










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Producción Animal
versión On-line ISSN 2224-7920
Rev. prod. anim. vol.30 no.3 Camagüey set.-dic. 2018
MANEJO Y ALIMENTACIÓN
Estructura y rendimiento forrajero de Moringa oleifera cv Nicaragua en diferentes frecuencias de corte
Forage Structure and Yields of Moringa oleifera vc Nicaragua at Different Cutting Frequencies
José Leonardo Ledea Rodríguez; Giselle Rosell Alonso; Diocles Guillermo Benítez Jiménez; Ramón Crucito Arias Pérez; Yordanis Nuviola Pérez
Instituto de Investigaciones Agropecuarias "Jorge Dimitrov", Estación Experimental de Pastos y Forrajes, Bayamo, Granma, Cuba. ledea1017@gmail.com
RESUMEN
Se evaluó el efecto de la frecuencia de corte (45 y 60 días) en el comportamiento morfológico y productivo de Moringa oleifera Lam cv Nicaragua, en condiciones de riego y fertilización orgánica en un suelo Fluvisol poco diferenciado del Valle del Cauto, durante 2012. Se utilizó un diseño de bloques al azar con cuatro réplicas. La frecuencia de corte afectó las variables morfológicas (P ![]() ? 0,001), pero no los rendimientos de MS de hojas, tallos y total (P
? 0,001), pero no los rendimientos de MS de hojas, tallos y total (P ![]() ? 0,05). Los momentos en que se efectuaron los cortes en ambas frecuencias predispusieron gran variabilidad entre el primero (febrero-marzo) y el tercero (junio-julio), en 60 días se obtuvieron los mayores valores de altura de la planta (P
? 0,05). Los momentos en que se efectuaron los cortes en ambas frecuencias predispusieron gran variabilidad entre el primero (febrero-marzo) y el tercero (junio-julio), en 60 días se obtuvieron los mayores valores de altura de la planta (P ![]() 0,001) con respecto a los 45 días; mientras que el número, grosor y número de hojas por brotes se mostraron variables (P
0,001) con respecto a los 45 días; mientras que el número, grosor y número de hojas por brotes se mostraron variables (P ![]() 0,001) en los diferentes momentos del corte en función de características intrínsecas de la planta y no de la frecuencia de corte. La proporción de hojas se mantuvo superior (P
0,001) en los diferentes momentos del corte en función de características intrínsecas de la planta y no de la frecuencia de corte. La proporción de hojas se mantuvo superior (P ![]() 0,001) en la frecuencia de 45 días, y el rendimiento de MS
0,001) en la frecuencia de 45 días, y el rendimiento de MS ![]() de hojas y MS
de hojas y MS ![]() solo difirieron en el primer corte de 45 días, respecto al resto de los promedios de esta misma frecuencia de corte y a los de 60 días. Las frecuencias de corte y el momento en que se realiza intervienen significativamente en el comportamiento estructural y productivo de la Moringa; mientras que la frecuencia de corte de forma independiente solo modifica las variables estructurales.
solo difirieron en el primer corte de 45 días, respecto al resto de los promedios de esta misma frecuencia de corte y a los de 60 días. Las frecuencias de corte y el momento en que se realiza intervienen significativamente en el comportamiento estructural y productivo de la Moringa; mientras que la frecuencia de corte de forma independiente solo modifica las variables estructurales.
Palabras clave: productividad, arbustivas forrajeras, altura de corte.
ABSTRACT
The effect of the cutting frequency (45 and 60 days) on Moringa oleifera Lam vc Nicaragua forage morphology and yields with irrigation and organic fertilization was studied on little differentiated fluvisols, in Cauto Valley, in 2012. A randomized block design with four repetitions was applied. The cutting frequency affected the morphological variables (P ![]() 0.001), but not DM from leaves, stems, and total yields (P
0.001), but not DM from leaves, stems, and total yields (P ![]() 0.05). The cutting times for both frequencies predisposed great variability between the first (February-March) and the third (June-July) cuts. The best plant height values were achieved in 60 days (P
0.05). The cutting times for both frequencies predisposed great variability between the first (February-March) and the third (June-July) cuts. The best plant height values were achieved in 60 days (P ![]() 0.001), in comparison to the 45-day frequency, whereas the number, and thickness and number of leaves per shoots were variable (P ? 0.001) at different cutting times based on innate plant features and not the cutting frequency. Leaf proportion was higher (P
0.001), in comparison to the 45-day frequency, whereas the number, and thickness and number of leaves per shoots were variable (P ? 0.001) at different cutting times based on innate plant features and not the cutting frequency. Leaf proportion was higher (P ![]() 0.001) in the 45-day frequency; the DM yields
0.001) in the 45-day frequency; the DM yields ![]() from leaves and DM
from leaves and DM ![]() only differed in the first 45-day cut, in comparison to the other averages within the same cutting frequency and the 60-day cut. The cutting frequencies and times had a significant influence on the structural and productive behavior of Moringa, whereas the cutting frequency alone only modified the structural variables.
