en la pre-montaña.
Las precipitaciones poseen marcada influencia sobre el crecimiento de la planta, y se refleja ésta en la altura de la misma. En la Fig. 3, se observa, que en el período climático en que se desarrolló el estudio, las precipitaciones se comportaron por encima de la media histórica mensual (Fig. 1), esto proporciona un incremento en la solubilidad de las sales presentes en el suelo, lo que pudo influir, independientemente del bajo nivel de especialización que presenta el sistema radical de las gramíneas (Rosenzweig et al., 2014), a que no se afectara la altura de la planta y sí que se viera favorecida por el efecto de las precipitaciones, alcanzando valores desde los 70 cm y superiores a los 200 cm . Según Argentel, Garatuza, Yépez y de los Santos (2016) esto está relacionado directamente con el desarrollo radical, y sobre todo la longitud de la raíz, que ha sido reportada por Nevo y Chen (2010) como un importante indicador para la evaluación de la tolerancia a la salinidad en numerosas especies vegetales.
Argentel, Garatuza, Yépez y de los Santos (2016) cuando evaluaron diferentes variedades de trigo en ecosistemas salinos de la región oriental de Cuba, obtuvieron que la altura lograda por las plantas, estuvo estrechamente relacionada con el desarrollo radical logrado por las mismas, sin embargo, este fenómeno descrito no coincide con lo expuesto por Hernández et al. (2003), quienes refirieron que en la estación lluviosa el desarrollo radical se reduce por la translocación de nutrientes a la parte aérea de la planta, al parecer las plantas tolerantes a la salinidad modifican el desarrollo de sus raíces, como mecanismo compensatorio o adaptativo ante la salinidad, por lo que intuimos que el CT-115 y CT-500 tuvieron un favorable desarrollo radical que les permitió una exploración más eficiente del medio donde se desarrollaron, y con ello, alcanzar las alturas reflejadas en la Fig. 3.
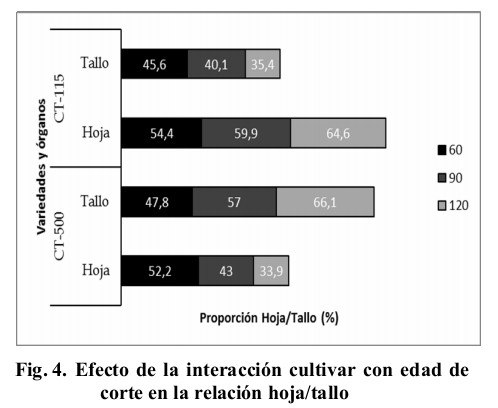
La altura de la planta repercutió en su composición estructural (Fig. 4), donde se pudo apreciar que en la medida que se incrementó la edad de corte se invirtió de forma negativa la proporción de hojas con respecto a los tallos; sin embargo, el CT-500 para los 60 días al corte mostró casi una proporción 50/50 %, para el resto de las edades se desplazó la proporción hacia los tallos, efecto que no manifestó el CT-115, donde para todas las edades estudiadas, presentó mayoritariamente hojas en vez de tallos.
El comportamiento del CT-500 es semejante, y los valores inferiores a los obtenidos por Reyes et al. (2015) en caña de azúcar y en CT-169 respectivamente en condiciones de pastoreo en ambas estaciones climáticas, mientras que el CT-115 asemejó el comportamiento obtenido por Roncallo, Sierra y Castro (2012) en genotipos de Cenchrus purpureus, y superó al observado en caña de azúcar por Reyes et al. (2015). Estas modificaciones estructurales están relacionadas con el hábito de crecimiento de las gramíneas, así como la incidencia de los factores climáticos y capacidad de respuesta de la planta ante los factores mencionados anteriormente (Calzada et al., 2014).
Estos resultados coinciden con los reportados por Herrera (2006), Álvarez (2009) y Ramírez (2010b) en cuanto a la alta proporción de hojas que presenta el CT-115, debido sobre todo al acortamiento de los entrenudos y al número de nudos que presenta (10 aproximadamente), lo que le posibilita brindar mayor cantidad de nutrientes contenidos en las hojas a los animales, representando el contenido de proteína el 11 % de su contenido en base seca (Febles y Herrera, 2006), en este sentido Álvarez (2009) refirió que entre los componentes del pasto las hojas constituyen la base fundamental por excelencia, ya que en ellas el animal encuentra los elementos nutritivos necesarios para su desarrollo, al estar acumuladas grandes cantidades de componentes solubles, por las características tisulares que comprenden células de pared fina y llenas de citoplasma (Pérez Infante, 2013).
