










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Producción Animal
versión On-line ISSN 2224-7920
Rev. prod. anim. vol.30 no.2 Camagüey mayo.-ago. 2018
ACUICULTURA
Probióticos: una realidad en el cultivo de camarones. Artículo de revisión
Probiotics, a Fact in Shrimp Culture
Adrián Toledo*, Néstor M. Castillo**, Olimpia Carrillo**, Amilcar Arenal*
* Laboratorio de Bioquímica, Departamento de Morfofisiología, Facultad de Ciencias Agropecuarias, Universidad de Camagüey, Cuba
** Departamento de Bioquímica, Facultad de Biología, Universidad de La Habana, Cuba. adrian.toledo@reduc.edu.cu
RESUMEN
La camaronicultura es uno de los sectores más lucrativos y de mayor crecimiento dentro de la acuicultura marina; sin embargo, la intensificación del cultivo para satisfacer una demanda cada vez mayor ha incrementado la incidencia de enfermedades produciendo pérdidas económicas considerables. La prevención y control de los brotes se basa fundamentalmente en antimicrobianos, lo cual es criticado por la acumulación de residuos en el ambiente, el desarrollo de resistencia y poca aceptación de los productos por parte de los consumidores. Alternativamente, se proponen métodos más amigables con el medio ambiente, como la aplicación de probióticos, un procedimiento de gran versatilidad y considerables beneficios ampliamente aceptado en la producción de camarón a nivel mundial. Los probióticos son capaces de controlar patógenos por múltiples mecanismos, promover el crecimiento del hospedero y mejorar la calidad del ambiente de cultivo. Adicionalmente pueden ser administrados por varias vías y en combinación con otras sustancias beneficiosas. Este artículo provee una actualización sobre la aplicación de probióticos en el cultivo de camarón, con énfasis en su influencia en el incremento de la productividad.
Palabras clave: camaronicultura, productividad, camarón, probiótico, mecanismo de acción.
ABSTRACT
Shrimp culture is one of the most lucrative and fast-growing sectors in marine aquaculture. However, culture intensification to meet the growing demands has increased the incidence of diseases, causing considerable economic losses. Outbreak prevention and control is mostly based on antimicrobial antibiotics, which is very controversial due to the accumulation of residues in the environment, increased resistance, and less consumer acceptance. As an alternatively, more environment-friendly methods are being suggested. The application of probiotics is a versatile procedure with broadly accepted benefits in shrimp production worldwide. Probiotics can control pathogens by means of multiple mechanisms, which can promote host growth and improve the quality of culture environment. Additionally, they can be administered through different routes in combination with other beneficial substances. This article offers an update on probiotic use in shrimp culture, with particular emphasis on production increase.
Keywords: shrimp culture, productivity, shrimp, probiotic, action mechanism.
INTRODUCCIÓN
La acuicultura constituye uno de los sectores de producción de alimentos de mayor crecimiento en las últimas décadas, y la producción de crustáceos una de las más ventajosas, de más rápidos logros y de mayor valor monetario (Stentiford et al., 2012). El camarón es un producto de alto valor, que se produce principalmente en Asia y América Latina, donde su cultivo genera riquezas y puestos de trabajo (FAO, 2004). Las especies más cultivadas en estas áreas son Litopenaeus vannamei (camarón blanco del pacífico) y Penaeus monodon (camarón tigre negro) (Wang y Gu, 2010). Aun cuando el sector tiene una tendencia de crecimiento favorable, existen obstáculos como la disponibilidad de materias primas para la elaboración de alimentos y el aumento de enfermedades (Stentiford et al., 2012).
En la producción a gran escala, donde los animales están expuestos a condiciones de estrés, el deterioro de las condiciones óptimas de cultivo conduce a la aparición de enfermedades que provocan pérdidas económicas considerables (Mohapatra et al., 2013). El control de enfermedades en la acuicultura, tanto profiláctico como terapéutico, se basó históricamente en el uso de antimicrobianos (Cabello et al., 2013); sin embargo, esta práctica es ampliamente criticada en la actualidad por su impacto en la acumulación de residuos en el ambiente y el desarrollo de resistencia, lo que también afecta la aceptación de los productos por parte de los consumidores (Gothwal y Shashidhar, 2015; Kumar et al., 2016; Liu et al., 2017). Por otro lado, se requiere de buenas prácticas acuícolas que garanticen un mejor aprovechamiento del alimento por parte de los animales con el fin de incrementar la productividad del proceso.