only differed in the first 45-day cut, in comparison to the other averages within the same cutting frequency and the 60-day cut. The cutting frequencies and times had a significant influence on the structural and productive behavior of Moringa, whereas the cutting frequency alone only modified the structural variables.
Keywords: productivity, forage shrubs, cutting height.
INTRODUCCIÓN
La Moringa oleifera es una planta arbórea que ha ganado importancia como forraje en los últimos años (Pérez, Sánchez, Armengol y Reyes, 2010). Es una planta rica en nutrientes, presenta alta producción de biomasa, resiste periodos de poca lluvia y altas temperaturas (Godino, Arias e Izquierdo, 2013). Estas potencialidades son criterios a tener en cuenta para su utilización en la alimentación animal, donde el clima condiciona de forma negativa la producción de alimentos de los recursos forrajeros.
En el orden de lo anterior, cabe mencionar que en Cuba, principalmente en la región oriental, se presentan condiciones climáticas que predisponen de manera marcada la producción de pastos en el período poco lluvioso (noviembre-abril). En esta región del país, se han utilizado las variedades Criolla y Nicaragua para la producción de semilla (Rosell et al. 2014); sin embargo, no han contemplado sus potencialidades para producir forrajes, ni el efecto de podas en diferentes edades en el comportamiento productivo, aunque Iglesias y Ortega (2016) señalaron que el forraje de esta arbustiva está apto para utilizarse entre los 35 y 45 días, con promedios de rendimientos entre las 10 t /ha/corte, pero sin tener en cuenta el efecto de la frecuencia de corte establecida en la estructura de la planta.
Con referencia a lo anterior, se debe mencionar que el tipo de clima y comportamiento de sus variables climáticas tienen una influencia directa en el condicionamiento del comportamiento agronómico y hábito de crecimiento (Verdecia, 2015); por lo que no siempre podrían mantener la estructura y productividad por cortes. Este es un efecto que ha sido muy poco valorado en la producción de arbustivas bajo cortes en el oriente de Cuba.
El objetivo del presente estudio fue evaluar el efecto de la frecuencia de corte en el comportamiento morfológico y productivo de Moringa oleifera cv Nicaragua.
MATERIALES Y MÉTODOS
Localidad, clima y suelo
El experimento abarcó desde febrero hasta octubre de 2012, y se desarrolló en la Estación Experimental de Pastos y Forrajes (EEPF) del Instituto de Investigaciones Agropecuarias (IIA) "Jorge Dimitrov", ubicada a 10 ½ km de la ciudad de Bayamo, y situada en los 20º 18´13" de latitud norte y los 76º 39´ 48" de longitud oeste.
El suelo del área experimental es del tipo Fluvisol poco diferenciado, según la nueva clasificación genética de los suelos de Cuba (Hernández, Pérez, Bosch, Rivero y Camacho, 2015). Presenta mediana fertilidad, su textura es de una arcilla ligera, con drenaje generalmente bueno y la topografía llana.
El clima de la región y donde está ubicada la estación, se clasifica como tropical húmedo (Barranco y Díaz, 1989). En el área de estudio las precipitaciones anuales promediaron 98,8 mm, con valores mínimos mensuales de 0 mm en los meses de enero y febrero. Estos se diferencian en la Fig. 1 con marcadores de relleno blanco. El valor máximo fue de 236 mm (septiembre), y durante el período evaluado, las precipitaciones acumuladas (1 185,3) representaron el 9,73 % del total de once años (1 1896 mm). En la Fig. 1 se ilustra la distribución mensual de las lluvias para el período experimental y la media histórica por meses.
Tratamiento y diseño experimental
Los tratamientos consistieron en las frecuencias de corte (45 y 60 días), y se ubicaron en un diseño de bloques al azar con cuatro réplicas.