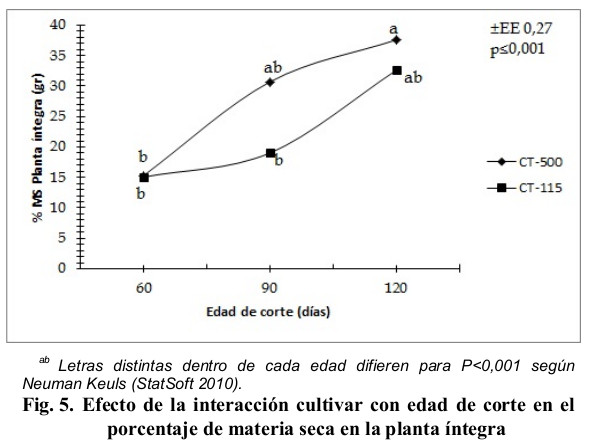
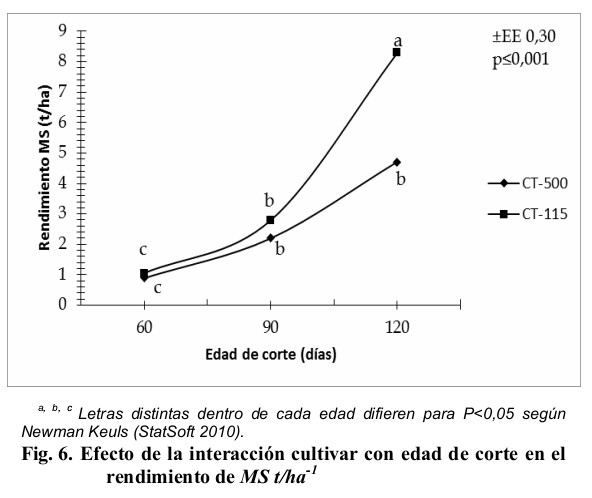
En consecuencia, al comportamiento estructural, se obtuvieron diferencias significativas en el porcentaje de materia seca de la planta íntegra (Fig. 5). El CT-500 fue significativamente superior a la edad de 120 días en la acumulación de materia seca en la planta íntegra, y este promedio difirió (P < 0,001) con sus propios promedios a las edades de 60 días y con los del CT-115 en esta misma edad y para los 90 días, los valores para las edades de 90 días y 120 días de las variedades CT-500 y CT-115, no fueron significativos ![]() con respecto al mayor valor. Este valor superó al referido por Cordoví, Ray, Nhatumbo y Chimbalambala (2013) en tres variedades de C. purpureus en clima semiárido del sur de Mozambique, mientras que los rendimientos de MS del CT-115 y CT-500 (Fig. 6) fueron superiores a los obtenidos por Fernández, Viamonte, Fonseca y Ramírez (2015) cuando evaluaron dos variedades de C. purpureus tolerantes a la sequía en ambas estaciones climáticas, también superaron los resultados de Valles de la Mora, Castillo y Bernal (2016) cuando evaluaron diez gramíneas tropicales en ambas estaciones climáticas en Tlapacoyan, Veracruz, México.
con respecto al mayor valor. Este valor superó al referido por Cordoví, Ray, Nhatumbo y Chimbalambala (2013) en tres variedades de C. purpureus en clima semiárido del sur de Mozambique, mientras que los rendimientos de MS del CT-115 y CT-500 (Fig. 6) fueron superiores a los obtenidos por Fernández, Viamonte, Fonseca y Ramírez (2015) cuando evaluaron dos variedades de C. purpureus tolerantes a la sequía en ambas estaciones climáticas, también superaron los resultados de Valles de la Mora, Castillo y Bernal (2016) cuando evaluaron diez gramíneas tropicales en ambas estaciones climáticas en Tlapacoyan, Veracruz, México.
Independientemente de este comportamiento, es de destacar que el CT-115 solo superó significativamente al CT-500 en rendimiento de MS.ha-1 a la edad de 150 días, y este valor difirió con el resto de los promedios de las edades evaluadas tanto los propios (CT-115) como los pertenecientes al CT-500. Para el resto de las edades no se apreciaron diferencias significativas (P > 0,05) en la acumulación de MS.ha-1.
