












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
Permalink
Introducción
El selenio es considerado como un elemento traza (Duntas y Benvenga, 2015), es requerido en muy bajas concentraciones, es de vital importancia para la defensa antioxidante (Cao et al., 2015), sistema inmune (Huang, Rose y Hoffmann, 2012; Markley et al., 2017; Dalgaard, Briens, Engberg y Lauridsen, 2018; Falk et al., 2018) y la reproducción (Ahsan et al., 2014; Martins et al., 2014; Surai y Fisinin, 2015). Muchas de las proteínas que contienen Se (selenoproteínas) forman parte del sistema antioxidante, en el que participan enzimas que contienen o requieren microminerales (Chiba, 2013; Labunskyy, Hatfield y Gladyshev, 2014; Roman, Jitaru y Barbante, 2014). El selenio es un componente de la enzima glutatión peroxidasa, la cual detoxifica los peróxidos lipídicos y provee protección a las membranas celulares y sub-celulares contra el estrés oxidativo (Lubos, Loscalzo y Handy, 2011). La función antioxidante del Se ha sido demostrada que persiste en tejidos musculares incluso post-mortem, favoreciendo la conservación de la canal (Lisiak et al., 2014; Mahan et al., 2014; Calvo, Toldrá, Rodríguez, López-Bote y Rey, 2017; Jiang, Tang, Xue, Lin y Xiong, 2017). El principal cambio bioquímico ante una deficiencia de Se es la disminución en la síntesis de selenoproteínas (Seyedali y Berry, 2014) y el descenso de la actividad de la enzima GSH-Px (Oropeza-Moe, Wisløff y Bernhoft, 2015). Por lo tanto, el nivel de GSH-Px en suero es un índice confiable del estado de Se en cerdos (Adkins y Ewan, 1984). El selenio es reconocido como un nutriente esencial a un nivel de 0,15 mg/kg de alimento para cerdas y cerdos en crecimiento-finalización y 0,3 mg/kg para lechones (National Research Council, 2012). El problema que se menciona ha sido estudiado previamente (Adkins y Ewan, 1984; Zhan, Wang, Zhao, Li, y Xu, 2007). El requerimiento de selenio está basado en la concentración que éste alcanza en tejidos y no ha sido establecido en base a la actividad de la enzima GSH-Px (Jenkins y Winter, 1973; Young, Castell y Edmeades, 1977). Las fuentes de selenio inorgánico (SeI) u orgánico (SeO) adicionadas a las dietas para cerdos influyen en la cantidad de Se retenido y excretado. La retención de selenio es alta y la excreción baja cuando la fuente utilizada es SeO (Ma, Lindemann, Pierce, Unrine y Cromwell, 2014; Surai y Fisinin, 2014, 2016). Ninguna de las fuentes utilizadas favorece de manera significativa el desempeño productivo de cerdos en crecimiento-finalización (Mahan y Parrett, 1996). La actividad de GSH-Px en suero alcanza una meseta a un nivel dietario de inclusión de 0,05 (Mahan, Cline y Richert, 1999) y 0,1 ppm (Mahan y Parrett, 1996) independientemente de la fuente utilizada. Sin embargo, SeO parece tener menor biodisponibilidad para favorecer la actividad de GSH-Px medida en suero al compararlo frente al selenito de sodio (selenio inorgánico) (Mahan y Parrett, 1996). El metanálisis es un método estadístico que resumen y cuantifica el conocimiento adquirido a través del análisis de los resultados de investigaciones ya publicados (Sauvant, Schmidely, Daudin y St-Pierre, 2008). Esta herramienta permite obtener una medida del efecto combinado con una mayor precisión que la de los estudios individuales incluidos en una revisión sistemática y, por lo tanto, una mayor potencia estadística (Catalá-López y Tobías, 2014). Por lo mencionado anteriormente la suplementación dietética de Se en cerdos favorecería la actividad de GSH-Px medida en suero. El objetivo de este trabajo de investigación fue determinar el efecto de la suplementación de Se (inorgánico/orgánico) sobre la actividad de GSH-Px medida en suero sanguíneo (porción líquida de la sangre sin coágulo, ni factores de coagulación) en cerdos y el posible impacto de otros factores sobre esta variable respuesta mediante metanálisis.