Procedimiento experimental
Las unidades experimentales fueron parcelas de 9 x 7 m, separadas a 2 m . La densidad de siembra fue 0,5 m entre surcos y 0,25 m entre plantas. Se aplicó riego (500 mm), según las recomendaciones de Herrera (2006) y 20 t MO ha de origen bovino, con un contenido de 1,38 % de nitrógeno, y un aporte de 62,9 kg N ha . El corte se aplicó a 10 cm a partir de la base del tallo. La preparación del suelo fue de forma convencional (aradura, grada, cruce, grada, pase de rastrillo y nivelación), mediante tracción animal para reducir la posible afectación del ecosistema.
Las observaciones de la dinámica de crecimiento de las plantas se iniciaron luego del corte de establecimiento y se continuaron en función de las frecuencias de corte pre-concebidas. Se seleccionaron de forma aleatoria tres plantas por réplica y se les midió altura de la planta contemplada desde la base del tallo hasta el ápice de la última hoja, número y grosor de rebrotes, número de hojas/brote, peso de los tallos leñosos, peso de los tallos tiernos, peso de las hojas y rendimiento de materia seca (MS) de hojas, tallos y total.
El rendimiento de MS se estimó a partir del peso fresco de hojas, tallos y planta íntegra, para ello las plantas fueron fraccionadas en hojas y tallos y se determinó la proporción hoja/tallo en base verde. Esta proporción se extrapoló a una muestra de 200 g, y esta se introdujo en una estufa a 60º C durante 72 h, posteriormente se pesaron cuando alcanzaron valores constantes para determinar los rendimientos de MS.
Análisis estadístico
El análisis estadístico de los datos se efectuó con el software Statistica versión 10.0. Las pruebas realizadas para comprobar la normalidad y la homogeneidad de varianza fueron las de K-S y de Bartlett, respectivamente. Se utilizó un análisis de varianza y la prueba de Duncan (1955) para la comparación de medias, mediante el siguiente modelo matemático
:
RESULTADOS Y DISCUSIÓN
Como resultado del análisis para evaluar la normalidad de los datos y la homogeneidad de las varianzas de todas las variables en estudio, se obtuvieron valores P ![]() 0,06 y P
0,06 y P ![]() 0,058 para los test de K-S y Bartlett, respectivamente, aceptándose consecuentemente la hipótesis nula en todos los casos para un nivel de significación de 5 %.
0,058 para los test de K-S y Bartlett, respectivamente, aceptándose consecuentemente la hipótesis nula en todos los casos para un nivel de significación de 5 %.
Las frecuencias de corte pre-establecidas definieron en diferentes momentos el comportamiento morfológico de la variedad Nicaragua. Se afectó la altura de la planta (Tabla 1) en ambas frecuencias, pero en cada momento del corte en la frecuencia de 60 días se obtuvieron los mayores valores (P ![]() 0,001), respecto a los 45 días; sin embargo, cuando se efectuó el último corte coincidente con los meses de septiembre-octubre, el comportamiento de la altura se modificó, señalándose los valores correspondientes a la frecuencia de corte (45 días), superiores numéricamente a los obtenidos para los 60 días, pero sin alcanzar la significación.
0,001), respecto a los 45 días; sin embargo, cuando se efectuó el último corte coincidente con los meses de septiembre-octubre, el comportamiento de la altura se modificó, señalándose los valores correspondientes a la frecuencia de corte (45 días), superiores numéricamente a los obtenidos para los 60 días, pero sin alcanzar la significación.
En los tres primeros momentos del corte (febrero-marzo, marzo-abril y junio-julio) de las frecuencias en estudio (45 y 60 días), no se observaron diferencias significativas en el número de brotes; sólo el segundo corte de la frecuencia de 45 días difirió estadísticamente (P ![]() 0,001) con el tercer corte de la frecuencia de 60 días. El número de brotes en ambas edades (45 y 60 días) se deprimió en el último corte, respecto a los tres primeros (Tabla 1).
0,001) con el tercer corte de la frecuencia de 60 días. El número de brotes en ambas edades (45 y 60 días) se deprimió en el último corte, respecto a los tres primeros (Tabla 1).