A pesar de las diferencias comentadas, se destaca que el rendimiento de materia seca no fue afectado por los niveles de salinidad del suelo, el cual presentó conductividades eléctricas en extractos de saturación de 0,047; 0,10 y 0,05 dS.m-1 en 0-20, 20-40 y 40-60 cm de profundidad respectivamente (Ledea, 2016). En este sentido, Schroeder et al. (2013), expusieron que cuando la conductividad en extractos de saturación supera los 3,6 dS.m-1 es cuando se manifiesta en los cultivos una disminución de los rendimientos, debido a dificultades en los procesos que determinan el ajuste osmótico. En consecuencia, Argentel, Garatuza, Yépez y de los Santos (2016) declararon que la acumulación de materia seca en suelos salinos depende en buena medida de las relaciones hídricas de la planta y la capacidad de regulación de esta variable, mediante la acumulación activa de osmolitos e iones inorgánicos, efecto que pudo haberse facilitado en este estudio por el régimen pluviométrico influido por la estación climática.
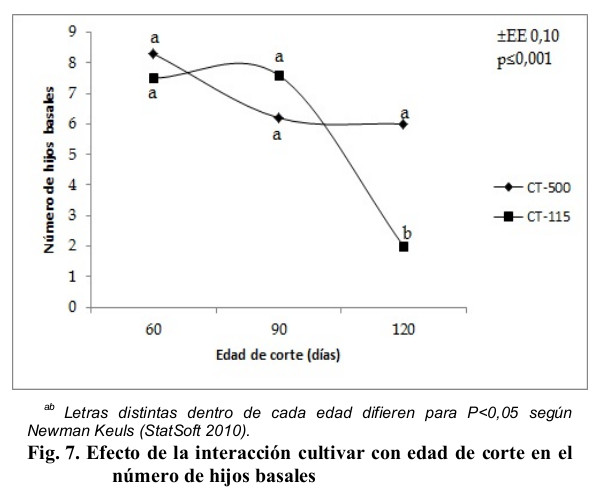
Muy relacionado con el comportamiento productivo, está la cantidad de hijos basales que puede generar una planta, en la Fig. 7 se muestra la interacción cultivar con edad de corte en esta variable, se apreció un comportamiento variable, sin embargo, los promedios de las edades 60 y 90 días no manifestaron diferencias estadísticas (P > 0,05), pero si con el valor obtenido por el CT-115 a la edad de 120 días.
La cantidad de hijos basales fue inferior a los obtenidos por Díaz (2007) en diferentes edades de rebrote (86, 112 y 154 días), y a los referidos por Martínez et al. (2010) cuando evaluaron al Cuba CT-169 y OM-22 en la estación lluviosa. El comportamiento reflejado en la Fig. 7 se debe principalmente a la movilización de sustancias de reservas dentro de la planta para generar hijos basales, en el caso del CT-500 para la edad de 60 días al corte, contenía un mayor contenido de carbohidratos solubles que fueron movilizados para el proceso de brotación de los hijos basales, según Ayala (1990) este proceso ocurre cuando comienza el envejecimiento del tallo, lo que sugiere una acelerada maduración por parte de esta variedad, sin embargo, para el CT-115 este proceso de maduración se identifica a los 90 días al corte para disminuir nuevamente a la edad de 120 días, debido a la culminación del crecimiento y movilización de los carbohidratos hacia las raíces para asegurar el rebrote (Fortes, 2014).
CONCLUSIONES
Se observó que los cultivares mostraron diferencias significativas (P < 0,001) para las edades de 60 y 90 días, mientras que, para el número de hijos basales y el porcentaje de materia seca en la planta íntegra, el CT-115 fue significativamente superior en las edades de 60 y 120 días, respectivamente. La tasa de crecimiento absoluto y duración de la biomasa se desfasaron significativamente a los 90 días a favor del CT-500. Ambas variedades demostraron el potencial productivo de materia seca por hectárea y solo se diferenciaron (P < 0,001) para la edad de 120 días en condiciones de áreas forrajeras con limitantes de salinidad en el suelo.
RECOMENDACIONES
Recomendamos considerar los resultados del presente estudio como datos preliminares para el diseño de la estructura varietal de áreas forrajeras con bajos niveles de salinización, y someter las nuevas variedades obtenidas por cultivos de tejidos a evaluación en áreas con mayores niveles de salinización.
REFERENCIAS BIBLIOGRÁFICAS
ÁLVAREZ, Y. (2009). Evaluación inicial de nuevas variedades de Pennisetum purpureum con tolerancia a la salinidad en el Valle del Cauto. Tesis de Maestría, Universidad de Granma. Cuba.
ARGENTEL MARTÍNEZ, L.; GARATUZA PAYÁN, J.; YÉPEZ GONZÁLEZ, E. A., & DE LOS SANTOS, S. (2016). Evaluación de la tolerancia de variedades mexicanas de trigo a la salinidad, a través de indicadores fisiológicos, bioquímicos y agronómicos, cultivadas en Cuba en condiciones de campo. Cultivos Tropicales, 37 (1), 91-101.