La administración de microorganismos para aumentar la resistencia a enfermedades y mejorar el estado nutricional de los camarones es un método amigable con el medio ambiente y más seguro (Martínez?Córdova et al., 2015). Los probióticos son microorganismos vivos que ejercen un efecto beneficioso sobre el hospedero, modifican la comunidad microbiana asociada a este o la del ambiente, y permiten un mejor aprovechamiento del alimento o potencian su valor nutricional, estimulan su respuesta hacia enfermedades o mejoran la calidad de su entorno (Verschuere et al., 2000).
En la actualidad existen varios productos comerciales que se emplean como probióticos (Miandare et al., 2016; Xue et al., 2016; Ferreira et al., 2017; Javadi y Khatibi, 2017); no obstante, el aislamiento y caracterización de nuevas cepas constituye un campo de investigación activo, en particular, aquellas aisladas del ambiente y/o del hospedero de interés (Wang y Gu, 2010; Franco et al., 2016a; Franco et al., 2016b).
El propósito de este trabajo es realizar una revisión sobre los recientes avances en la aplicación de probióticos en el cultivo de camarón con énfasis en su influencia en el incremento de la productividad.
DESARROLLO
Mecanismos de acción
Los probióticos ejercen su efecto beneficioso mediante múltiples mecanismos, no sólo sobre el organismo de interés sino también sobre el ambiente que le rodea. Entre los principales mecanismos de acción descritos para probióticos que se emplean en la acuicultura se incluyen: la capacidad para colonizar y adherirse al tracto intestinal, la modulación del sistema inmune, la producción de compuestos benéficos, la producción de sustancias antagónicas contra patógenos y la mejora de la calidad del medio acuático.
Colonización y adhesión en el tracto gastrointestinal
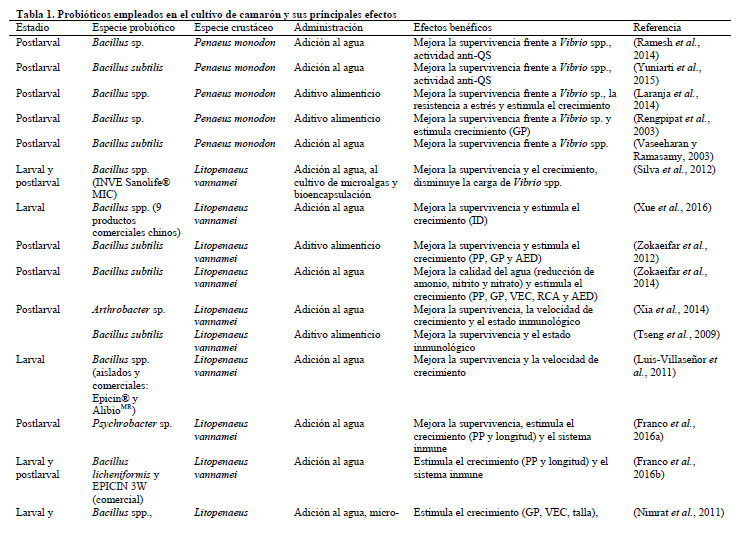
La habilidad de las bacterias para adherirse y sobrevivir en el mucus entérico es decisiva en el establecimiento de la microbiota intestinal. La capacidad de adherencia es una característica que es aprovechada de igual manera, tanto por las bacterias probióticas como por las patógenas. En el caso de las probióticas, es uno de los criterios más importantes para su selección y aplicación en acuicultura (Lamari et al., 2014; Vieira et al., 2016), mientras que para las patógenas la habilidad para adherirse se relaciona con la virulencia y se considera como el primer paso para una infección (Defoirdt, 2014). En la acuicultura, la información disponible indica que las bacterias aisladas de animales cultivados o de su entorno tienen mayor capacidad de adhesión al mucus gastrointestinal y a los tejidos, respecto a bacterias foráneas. Por tal razón la acción de muchos probióticos suele ser transitoria y requieren ser administrados de forma continua, ya sea como suplemento en el alimento o a través del agua de cultivo para que mantengan su efecto biológico, a menos que sean desarrollados a partir de cepas aisladas del propio ecosistema acuático (Nimrat et al., 2011) (Tabla 1); no obstante, se documentó que aislados microbianos de un organismo pueden colonizar otras especies cultivadas, lo que indica así la falta de especificidad para la colonización del tracto digestivo (Sánchez-Ortiz et al., 2016).