Mientras que el grosor de los brotes tuvo un comportamiento más variable estimulado por el efecto de la frecuencia de corte en sus diferentes momentos, el valor significativamente superior se obtuvo en el segundo momento de corte en la frecuencia de 60 días, similar con el anterior y el posterior dentro de esta frecuencia. Tampoco difirió (P ![]() 0,05) del último momento (septiembre-octubre) en la frecuencia de 45 días. Sin embargo, se diferenció (P
0,05) del último momento (septiembre-octubre) en la frecuencia de 45 días. Sin embargo, se diferenció (P ![]() 0,001) del resto de los valores obtenidos en los diferentes momentos del corte en la frecuencia de 45 días. El valor significativamente inferior se obtuvo en el cuarto momento del corte (septiembre-octubre) (P
0,001) del resto de los valores obtenidos en los diferentes momentos del corte en la frecuencia de 45 días. El valor significativamente inferior se obtuvo en el cuarto momento del corte (septiembre-octubre) (P ![]() 0,001) en la frecuencia de 60 días.
0,001) en la frecuencia de 60 días.
El número de hojas por brote fue evolucionando en función del número de cortes en la frecuencia de 45 días. Comportamiento inverso se observó para la frecuencia de 60; de esta forma, los valores significativamente superiores se obtuvieron en el primer momento para los 60 días y en el último para los 45 días. En los momentos intermedios se observó una transición al mejoramiento (45 días) y depresión (60 días) de la variable en estudio.
Este comportamiento pudo haber influido en la proporción de hojas y tallos, además del efecto de la frecuencia de corte y momento en que se efectuaron (Tabla 2).
La mayor proporción de hojas (P ![]() 0,001) se obtuvo con la frecuencia de 45 días en el tercer corte (junio-julio), y este difirió (P
0,001) se obtuvo con la frecuencia de 45 días en el tercer corte (junio-julio), y este difirió (P ![]() 0,001) del resto de los por cientos para ambas frecuencias en los diferentes momentos en que se efectuó el corte.
0,001) del resto de los por cientos para ambas frecuencias en los diferentes momentos en que se efectuó el corte.
Los valores más deprimidos para la proporción de hojas, se obtuvieron en el cuarto corte (septiembre-octubre) en esta misma frecuencia, y fue común con sus valores propios cuando se efectuó el segundo corte en esta misma frecuencia y con el del primer corte en la frecuencia de 60 días; mientras que la proporción de tallos se mantuvo significativamente superior para P ![]() 0,001 en la frecuencia de 60 días en los cortes 2 y 3, correspondientes al momento abril-julio.
0,001 en la frecuencia de 60 días en los cortes 2 y 3, correspondientes al momento abril-julio.
En la frecuencia de 45 días se apreciaron las proporciones más bajas de este órgano, con la menor en el tercer corte (junio-julio). Este valor difirió de todos los señalados en los diferentes momentos y con los promedios de la frecuencia de 60 días.
En el rendimiento de MS ![]() aportado por las hojas (Tabla 3) se comportó variable en el tiempo. Sólo difirieron significativamente (P
aportado por las hojas (Tabla 3) se comportó variable en el tiempo. Sólo difirieron significativamente (P ![]() 0,001) los promedios que se alcanzaron en el tercer corte (junio-julio), respecto al obtenido en el primer corte (febrero-marzo) dentro de la frecuencia de 45 días.
0,001) los promedios que se alcanzaron en el tercer corte (junio-julio), respecto al obtenido en el primer corte (febrero-marzo) dentro de la frecuencia de 45 días.
La producción de materia seca a partir de los tallos se incrementó notablemente en el tercer corte de la edad de 60 días, y se diferenció significativamente (P ![]() 0,001) a la aportada por la frecuencia de 45 días en el segundo y tercer corte (marzo-julio), en el resto de los momentos del corte, los promedios entre las edades de 45 y 60 días no presentaron diferencias significativas (P
0,001) a la aportada por la frecuencia de 45 días en el segundo y tercer corte (marzo-julio), en el resto de los momentos del corte, los promedios entre las edades de 45 y 60 días no presentaron diferencias significativas (P ![]() 0,05) (Tabla 3). Para el rendimiento de MS total, los mayores promedios se obtuvieron para las frecuencias en evaluación en el tercer corte, y solo difirieron para P
0,05) (Tabla 3). Para el rendimiento de MS total, los mayores promedios se obtuvieron para las frecuencias en evaluación en el tercer corte, y solo difirieron para P ![]() 0,05 con el promedio obtenido en el primer corte con la frecuencia de 45 días.