MATERIALES Y MÉTODOS
Fuente de información (datos)
Se realizó una búsqueda electrónica de artículos científicos (entre enero y marzo del año 2018) en revistas indexadas con revisión doble ciego para evitar revistas predadoras basado en la metodología de Bougouin et al. (2014) en las siguientes bases electrónicas: Elsevier, Google Scholar, MEDLINE, PubMed, Science Direct, Scopus, CAB Abstract, Directory of Open Access Journals, Cambridge University Press (Fig. 1). Se utilizó una combinación de palabras clave: selenio, Se, dieta, alimento, nutrición, orgánico, inorgánico, cerdos, glutatión peroxidasa, GSH-Px en español y sus equivalentes en inglés, sin restricciones de fecha. Este trabajo de investigación no siguió los protocolos establecidos por PRISMA-P (Moher et al., 2015) pues estos han sido desarrollados para estudios en humanos. Este estudio de metanálisis sigue una metodología propia de estudios en ciencia animal, como se detalla en varios artículos publicados en los cuales se utilizó metanálisis en nutrición porcina (Apple et al., 2007; Kiefer y Sanches, 2009; Sales, 2011; Andretta et al., 2012; Létourneau-Montminy, Jondreville, Sauvant y Narcy, 2012; Remus et al., 2015; Hung, Hanson, Shurson y Urriola, 2017; Metzler-Zebeli et al., 2017; Torres-Pitarch et al., 2017; Zeng, Shurson y Urriola, 2017; Torres-Pitarch, Manzanilla, Gardiner, O’Doherty y Lawlor, 2019).

Criterios de inclusión
Se utilizó aquellos artículos en los cuales se administró selenio exclusivamente a través de la dieta y con animales sanos, el proceso de selección y descarte de artículos se aprecia en la Fig. 1. Los artículos debían incluir información respecto al número de unidades experimentales (UE) por tratamiento (repeticiones), número de animales por UE y número de animales muestreados por cada UE. Los experimentos debían incluir al menos 2 tratamientos (incluyendo el grupo control, GC), las fuentes de selenio utilizadas para la suplementación (inorgánica: selenito de sodio; orgánica: selenio-levadura) y nivel de Se, Zn, Cu, vitaminas A y E suplementados a través del alimento (nutrientes aportados a través de premezclas alimenticias comerciales, que son detallados en la fórmula de cada dieta). La actividad de GSH-Px en los trabajos seleccionados fue determinado por el método acoplado según Lawrence y Burk (1976). Respecto a la dosis o niveles de Se suplementado a través de la dieta solo se consideró aquellos valores de 0,5 ppm o inferiores puesto que valores superiores afectan de manera perjudicial el consumo de alimento en los cerdos y crea interacción con otros minerales en el alimento (National Research Council, 2012; PIC, 2016; Rostagno et al., 2017), además de ser poco usados de manera práctica por la industria. Los estudios debían haberse realizado en cerdos en etapas no reproductivas (lechones, crecimiento, finalización). Además, los artículos seleccionados debían incluir los valores de la media (promedio) y alguna medida de variación, desviación estándar (SD), error estándar (SE) de la variable en estudio para poder realizar los cálculos correspondientes caso contrario eran descartados. En ningún momento se entró en contacto con los autores de los artículos usados para la elaboración de este manuscrito.
Análisis estadístico
Para el procesamiento estadístico de los datos se utilizó MIX 2.0 Pro en Microsoft Excel (Bax, 2016). Se determinó el tamaño del efecto (TE) de la suplementación de Se sobre la actividad de GSH-Px (unidades.mL-1) por diferencia de medias (MD), con intervalos de confianza (IC) al 95 %. La heterogeneidad se evaluó por medio del índice de inconsistencia (I2) (Higgins y Thompson, 2002). El sesgo de publicación se evaluó mediante el gráfico de embudo (Funnel plot) y la prueba de regresión de Egger (Egger et al.,1997). Se utilizó un modelo de efectos aleatorios según las recomendaciones de Borenstein et al. (2011) y Sauvant et al. (2008). Se ejecutaron 8 meta-análisis que incluyó un total de 66 registros de comparación y 1 624 animales: (Mahan y Parrett, 1996; Marin-Guzman, Mahan, Chung, Pate y Pope, 1997; Mahan et al., 1999; Mahan y Peters, 2004;). Las etapas productivas fueron 2: (1) lechones y (2) cerdos en crecimiento-finalización. La fuente de selenio fue dividida en 2 categorías: (1) inorgánica y (2) orgánica, no se incluyó tipo de selenio en este trabajo pues luego de la selección de los artículos se encontró que dentro de la fuente inorgánica únicamente se había empleado selenito de sodio y dentro de la fuente orgánica se había utilizado solo selenio-levadura. Los cerdos utilizados en los trabajos individuales fueron híbridos Yorkshire-Landrace y Duroc x Yorkshire-Landrace. Para tratar de explicar la heterogeneidad entre los estudios se realizaron metaregresiones utilizando como covariables: número de UE por tratamiento (repeticiones), número de animales por UE, número de animales muestreados por UE, nivel de suplementación de Se, Cu, Zn, vitaminas A y E.