Producción de antimicrobianos y compuestos antivirales
Los microorganismos con actividad probiótica también pueden tener la capacidad de generar productos extracelulares que inhiben o matan otras bacterias potencialmente patógenas: sustancias antimicrobianas (Pham et al., 2014; Ming et al., 2015), ácidos orgánicos (Tejero-Sariñena et al., 2012; Fakruddin et al., 2017) y bacteriocinas (Iyapparaj et al., 2013; Muñoz-Atienza et al., 2013; Ming et al., 2015).
Los probióticos no sólo tienen capacidad antibacteriana, también se describe actividad antiviral de algunos aislados como Pseudomonas sp., Vibrio sp. y Aeromonas sp., contra el virus de la necrosis hematopoyética (IHNV) (Kamei et al., 1988). Maeda et al. (1997) aislaron una cepa de Pseudoalteromonas undina, que ejerció efectos antivirales e incrementó la supervivencia en camarón (Penaeus sp.) infectado experimentalmente con el virus de la necrosis neuro Sima-aji (SJNNV), Baculo e Iridovirus. Sánchez-Ortiz et al. (2016) hallaron que la administración de Bacillus spp. en la dieta de L. vannamei naturalmente infectados con virus del síndrome de la mancha blanca (WSSV) y IHNV reducía la prevalencia viral en los animales y estimulaba el crecimiento y la expresión de genes del sistema inmune como el pro-fenoloxidasa (proPO) y el superóxido dismutasa (SOD), lo que influyó en un mayor índice de sobrevivencia respecto a los grupos no tratados (Tabla 1).
La fagocitosis y la apoptosis son los principales mecanismos descritos de la respuesta inmune antiviral en camarón (Wang y Zhang, 2008), aunque también existen reportes de la efectividad de estrategias terapéuticas basadas en ARN interferentes y de la estimulación de la respuesta inmune innata de camarón en respuesta a proteínas de envoltura virales (Thomas et al., 2014; Taju et al., 2015). No obstante, la aplicación práctica de algunos de estos hallazgos no resulta factible a nivel productivo. La selección y empleo de probióticos, reconocidos potenciadores de la inmunidad celular, con este fin es una alternativa promisoria.
Hasta la fecha se han descrito escasos métodos para detectar, específicamente, la actividad antiviral de probióticos y su mecanismo de acción. Algunos de los ensayos propuestos incluyen: el pretratamiento de las células con el probiótico; la coincubación del probiótico y el virus; la adsorción del virus al probiótico y el efecto antiviral de sobrenadantes de cultivo (Lakshmi et al., 2013). Recientemente se reportó una nueva línea eucariótica como modelo para este fin (Boti? et al., 2007), que se ha aplicado con éxito en la virología humana, pero no en la acuicultura (Lakshmi et al., 2013). La estandarización de estas técnicas para el estudio de la interacción probiótico-virus en la camaronicultura constituye una necesidad impostergable para el desarrollo de estrategias de selección de nuevas cepas y de terapias más racionales y efectivas.
Producción de compuestos benéficos
Las bacterias marinas y las levaduras pueden llegar a constituir una fuente de proteínas importante en el mejoramiento del aporte nutricional de algunas especies acuáticas cultivadas (Achupallas et al., 2015; Melo et al., 2015; Gamboa-Delgado et al., 2016; Qiu y Davis, 2017).
De forma similar, los lípidos producidos por microorganismos marinos se indican como sustancias de gran importancia para la nutrición de especies acuáticas (Hoseinifar et al., 2016). La producción de enzimas como lipasas, quitinasas y proteasas por parte de microorganismos seleccionados, pueden contribuir al proceso digestivo de los organismos cultivados e impactar, positivamente, en su comportamiento productivo (Shen et al., 2010; Zokaeifar et al., 2012; Chai et al., 2016; Seenivasan et al., 2016; Xue et al., 2016) (Tabla 1).