0,05 con el promedio obtenido en el primer corte con la frecuencia de 45 días.
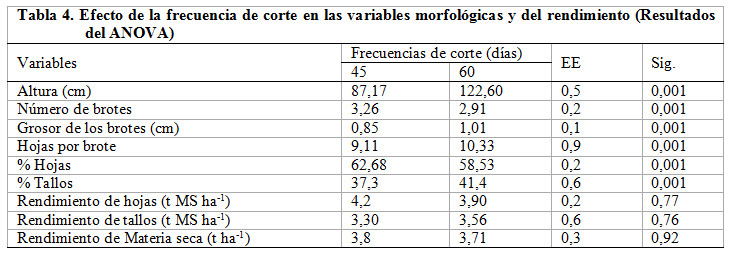
Las variables morfológicas se modificaron para P ![]() 0,001 por la frecuencia de corte; sin embargo, los rendimientos (hoja, tallo y total) no mostraron diferencias significativas (P
0,001 por la frecuencia de corte; sin embargo, los rendimientos (hoja, tallo y total) no mostraron diferencias significativas (P ![]() 0,05) (Tabla 4).
0,05) (Tabla 4).
Las frecuencias de corte y el momento del año en que coincidieron le determinaron fluctuaciones a la altura de la planta (Tabla 1), según Godino, Arias e Izquierdo (2013) para que la planta alcance un desarrollo favorable el rango superior de temperatura debe oscilar entre 25 y 35º C, y en el Valle del Cauto las temperaturas se encuentran en el rango mencionado aunque la humedad relativa supera y se mantiene cercana al 80 % (ONEI, 2015). Godino, Arias e Izquierdo (2013) señalaron que humedades relativas de forma sostenida por encima del 85 % afectan el proceso de crecimiento y desarrollo; lo que demuestra la plasticidad del ecotipo en estudio a las condiciones climáticas del Valle del Cauto, pues solo a partir del segundo tercio del período de evaluación la disponibilidad de agua no fue una limitante (Fig. 1), y ha sido identificada esta última como la principal variable climática que determina el comportamiento productivo de la Moringa.
Alfaro y Martínez (2008) registraron alturas superiores a las referidas en el presente estudio (1,20-1,50 m) a los 45 días en Guatemala, efecto que pudo estar relacionado con las condiciones climáticas del área experimental donde desarrollaron los estudios que en comparación con el de la presente investigación, existieron períodos (febrero-marzo y marzo-abril) de exposición a escasas precipitaciones (![]() 100 mm), solo auxiliadas por el riego estratégico para garantizar la persistencia del cultivo, no así para satisfacer las necesidades hídricas de las plantas.
100 mm), solo auxiliadas por el riego estratégico para garantizar la persistencia del cultivo, no así para satisfacer las necesidades hídricas de las plantas.
Obando y Ortega (2016) obtuvieron que la altura de las plantas, así como otras variables morfológicas se vieron afectadas por las densidades de siembra (1 x 0,5; 1 x 0,75 y 1 x 1 m) utilizadas en su estudio, y que esto determinó la expresión del potencial productivo. En el presente estudio, la densidad establecida (0,5 x 0,25 m) no afectó a la variable en cuestión, pues la siembra se realizó a partir de estacas. Rosell et al. (2014) determinaron que en el proceso de brotación y desarrollo, la variable que determina es el grosor de este tipo de semilla y no la densidad de siembra, al menos en estas edades tempranas.
En los períodos en que los brotes deben regenerarse, determinados por momentos del año, y que fueron establecidos por las frecuencias de corte influyen también el número de cortes; de esta forma, en las condiciones del oriente de Cuba, la variabilidad la determina la combinación de estos dos factores (Tabla 1), debido fundamentalmente al desgaste provocado por las frecuencias pre-establecidas en la acumulación de reservas en el período lluvioso (mayo-octubre), donde existe mayor exposición a horas luz y estabilidad en los niveles de irradianza (Cáceres, 1985). Parte del estudio coincidió con el último período evaluado para ambas frecuencias de corte, donde se obtuvieron los menores números de brotes (P ![]() 0,001) en ambas edades (45 y 60 días). A pesar de ser el período que más tributa a la acumulación de compuestos solubles a partir de la fotosíntesis, esta respuesta de la planta está relacionada con el número de cortes que no permitió la acumulación de reservas, y un supuesto pobre desarrollo del sistema radical.