AYALA, J. R. (1990). Plantación y establecimiento. King grass. Plantación, establecimiento y manejo en Cuba. La Habana, Cuba: Ed. EDICA, Instituto de Ciencia Animal.
BARRANCO, G. y DÍAZ, L. R. (1989). Clima. Nuevo Atlas Nacional de Cuba. Cuba: Inst. Geogr. ACC, ICGC, MINFAR.
BENÍTEZ, D, G.; CISNEROS, M.; PÉREZ-INFANTE, F.; NUVIOLA, Y. (2010). El Balance alimentario para los rebaños en pastoreo. Holguín, Cuba: Ed. ARGRAF.
BENÍTEZ, D.; RAMÍREZ, ALINA.; DÍAZ, MARGARITA.; RAY, J.; GUERRA, J. y VEGAS, A. (2007). Comportamiento de machos vacunos en un sistema racional de pastoreo en el Valle del Cauto. Rev. Cubana Cienc. Agric., 41 (3), 227-230.
CALZADA-MARÍN, J. M.; ENRÍQUEZ-QUIROZ, J. F.; HERNÁNDEZ-GARAY, A.; ORTEGA-JIMÉNEZ, E. y MENDOZA-PEDROZA, S. I. (2014). Análisis de crecimiento del pasto maralfalfa (Pennisetum sp.) en clima cálido subhúmedo. Revista mexicana de ciencias pecuarias, 5 (2), 247-260.
CORDOVÍ, E.; RAY, J. V.; NHATUMBO, S. y CHIMBALAMBALA, A. (2013). Comportamiento de tres especies del género Pennisetum purpureus en clima semiárido del sur de Mozambique. Rev. Granma Ciencia, 17 (3), 1-6.
DÍAZ, D. (2007). Evaluación agronómica de nuevas variedades Pennisetum purpureum en condiciones de sequía el Valle del Cauto. Tesis de maestría, Universidad de Matanzas, Cuba.
FAJARDO, H. (2008). La innovación tecnológica en la producción pecuaria en la provincia de Granma: Una necesidad impostergable. Rev. Granma. Ciencia, 12 (1), 12-16.
FEBLES, G. y HERRERA, R.S. (2006). Introducción y características botánicas. Pennisetum purpureum para la ganadería tropical. La Habana, Cuba: Ed. EDICA.
FERNÁNDEZ, M. J.; VIAMONTE, I. M.; FONSECA, N. y RAMÍREZ, A. (2015). Evaluación de dos cultivares de Pennisetum purpureum tolerantes a la sequía en la región de Cauto Cristo, Granma, Cuba. Rev. Ciencia y Tecnología Ganadera, 9 (1), 23-29.
FORTES, D. (2014). Comportamiento de algunos indicadores morfofisiológicos y de calidad de Pennisetum purpureum vc. Cuba CT-115 utilizado como banco de biomasa. Tesis de Doctorado, Instituto de Ciencia Animal, Cuba.
HERNÁNDEZ, L.; RODRÍGUEZ, I.; CRESPO, G.; SANDRÍNO B y FRAGA, S. (2003). Los componentes de la fitomasa subterránea en diferentes pastizales de la provincia de La Habana. II Foro latinoamericano de Pastos y Forrajes, Instituto de Ciencia Animal, La Habana, Cuba.
HERNÁNDEZ, A.; PÉREZ, J. M.; BOSCH, D.; RIVERO, L. y CAMACHO, I. (2015). Nueva versión de la clasificación genética de los suelos de Cuba. Ciudad de La Habana, Cuba: Instituto de Suelos, Ministerio de la Agricultura, AGRINFOR.
HERRERA, R. S. (2006). La fisiología vegetal y los pastos. Conferencias curso de pastos, ICA, La Habana, Cuba.
HERRERA, R. S. (2000). Obtención de plántulas de Pennisetum purpureum con resistencia a la sequía y salinidad mediante técnicas biotecnológicas. La Habana, Cuba: CITMA-ICA.
HERRERA, R. S.; CHAPLE, Z.; CRUZ, A. M.; ROMERO, A. y GARCÍA, M. (2003). Obtención de plántulas de Pennisetum purpureum resistentes a la sequía y salinidad. Nota técnica. Rev. Cubana de Cienc. Agríc.,37 (2), 89-93.
KEULS, M. (1952). The use of the "studentized range" in connection with an analysis of variance. Euphytica, 1 (2), 112-122.