Mejora de la calidad del agua
Las bacterias Gram positivas, principalmente del género Bacillus, seleccionadas como probióticos pueden convertir la materia orgánica en CO2; en contraste, las bacterias Gram-negativas se caracterizan por convertir materia orgánica en biomasa bacteriana o limo (Dalmin et al., 2001; Zokaeifar et al., 2014). Mujeeb Rahiman et al. (2010) aplicaron aislados Bacillus sp. y Vibrio sp. como aditivos a la dieta y al agua de cultivo de Macrobrachium rosenbergii a diferentes dosis y frecuencias de administración, lo que resultó en la reducción de la concentración de amonio y nitrato del medio, el incremento significativo de la supervivencia, el crecimiento y la estimulación del sistema inmune. Al aplicar el producto comercial EM (EM®, Japón), compuesto por bacterias ácido-lácticas y levaduras, al agua de un cultivo intensivo de L. vannamei, Melgar Valdés et al. (2013) encontraron que el tratamiento redujo la materia orgánica, la concentración de nitratos, reguló el pH e incrementó la disponibilidad de fósforo en el agua y mejoró indicadores de productividad como la supervivencia y el factor de conversión alimentaria (Tabla 1).
No obstante, algunos estudios informan que la aplicación de probióticos no mejora los parámetros evaluados en el cultivo de camarón (Silva et al., 2012; Bolívar Ramírez et al., 2013). Estos resultados sugieren que la frecuencia de aplicación y las dosis empleadas, así como el sistema de producción (en estanques al aire libre o en condiciones más controladas) influyen en dicho efecto. Este asunto es de particular interés si se toma en cuenta que el desarrollo de los animales y los parámetros de calidad del agua resultan menos afectados en la medida que los cultivos se acercan más a las condiciones del medio natural. La calidad microbiológica del agua constituye un factor de riesgo para la aparición de brotes, de ahí que otro blanco de la aplicación de probióticos sea reducir la prevalencia de patógenos oportunistas en el ambiente (Chumpol et al., 2017). La adición de Bacillus sp. como suplemento en la dieta de P. monodon disminuyó la carga de Vibrio sp. en el estanque, lo cual favoreció la prevalencia de bacterias heterotróficas (Boonthai et al., 2011). Silva et al. (2012) evaluaron la adición de un producto comercial compuesto por Bacillus spp. en diferentes estadios de L. vannamei y demostraron que el tratamiento disminuyó la carga de Vibrio sp., tanto en el intestino de los animales como el agua.
Adicionalmente, se han propuesto nuevas tecnologías basadas en microorganismos que contribuyen al mantenimiento de condiciones óptimas de cultivo, entre ellas, el "Biofloc" propone estimular el desarrollo y prevalencia de comunidades microbianas heterotróficas en el medio de cultivo capaces de remover la materia orgánica mediante la adición de fuentes de carbono (Crab et al., 2012). También se explota el potencial de esta tecnología en la generación de biomasa microbiana como fuente alternativa de proteínas (Ahmad et al., 2017). Existen varios reportes de la efectividad de este procedimiento, así como artículos de revisión que discuten en detalle sus particularidades (Crab et al., 2012; Ekasari et al., 2014; Kim et al., 2015; Melo et al., 2015; Suita et al., 2015; Bossier et al., 2016; Ahmad et al., 2017; Ferreira et al., 2017).
Inmunomodulación
La defensa frente a patógenos en crustáceos se basa fundamentalmente en mecanismos de la inmunidad innata (Song y Li, 2014). El sistema inmune de los camarones involucra a hemocitos (para la encapsulación, formación de nódulos y fagocitosis), a varios componentes plasmáticos (péptidos antimicrobianos, histonas, enzimas lisosomales, proteínas de unión a lipopolisacáridos y B-1,3 glucanos y moléculas de reconocimiento) y sistemas multiméricos (proteínas de la cascada de coagulación y sistema pro-fenoloxidasa) (Aguirre-Guzmán et al., 2009). La incidencia cada vez más frecuente de brotes de enfermedades y las pérdidas económicas que traen consigo, motivan el estudio de estos mecanismos de defensa, pues ofrecen novedosas alternativas para el manejo de enfermedades (Aguirre-Guzmán et al., 2009).
Huang et al. (2013), en ensayos de infección experimental de L. vannamei con V. harveyi demostraron que los animales resistentes mostraban una respuesta inmune más rápida, de mayor magnitud y efectividad en la eliminación del patógeno respecto a animales normales. Lo que sugiere que la estimulación del sistema inmune mediante el incremento de los niveles basales de algunos de sus componentes, puede ser relevante en la eliminación del agente infeccioso y el mantenimiento de la homeostasia. La aplicación de probióticos en la camaronicultura con el fin de estimular el sistema inmune es una de las áreas investigativas que más se explora. En tal sentido, existen varias publicaciones que documentan cómo los indicadores del estado inmunológico del camarón son modulados por probióticos (Tabla 1) (Mujeeb Rahiman et al., 2010; Shen et al., 2010; Wang y Gu, 2010; Zokaeifar et al., 2014; Franco et al., 2016b).