0,001) en ambas edades (45 y 60 días). A pesar de ser el período que más tributa a la acumulación de compuestos solubles a partir de la fotosíntesis, esta respuesta de la planta está relacionada con el número de cortes que no permitió la acumulación de reservas, y un supuesto pobre desarrollo del sistema radical.
En un estudio similar Petit, Casanova y Solorio (2010) señalaron que las principales diferencias entre el número de rebrotes entre la Moringa oleifera, Leucaena leucephala y Guazuma ulmifolia manejadas en monocultivo en la estación de lluvias, radicó en las diferencias en el desarrollo radicular que alcanzaron las dos especies mencionadas, respecto a la Moringa, que siempre se mantuvo inferior de manera significativa con respecto a la cantidad de brotes en su estudio y que también fueron inferiores a los reflejados en la Tabla 1.
Sin embargo, hubo consonancia entre su número y grosor, en la medida que rebrotaron menos, los que lograron culminar este proceso fueron más robustos que el resto y se concentraron en la frecuencia de 60 días, donde la planta contó con mayor tiempo para la acumulación de compuestos orgánicos antes de ser segada.
No obstante, el número de hojas no coincidió con los momentos en los que se obtuvieron mayor número de rebrotes (P ![]() 0,001) (Tabla 1). Según Magaña (2012) esta respuesta está relacionada por el efecto del clima y no por la edad de corte, supuesto que coincide con lo obtenido en este estudio, pues en cada período en que crecieron las plantas se manifestaron condiciones climáticas similares, pero no iguales, que se manifestaron en su expresión fenotípica. El comportamiento del grosor medio de los brotes mostró variabilidad, estimulada por el efecto de la frecuencia de corte con el número de estos (Tabla 1).
0,001) (Tabla 1). Según Magaña (2012) esta respuesta está relacionada por el efecto del clima y no por la edad de corte, supuesto que coincide con lo obtenido en este estudio, pues en cada período en que crecieron las plantas se manifestaron condiciones climáticas similares, pero no iguales, que se manifestaron en su expresión fenotípica. El comportamiento del grosor medio de los brotes mostró variabilidad, estimulada por el efecto de la frecuencia de corte con el número de estos (Tabla 1).
En la revisión bibliográfica no se encontraron referencias del efecto de la poda en el comportamiento de los brotes y su grosor, solo se hace mención al efecto del pastoreo, Hrabar, Hattas y Du Toit (2009) en una poda simulada en Colophospermum mopane por elefantes, observaron una compensación entre la disminución en el tamaño de rebrotes y un aumento en el número de ellos, relacionando este proceso principalmente con la disponibilidad de agua en el suelo. En el presente estudio la disponibilidad de agua no fue una limitante; no así la edad, quién marcó las diferencias en el proceso de acumulación de compuestos orgánicos para el desarrollo de los rebrotes, pero parece existir una respuesta compensatoria en función de las condiciones climáticas en las que se encuentre la planta para el desarrollo del rebrote luego de la defoliación o sega, según sea el caso.
La proporción de hojas respecto a la de tallos se vio afectada más por el momento del corte que por la frecuencia de corte en las que se utilizaron (Tabla 2). Se observó un comportamiento variable y existió en todo momento mayor proporción de tallos que de hojas, aunque la acumulación no debe representar una preocupación para su aprovechamiento en la alimentación animal, pues los tallos presentan una calidad química (Rosell et al., 2014) y le puede permitir al animal hacer un uso eficiente de la planta íntegra.
Resulta oportuno destacar que Cordoví, Ray, Tamele, Nhantumbo y Chimbalambala (2013) señalaron valores de rendimiento expresado en base seca, superiores a los obtenidos en el presente estudio (Tabla 3) en todos los momentos en que se realizó el corte. Infiriendo de este comportamiento, que los mecanismos intrínsecos mediante los cuales las plantas regulan y expresan su fenotipo, determinaron en el ecosistema experimentaron con diferentes arbustivas, respuestas específicas para ese ecosistema, estimuladas por un amplio número de combinaciones de variables climáticas y edáficas que determinaron la respuesta productiva obtenida, y por eso las diferencias productivas con respecto al presente estudio.