Resultados y discusión
La suplementación dietética de Se incrementa de manera significativa la actividad de GSH-Px en todos los metanálisis realizados Tabla 1: general-general (MD= 0,326; IC= 0,282 a 0,371; P <0,00001), general-inorgánica (MD= 0,327; IC= 0,269 a 0,385; P <0,00001), general-orgánica (MD= 0,325; IC= 0,255 a 0,396; P <0,00001), lechones general (MD= 0,261; IC= 0,234 a 0,288; P <0,00001), lechones-inorgánico (MD= 0,261; IC= 0,234 a 0,288; P <0,00001). En la etapa productiva de crecimiento-finalización (CF) también se encontró diferencia significativa en los metanálisis realizados general (MD = 0,328; IC = 0,281 a 0,375; P < 0,00001), inorgánico (MD = 0,331; IC = 0,265 a 0,396; P < 0,00001) y orgánico (MD = 0,325; IC = 0,255 a 0,396; P < 0,00001). En el análisis por subgrupos a través de la prueba de z se encontró que no existe diferencia entre la suplementación de SeI versus SeO (MD= 0,00185; IC= -0,126 a 0,130; P= 0,997) sobre la actividad de GSH-Px. En cuanto a la etapa productiva se determinó que no hay diferencia en la actividad de GSH-Px tras la suplementación de Se en lechones (L) frente a cerdos en CF (MD= 0,0666; IC= -0,007 a 0,140; P=0.07857). Adicionalmente, se presenta en la Tabla 1 un resumen de la actividad de GSH-Px en valores absolutos entre los grupos tratamiento y control. El efecto de la suplementación de Se sobre la actividad de GSH-Px no se presentó de manera consistente entre los estudios como lo reflejan los valores altos de heterogeneidad: general-general (I2= 96,06 %; IC= 95,48 a 96,57 %), general-inorgánica (I2= 95,71 %; IC= 94,77 a 96,48 %), general-orgánica (I2= 96,48 %; IC= 95,73 a 97,09 %). En la etapa productiva L al encontrarse un número reducido de estudios el resultado del I2 fue de 0 %. Respecto a la etapa de CF se encontró valores altos de heterogeneidad general (I2= 96,16 %; IC= 95,59 a 96,65 %), inorgánico (I2= 95,91 %; IC= 95 a 96,66 %) y orgánico (I2= 96,48 %; IC= 95,73 a 97,09 %). El análisis de metaregresión determinó que la actividad de GSH-Px se ve afectada de manera significativa por factores relacionados al diseño experimental y contenido nutricional de las dietas utilizadas en los estudios individuales como se muestra en las Fig. 2, Fig. 3, Fig. 4. y Fig. 5. A través del gráfico de embudo (Fig. 6) y de la prueba de regresión de Egger (Fig. 7) se sospecha de la presencia de sesgo de publicación. El gráfico de embudo muestra que la gran mayoría de estudios se concentran al lado derecho del valor 0 de la diferencia de medias (MD), mostrando que habría una tendencia hacia la publicación de estudios con resultados positivos, de esta manera no se grafica el embudo invertido indicador de la ausencia de sesgo de publicación, lo cual fue posteriormente confirmado mediante la prueba de regresión de Egger (Intercepto=3,74, P=0,03; Pendiente=0,17, P=0,005).

Fig.2 Metaregresión análisis general: actividad GSH-Px en suero sanguíneo y número de repeticiones Coeficiente de regresión= -0,009 (P < 0,001)
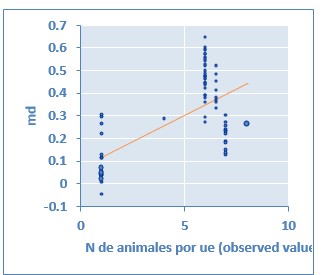
Fig. 3 Metaregresión análisis general: actividad GSH-Px en suero sanguíneo y número de animales por unidad experimental Coeficiente de regresión= 0,047 (P < 0,001)
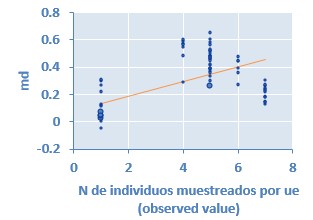
Fig. 4. Metaregresión análisis general: actividad GSH-Px en suero sanguíneo y número de individuos muestreados por unidad experimental Coeficiente de regresión= 0,053 (P < 0,001)

Fig. 5 Metaregresión análisis general: actividad GSH-Px en suero sanguíneo y nivel de selenio en la dieta Coeficiente de regresión= -0,1 (P = 0,002)
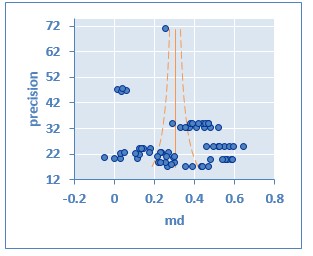
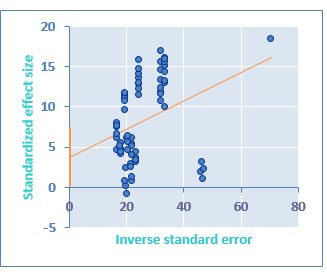
Tabla 1 Actividad enzimática GSH-Px (valores absolutos) y tamaño de efecto de la enzima GSH-Px en suero sanguíneo en cerdos