El sistema pro-fenoloxidasa (proPO) es uno de los principales componentes del sistema inmune de peneidos, cuyo último paso en la cascada enzimática culmina con la activación de la enzima fenoloxidasa mediante proteólisis, la cual produce derivados fenólicos tóxicos y melanina. Esta sustancia, muy conservada evolutivamente, tiene actividad microbicida y contribuye a confinar al patógeno al sitio de entrada. La superóxido dismutasa (SOD) es uno de los principales mecanismos de defensa contra el estrés oxidativo causado por la contaminación, infecciones, hipoxia, hiperoxia, temperatura e inmunoestimulantes (Neves et al., 2000). También se reconoce la importancia de las enzimas peroxidasa y catalasa frente a especies reactivas del oxígeno (Castex et al., 2010; Sánchez Ortiz et al., 2013); por otro lado, la relevancia de enzimas bacteriolíticas como la lisozima en la defensa contra bacterias patógenas está bien documentada (Burge et al., 2007; Karthik et al., 2014). La actividad enzimática y la expresión de la lisozima en camarones puede estimularse por la adición de probióticos, tanto en infecciones experimentales (Maeda et al., 2013) como en estudios a escala productiva (NavinChandran et al., 2014; Miandare et al., 2016).
La cuantificación de la actividad enzimática y la expresión génica de las proteínas involucradas en la respuesta inmune son analizadas con frecuencia como indicadores del estado inmunológico del camarón (Miandare et al., 2016; Sánchez-Ortiz et al., 2016). Zokaeifar et al. (2014) evaluaron, en condiciones experimentales, el comportamiento de la expresión de genes del sistema inmune en L. vannamei tratados durante ocho semanas con Bacillus subtilis L10 y G1, y luego infectados con Vibrio harveyi. Así, obtuvieron que la expresión de proPO y otros genes de proteínas de unión a patrones moleculares de patógenos se incrementó, respecto a los controles pertinentes. También encontraron que disminuía la mortalidad acumulada entre los animales tratados respecto al control (36,7-50 % vs 80 % para el control). Estos resultados indican que el incremento de la expresión de los genes evaluados está involucrado con la aplicación de probióticos, lo que condujo a una mayor resistencia en los animales. Wang y Gu (2010) realizaron un estudio en tanques de 12 500 L por un periodo de 35 días, en el que evaluaron el efecto de Lactobacillus acidophilus RS058, Rhodopseudomonas palustris GH642 y Bacillus coagulans NJ105 en el crecimiento y respuesta inmune de juveniles de L. vannamei. La actividad enzimática PO y SOD, así como los parámetros de crecimiento evaluados resultaron mayores en los camarones tratados con las cepas probióticas, respecto a sus controles; sin embargo, la actividad de la enzima peroxidasa no mostró diferencias entre los mencionados grupos.
La inmunidad celular es otro de los parámetros de interés en relación con el estado inmunológico de los peneidos, pues el incremento del número de hemocitos circulantes se relaciona con una mayor capacidad para fagocitar y eliminar agentes extraños (Sánchez-Ortiz et al., 2015). Xia et al. (2014) al administrar la cepa Arthrobacter sp. CW9 al agua de cultivo de L. vannamei en un ensayo que duró 24 días, detectaron incremento en la actividad fagocítica, mayor efectividad en la eliminación de patógenos, así como mejor crecimiento y sobrevivencia comparado con el grupo control. Resultados similares describieron NavinChandran et al. (2014) para P. monodon tratado con Bacillus cereus.
Muchos estudios recientes proponen el empleo de inmunoestimulantes teniendo como antecedentes los resultados de la administración de probióticos o sus derivados; sin embargo, algunos autores advierten sobre los efectos adversos de la inmunoestimulación prolongada en el camarón (Smith et al., 2003).