Cabe agregar que los valores de altura (Tabla 4) superaron los de Fonseca, Molinet, Santiesteban y Torres (2015) cuando combinaron la edad y altura de corte sobre algunas variables morfológicas de planta de M. oleifera, en suelo fluvisol. Fonseca et al. (2015) señalaron como principal limitante para que expresara el potencial productivo, el déficit hídrico al que estuvieron mantenidas las plantas durante la fase experimental, que no eliminó las plantas, pero sí limitó su productividad; además, de un reducido desarrollo radicular por haberse encontrado en período de fomento cuando se realizó el corte de establecimiento.
En este mismo orden de ideas, para el comportamiento del número de brotes y su grosor (Tabla 4), el resultado fue el esperado. En edades tempranas las plantas movilizan las reservas para generar las primeras hojas y comenzar, a partir de la fotosíntesis, la formación de carbohidratos solubles que son utilizados para la formación y desarrollo del resto de los órganos y fracciones botánicas; es por eso que en la frecuencia de 45 días se observó mayor número de brotes, este comportamiento coincide con lo señalado por Ybalmea, Sánchez, Febles y Mora (2000) donde obtuvieron que el número de brotes se redujo con el incremento de la edad, relacionado, según los autores, a que las dominancias de los brotes más desarrollados demandan mayor cantidad de nutrientes y esto se combina con el efecto de sombra que producen en los más pequeños, provocando su muerte. Es este un criterio que pudo haber influido, pero que también es parte de la respuesta fisiológica de la planta ante el crecimiento.
Dadas las condiciones que anteceden, llamó la atención que los rendimientos de MS total que se contemplaron en el estudio no mostraran diferencias desde el punto de vista estadístico. Loyola, Pérez, Triana, Valido y Yeró (2014) cuando evaluaron la productividad de la Moringa utilizada como cerca viva con edades superiores a tres años, obtuvieron rendimiento de 2,5 t MS ha corte-1, inferiores a los referidos en la Tabla 3 para la frecuencia de 45 días; sin embargo, superaron los obtenidos en la frecuencia de 60, de esto se infiere que, más que la frecuencia de corte a la que someta la planta, el manejo al que sea sometida predispondrá la producción de MS.
Este criterio coincide con los resultados de Cordoví et al. (2013) cuando caracterizaron a varias gramíneas arbustivas en clima semiárido del sur de Mozambique en condiciones de secano, teniendo como criterio para el corte el momento de floración, y cuando este fenómeno estuvo ausente al inicio de la senescencia.
CONCLUSIONES
En las edades de 45 y 60 días las plantas mostraron desarrollos aceptables y que se correspondieron con la etapa de crecimiento de la planta, y que los momentos en los que se realizaron los cortes determinaron comportamientos morfológicos variables en las plantas y que puede definir la persistencia del cultivo y tiempo de recuperación.
RECOMENDACIONES
Se recomienda realizar el corte de las plantas de M. oleifera a los 60 días.
REFERENCIAS BIBLIOGRÁFICAS
ALFARO, N. C. y MARTÍNEZ, W. (2008). Uso potencial de la moringa (Moringa oleifera, Lam) para la producción de alimentos nutricionalmente mejorados. Guatemala: Consejo Nacional de Ciencia y Tecnología (CONCYT), Secretaría Nacional de Ciencia y Tecnología (SENACYT), Fondo Nacional de Ciencia y Tecnología (FONACYT), Instituto de Nutrición de Centro América y Panamá (INCAP).
BARRANCO, G. y DÍAZ, L. R. (1989). Clima. Nuevo Atlas Nacional de Cuba. La Habana, Cuba: Instituto de Geografía de la Academia de Ciencias de Cuba, e Instituto Cubano de Geodesia y Cartografía.
CÁCERES, O. (1985). Estudio de los principales factores que afectan el valor nutritivo de las gramíneas forrajeras tropicales en Cuba. Tesis de doctorado, Centro Universitario "Camilo Cienfuegos" , Cuba.
CORDOVÍ, E.; RAY, J. V.; TAMELE, O.; NHANTUMBO, S y CHIMBALAMBALA, A. (2013). Caracterización de especies arbóreas y arbustivas forrajeras en clima semiárido del sur de Mozambique. Rev. Pastos y Forrajes, 36(4), 434-439.