LEDEA, J. L. (2016). Caracterización químico nutritiva de nuevas variedades de Cenchrus purpureus tolerantes a la sequía en el Valle del Cauto. Tesis de Maestría, Universidad de Granma, Cuba.
MASSEY, F. J. (1951). The Kolmogorov-Smirnov test for goodness of fit. Journal of the American statistical Association, 46 (253), 68-78.
MARTÍNEZ, R. O.; TUERO, R.; TORRES, V., & HERRERA, R. S. (2010). Models of Biomass Accumulation and Quality in Varieties of Elephant Grass, Cuba CT-169, OM - 22 and King Grass during the Rainy Season in the Western Part of Cuba. Cuban J. Agric. Sci., 44 (2), 187.
NEVO, E. y CHEN, G. (2010). Drought and Salt Tolerances in Wild Relatives for Wheat and Barley Improvement. Plant, Cell & Environment, 33 (4), 670-685.
ONEI (2015). Anuario estadístico de Cuba 2014. Territorio. Cuba: Oficina Nacional de Estadística e Información. Recuperado el 12 de enero de 2015, desde http://www.one.cu/anuarioestadisticodecuba2014
PEÑA, F. H.; ÁLVAREZ, A. y DÍAZ, E. (2002). IV Clasificación de los suelos del Valle del Cauto. Rev. Granma Ciencia, 6 (3), 56-60.
PÉREZ-INFANTE, F. (2013). Ganadería eficiente. La Habana, Cuba: Editorial: Asociación Cubana de Producción Animal.
RAMÍREZ, A. (2010a). Caracterización y tipificación de las fincas ganaderas en ecosistemas montañosos del macizo Sierra Maestra. Tesis de Doctorado, Instituto de Ciencia Animal, Cuba.
RAMÍREZ, J. L. (2010b). Rendimiento y calidad de cinco gramíneas en el Valle del Cauto. Tesis de Doctorado, Instituto de Ciencia Animal, Cuba.
REYES, J. J.; PADILLA, C.; MARTÍN, P. C.; GÁLVEZ, M.; REY, S.; NODA, A. y REDILLA, C. (2015). Consumo de forrajes tropicales por vacas lecheras, mestizas Siboney, manejadas en condiciones de estabulación. Avances en Investigación Agropecuaria, 19 (1), 31-40.
RINCÓN, Á.; LIGARRETO, G. y SANJUANELO, D. (2007). Crecimiento del maíz y los pastos (Brachiaria sp.) establecidos en monocultivo y asociados en suelos ácidos del piedemonte llanero colombiano. Agronomía Colombiana, 25 (2), 264-272.
RONCALLO, B.; SIERRA, A. M. y CASTRO, E. (2012). Forage Yield of Cutting Grass and Effect on Compositional Quality and Yield of Milk in a Dry Caribbean. Revista Corpoica Ciencia y Tecnología Agropecuaria, 13 (1), 71-78.
ROSENZWEIG, C.; ELLIOTT, J.; DERYNG, D.; RUANE, A. C.; MÜLLER, C.; ARNETH, A.; et al. (2014). Assessing Agricultural Risks of Climate Change in the 21st Century in a Global Gridded Crop Model Intercomparison. Proceedings of the National Academy of Sciences, 111 (9), 3268-3273.
SCHROEDER, J. I.; DELHAIZE, E.; FROMMER, W. B.; GUERINOT, M. L.; HARRISON, M. J.; HERRERA-ESTRELLA, L. et al. (2013). Using Membrane Transporters to Improve Crops for Sustainable Food Production. Nature, 497 (7447), 60-66.
UVIDIA, H.; LEONARD, I.; BENÍTEZ, D. y BUESTAN, D. (2013). Dinámica del crecimiento de la Maralfalfa (Pennisetum sp.), en condiciones de la Amazonia Ecuatoriana. Revista Amazónica Ciencia y Tecnología, 2 (1), 14-18.
VALLES DE LA MORA, B.; CASTILLO, E. y BERNAL, H. (2016). Rendimiento y degradabilidad ruminal de la materia seca y energía de diez pastos tropicales cosechados a cuatro edades. Rev. Mex. Cienci. Pec., 7 (2), 141-158.
Recibido: 12-7-2017
Aceptado: 20-7-2017
José Leonardo Ledea Rodríguez, Instituto de Investigaciones Agropecuarias Jorge Dimitrov, Estación Experimental de Pastos y Forrajes, Bayamo, Granma, Cuba. ledea@dimitrov.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}