Actividad anti-Quorum sensing
El antagonismo contra patógenos es uno de los criterios de selección más empleados para la obtención de nuevas cepas con potencial probiótico (Bright Singh et al., 2014; Shazwani et al., 2015). No obstante, algunos investigadores proponen un enfoque alternativo que consiste en reducir la virulencia del patógeno sin comprometer directamente su crecimiento (Czajkowski y Jafra, 2009; Brackman et al., 2011; Defoirdt et al., 2012). Esta perspectiva, denominada terapia antivirulencia, se basa en que la expresión de muchos de los genes involucrados en la patogenicidad en bacterias está regulada por el quorum sensing (QS): un proceso de comunicación bacteriana célula-célula mediado por moléculas señales de bajo peso molecular que provocan respuestas dependientes de la densidad poblacional (Defoirdt et al., 2010). La interferencia del QS o quorum quenching podría permitir el control de las enfermedades bacterianas con una menor tendencia al desarrollo de resistencia y a la alteración de la microbiota normal del hospedero, respecto a los antimicrobianos (Tabla 1) (Defoirdt, 2016). Las estrategias descritas con este fin incluyen el empleo de inhibidores y enzimas degradadoras de AHL (acil-homoserín-lactonas; moléculas señal inductoras) (Brackman et al., 2008; Pande et al., 2013; Torres et al., 2013; Pande et al., 2015; Torres et al., 2016). Las enzimas degradadoras de AHL tienen amplia distribución en bacterias, en especial en especies del género Bacillus (Defoirdt et al., 2011). Ramesh et al. (2014) aislaron cepas de Bacillus a partir de intestino de Penaeus monodon a las cuales evaluaron la actividad contra Vibrio spp. y anti-QS. De los 12 aislados obtenidos sólo APV03 y APV07 mostraron ambos mecanismos. La administración de las cepas, solas o en combinación, protegieron a postlarvas y juveniles de P. monodon frente a la infección por Vibrio harveyi, mientras que la actividad quorum quenching de ambas cepas fue evaluada in vitro frente a la cepa indicadora Chromobacterium violaceum. Yuniarti et al. (2015) demostraron que una cepa aislada de intestino de Penaeus monodon, identificada como B. subtilis y productora de enzimas degradadoras de AHL, protegía a juveniles de P. monodon frente a una infección con V. harveyi. La cepa crecida en cocultivo con V. harveyi fue capaz de inhibir el crecimiento del patógeno y reducir la concentración de AHL en el medio en ensayo in vitro; sin embargo, los resultados in vitro no se correspondieron con los observados en el ensayo de protección in vivo, puesto que no se observó diferencias entre las concentraciones de AHL entre los grupos tratados con B. subtilis y el control, aunque sí se observó tendencia a disminuir la concentración de AHL con el incremento de la concentración de B. subtilis en el medio.
Modos de administración
Los probióticos ejercen sus efectos benéficos, principalmente a nivel del tracto gastrointestinal, de ahí que muchos de los modos de administración desarrollados estén dirigidos a incrementar su estabilidad y facilitar su asimilación (Tabla 1). La adición de los probióticos en la dieta es una de las formas más empleadas (Shen et al., 2010; Boonthai et al., 2011; Liu et al., 2014); por esta vía los probióticos se incorporan simultáneamente con el alimento, lo que está relacionado con una contribución enzimática a la digestión y mejor aprovechamiento de los nutrientes ingeridos (Zokaeifar et al., 2012; Nimrat et al., 2013). También debe tenerse en cuenta que la periodicidad de la aplicación del probiótico permite mantener un balance favorable de microorganismos benéficos en el intestino del camarón que compiten con otros colonizadores intestinales como Vibrio spp. y otros patógenos (Luis-Villasenor et al., 2013); no obstante, este tipo de administración expone a los microorganismos probióticos a condiciones físicas y químicas extremas que podrían afectar su viabilidad y disminuir su efecto en el hospedero (Nimrat et al., 2011).
La microencapsulación es un método alternativo que consiste en recubrir las células con una matriz de polímeros, principalmente alginatos, lo que permite extender los periodos de almacenamiento de los cultivos, mejorar su viabilidad en los alimentos y en el tracto intestinal de sus hospederos, así como protegerlos de bacteriófagos (Nimrat et al., 2012). Algunos alimentos vivos de alto valor nutritivo y amplio uso en la camaronicultura como los rotíferos y Artemia spp. también se han evaluado como vehículo para la administración de probióticos (Hadiroseyani y Sutanti, 2014; Jamali et al., 2015). Este proceso, denominado bioencapsulación, aprovecha la capacidad filtradora de estos organismos que incorporan los probióticos al ser agregados al medio de cultivo. Aun cuando varios autores refieren la funcionalidad de las encapsulaciones (Ziaei-Nejad et al., 2006; Nimrat et al., 2011; Jamali et al., 2015), su moderada influencia en los parámetros productivos del cultivo de camarón, así como su complejo escalado y considerable costo sugieren la necesidad de estudios de rentabilidad para su aplicación en la fase productiva (Kumar et al., 2016). Por otro lado, los probióticos pueden ser agregados directamente al agua donde se cultivan los animales, sobre todo para el caso de aquellos capaces de remover la materia orgánica (Dalmin et al., 2001) y sustancias tóxicas que mejoran la calidad del medio acuático (Wang y Gu, 2010; Silva et al., 2012; Laranja et al., 2014; Franco et al., 2016b).