FONSECA, R.; MOLINET, D.; SANTIESTEBAN R.; ANYA, K. y TORRES, M. (2015). Efecto de combinaciones de altura y frecuencia de corte sobre el rendimiento de la Moringa (Moringa oleifera Lam) en Granma. Rev. Granma Cienc, 19(2), 1-7.
GODINO, M.; ARIAS, C. y IZQUIERDO, M. I. (2013). Interés forestal de la Moringa oleifera y posibles zonas de implantación en España. Pontevedra, España: Ed. Sociedad Española de Ciencias Forestales.
HERNÁNDEZ, A.; PÉREZ, J. M.; BOSCH, D.; RIVERO, L y CAMACHO, I. (2015). Nueva versión de la clasificación genética de los suelos de Cuba. La Habana, Cuba: Instituto de Suelos, Ministerio de la Agricultura y AGRINFOR.
HERRERA, R. S. (2006). Pastos Tropicales. Contribución a la fisiología, establecimiento, rendimiento de biomasa, producción de semillas y reciclaje de nutrientes. Mayabeque, Cuba: Ed. EDICA.
HRABAR, H.; HATTAS, D. y DU TOIT, J. T. (2009). Differential Effects of Defoliation by Mopane Caterpillars and Pruning by African Elephants on the Regrowth of Colophospermum mopane Foliage. Journal of Tropical Ecology, 25(3), 301-309.
IGLESIAS, O. y ORTEGA, M. E. (2016). Evaluación de producción primaria de la Moringa (Moringa oleifera Lamark) en la estación experimental "Mutile". Rev. Científ. Investigación y Saberes, 5(2), 84-91.
LOYOLA, O.; PÉREZ, I.; TRIANA, D.; VALIDO, A. y YERÓ, I. (2014). Evaluación agroproductiva de Moringa oleifera Lam en cercas vivas en condiciones edafoclimáticas. Rev. Prod. Anim., 26(2), 15-20.
MAGAÑA, W. (2012). Aprovechamiento pos cosecha de la moringa (Moringa oleifera). Rev. Iber. Tecnología Postcosecha, 13(1), 171-174.
OBANDO, W. I. y ORTEGA OJEDA, M. E. (2016). Evaluación de producción primaria de la moringa (Moringa oleifera Lamark) en la estación experimental "Mutile". Investigación y Saberes, 5 (2), 84-91.
ONEI (2014). Anuario estadístico de Cuba. Medio Ambiente 2014. Recuperado el 16 de febrero de 2015, de http://www.one.cu/anuarioestadisticodecuba2014medioambiente/.
PÉREZ, A.; SÁNCHEZ, TANIA; ARMENGOL NAYDA y REYES, F. (2010). Características y potencialidades de Moringa oleifera, Lamark. Una alternativa para la alimentación animal. Pastos y Forrajes, 33 (4), 33-40.
PETIT, J.; CASANOVA, F. y SOLORIO, F. (2010). Rendimiento de forraje de Leucaena leucocephala, Guazuma ulmifolia y Moringa oleifera asociadas y en monocultivo en un banco de forraje. Revista Forestal Venezolana, 54 (2), 161-167.
ROSELL, G.; BENÍTEZ, D. G.; ARIAS, R. C.; RAY, J. V.; TAMAYO, E., VERDECIA, P et al. (2014). Moringa oleifera. Experiencias en el oriente de Cuba. Granma, Cuba: Ed. Dimitrov.
ROSELL, P. A.; LEMES, BLANCA; JIMÉNEZ, A.; PEÑA, S. y MILÁN, C. (2003). Diagnóstico Urbano-Ambiental. Granma, Cuba: Ed. OPPM Bayamo.
VERDECIA, D. M. (2015). Calidad nutritiva de árboles, arbustos y leguminosas volubles, con énfasis en su contenido de metabolitos secundarios. Tesis de doctorado, Instituto de Ciencia Animal, Mayabeque, Cuba.
YBALMEA, R.; SÁNCHEZ, R.; FEBLES, G. y MORA, ENEIDA (2000). Plantación horizontal de la semilla asexual del piñón florido (Gliricidia sepium). Revista Cubana de Ciencia Agrícola, 34 (1), 73-79.
Recibido: 10-3-2018
Aceptado: 16-3-2018
José Leonardo Ledea Rodríguez, Instituto de Investigaciones Agropecuarias "Jorge Dimitrov", Estación Experimental de Pastos y Forrajes, Bayamo, Granma, Cuba. ledea1017@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}