Influencia de probióticos en parámetros productivos del cultivo de camarón
La prevención y control de enfermedades en el cultivo de camarón, sobre todo en estadios larvales, constituye uno de los puntos críticos del proceso productivo. En tal sentido, la actividad antagónica de los probióticos ha ganado aceptación en el mejoramiento de la supervivencia de los animales (Vaseeharan y Ramasamy, 2003; Luis-Villaseñor et al., 2011). Balcázar y Rojas-Luna (2007) demostraron el potencial de la cepa Bacillus subtilis UTM 126, aislada de intestino de L. vannamei, en el control de V. harveyi en la camaronicultura. El tratamiento de juveniles de L. vannamei con una dieta suplementada con 105 UFC.g-1 de la mencionada cepa disminuyó la mortalidad acumulada hasta 18,25 % en relación a 51,75 % para el grupo control.
Sin embargo, aunque muchos estudios demuestran la protección que resulta de la administración de probióticos en una etapa específica de crecimiento, ya sea larval o postlarval, existen escasas referencias que investiguen su efecto durante la ontogenia del camarón y su importancia en la producción. Ziaei-Nejad et al. (2006) evaluaron el efecto de la aplicación de un probiótico comercial compuesto por Bacillus spp. en el desarrollo y la supervivencia de Fenneropenaeus indicus en varias etapas de desarrollo (Tabla 1). Al comparar los efectos de las distintas formas de administración del producto (adición al agua o bioencapsulado en Artemia) en los estadios comprendidos entre M-I y PL14, concluyeron que Bacillus spp. colonizaron el tracto digestivo de los animales, lo que contribuyó a una actividad enzimática digestiva (proteasa, lipasa y amilasa), peso húmedo y supervivencia superiores con relación a los respectivos controles; no obstante, las distintas vías de administración del probiótico evaluadas no mostraron diferencias significativas ni en la supervivencia ni en el peso, aun cuando los conteos de Bacillus spp. fueron ligeramente superiores en las larvas tratadas con Artemia enriquecida. Además, se evidenció que la aplicación de probióticos en estadios larvales fue determinante para mejorar los parámetros de crecimiento de la fase productiva en estanques, dado que la aplicación del probiótico por adición al agua resultó en una baja colonización del intestino de los camarones en su estado adulto. En otro estudio que involucró estadios larvales y postlarvales de L. vannamei realizado por Franco et al. (2016b), se compararon los efectos en la inmunidad y calidad larval de la cepa CIGBC-232, aislada de intestino de camarones saludables, y el probiótico comercial EPICIN 3W en condiciones de producción. Los resultados de esta investigación revelaron que la aplicación de CIGBC-232 redujo significativamente la carga de Vibrio spp. en los tanques, respecto al producto comercial e incrementó el peso y talla de los animales.
Por otro lado, Rengpipat et al. (2003) evaluaron el efecto de la inclusión de un probiótico en la dieta de P. monodon a nivel productivo en estanques de fondo de tierra durante 100 días en las estaciones caliente y fría. La administración de la comida suplementada resultó en mayor supervivencia y talla de los animales tratados, respecto al control en ambas estaciones. La aplicación del probiótico como aditivo alimentario en el ensayo resultó en 30 % más de ganancia de peso diaria (GPD) y 28 % más de supervivencia, respecto al control. La mejora de estos parámetros permitió que el rendimiento anual estimado (dos ciclos de cultivo de 100 días) fuera 49 % superior en camarones alimentados con la dieta suplementada; mientras que Melgar Valdés et al. (2013) obtuvieron que la aplicación de un probiótico comercial mejoró los parámetros de producción de L. vannamei, así como la calidad del agua del estanque en condiciones de cultivo intensivas.
Si bien la mayoría de las publicaciones informan los efectos positivos de la utilización de probióticos en la camaronicultura, algunos autores refieren la ineficaz o escasa actividad de productos comerciales a escala productiva (Xue et al., 2016). De ahí que se requiera la comprobación de calidad de los productos comerciales, así como la determinación de la dosis óptima y su modo de empleo antes de su introducción en la producción.
CONCLUSIONES
Los probióticos intervienen en los parámetros productivos del camarón relacionados con un mayor aprovechamiento de nutrientes, un sistema inmune potenciado y mayor supervivencia de los animales. La fácil manipulación y seguridad hacen de la aplicación de probióticos una práctica cada vez más aceptada, pues mejora las condiciones del medio de cultivo y ofrecen ventajas para la expansión y perfeccionamiento de una camaronicultura sostenible.
REFERENCIAS BIBLIOGRÁFICAS
CASTEX, M.; LEMAIRE, P.; WABETE, N. y CHIM, L. (2010). Effect of Probiotic Pediococcus acidilactici on Antioxidant Defences and Oxidative Stress of Litopenaeus stylirostris under Vibrio nigripulchritudo Challenge. Fish Shellfish Immunol, 28 (4), 10-16.
CHAI, P. C.; SONG, X. L.; CHEN, G. F.; XU, H. y HUANG, J. (2016). Dietary Supplementation of Probiotic Bacillus PC465 Isolated from the Gut of Fenneropenaeus chinensis Improves the Health Status and Resistance of Litopenaeus vannamei Against White Spot Syndrome Virus. Fish Shellfish Immunol, 54 (2), 2-10.
DEFOIRDT, T.; BOON, N. y BOSSIER, P. (2010). Can Bacteria Evolve Resistance to Quorum Sensing Disruption? Plos Pathogens, 6 (1), 13-20.
DEFOIRDT, T.; SORGELOOS, P. y BOSSIER, P. (2011). Alternatives to Antibiotics for the Control of Bacterial Disease in Aquaculture. Curr. Opin. Microbiol., 14 (3), 51-58.
MELGAR VALDÉS, C. E.; BARBA MACÍAS, E.; ÁLVAREZ-GONZÁLEZ, C. A.; TOVILLA HERNÁNDEZ, C. y SÁNCHEZ, A. J. (2013). Microorganisms Effect with Probiotic Potential in Water Quality and Growth of the Shrimp Litopenaeus vannamei (Decapoda: Penaeidae) in Intensive Culture. Rev Biol Trop, 61 (3), 1215-1228.
NIMRAT, S.; TANUTPONGPALIN, P.; SRITUNYALUCKSANA, K.; BOONTHAI, T. y VUTHIPHANDCHAI, V. (2013). Enhancement of Growth Performance, Digestive Enzyme Activities and Disease Resistance in Black Tiger Shrimp (Penaeus monodon) Postlarvae by Potential Probiotics. Aquaculture International, 21 (1), 655-666.
Shen, W.-Y.; Fu, L.-L.; Li, W.-F. y Zhu, Y.-R. (2010). Effect of Dietary Supplementation with Bacillus subtilis on the Growth, Performance, Immune Response and Antioxidant Activities of the Shrimp (Litopenaeus vannamei). Aquaculture Research, 41 (11), 91-98.
SILVA, E. F.; SOARES, M. A.; CALAZANS, N. F.; VOGELEY, J. L.; DO VALLE, B. C.; SOARES, R. et al. (2012). Effect of probiotic (Bacillus spp.) Addition During Larvae and Postlarvae Culture of the White Shrimp Litopenaeus vannamei. Aquaculture Research, 44 (1), 13-21.
SMITH, V. J.; BROWN, J. H. y HAUTON, C. (2003). Immunostimulation in Crustaceans: Does it Really Protect Against Infection? Fish & Shellfish Immunology, 15, 71-90.
SONG, Y.-L. y LI, C.-Y. (2014). Shrimp Immune System-Special Focus on Penaeidin. Journal of Marine Science and Technology, 22 (1), 1-8.
Recibido: 10-1-2018
Aceptado: 16-1-2018
Adrián Toledo. Laboratorio de Bioquímica, Departamento de Morfofisiología, Facultad de Ciencias Agropecuarias, Universidad de Camagüey, Cuba. adrian.toledo@reduc.edu.cu


{kind=link}