| Metanálisis | Resumen variable respuesta y tamaño de efecto (Unidades. mL-1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Tratamiento | Control | Tamaño de efecto | P | ||||||
| Media | SD | Media | SD | MD | IC | ||||
| General | General | 0,823 | 0,22 | 0,496 | 0,178 | 0,326 | 0,282 | 0,371 | <0,00001 |
| Inorgánico | 0,812 | 0,23 | 0,484 | 0,19 | 0,327 | 0,269 | 0,385 | <0,00001 | |
| Orgánico | 0,834 | 0,22 | 0,508 | 0,168 | 0,325 | 0,255 | 0,396 | <0,00001 | |
| Lechones | General | 0,75 | 0,08 | 0,1 | 0,056 | 0,261 | 0,234 | 0,288 | <0,00001 |
| Inorgánico | 0,75 | 0,08 | 0,1 | 0,056 | 0,261 | 0,234 | 0,288 | <0,00001 | |
| Crecimiento-finalización | General | 0,837 | 0,21 | 0,508 | 0,166 | 0,328 | 0,281 | 0,375 | <0,00001 |
| Inorgánico | 0,839 | 0,2 | 0,508 | 0,168 | 0,331 | 0,265 | 0,396 | <0,00001 | |
| Orgánico | 0,834 | 0,22 | 0,508 | 0,168 | 0,325 | 0,255 | 0,396 | <0,00001 | |
En el presente trabajo de metanálisis se aprecia claramente que la suplementación de Se favorece de manera significativa el TE de GSH-Px en todos los análisis realizados frente a los GC. La actividad enzimática fue mayor cuando la fuente suplementada fue SeI similar a lo encontrado en pollos de engorde en donde el selenito de sodio presenta mayor biodisponibilidad para la producción y actividad de GSH-Px (Almad et al., 2012) al compararlo frente al SeO, favoreciendo de esta manera la capacidad de defensa antioxidante del animal a un menor costo final del alimento ya que el SeI es más barato al compararlo frente al SeO. Existe una tendencia del selenito de sodio a mejorar la actividad de GSH-Px y se aprecia que si bien el SeO se retiene en mayor medida en los tejidos corporales este no favorece en la misma medida la actividad de GSH-Px. Sin embargo, el análisis de metaregresión demuestra que el nivel (dosis) de Se en la dieta (Fig. 5) afecta de manera significativa la actividad enzimática (Coef. Regresión= -0,1; P=0,002). Al realizarse el análisis por subgrupos (fuente de Se), se encontró que este efecto se debe al SeI (Coef. Regresión= -0,17; P < 0,001), no así con el SeO (0,09; P = 0,123). En un metanálisis realizado en pollos de engorde se encontró que la actividad de GSH-Px no está relacionada con la concentración de Se suplementado en la dieta (Zoidis, Demiris, Kominakis y Pappas, 2014).
Es importante considerar que el nivel de suplementación de Se a través del alimento para cerdos oscila entre 0,15 a 0,3 ppm según el National Research Council (2012). No existió diferencia estadística (P < 0,05) entre la suplementación de SeI versus SeO sobre la actividad de GSH-Px en cerdos, tampoco la etapa productiva determinó diferencia (L frente a CF) lo cual desde el punto de vista económico es importante pues el costo del SeI es inferior al SeO formulándose dietas más baratas. Con lo anterior se podría suponer que la suplementación de SeI es suficiente para cerdos en etapas de recría (lechones destetados), crecimiento y finalización. Sin embargo, se debe prestar atención a cerdos que se encuentran en etapa reproductiva (gestación, lactancia y verracos) puesto que en ellos se requiere construir reservas tisulares de Se debido a que son estados fisiológicos que demandan mayor necesidad de este mineral, obteniéndose buenos resultados con el uso del SeO (Surai y Fisinin, 2015, 2016).
En un estudio realizado por Aaron y Hays (2004) resalta la importancia de considerar el número de repeticiones, así como el número de individuos por UE antes de iniciar un trabajo de investigación ya que de esta manera es posible detectar diferencias significativas sí estas existiesen, evitando la pérdida de información valiosa. Precisamente a través de estudios de metaregresión se determinó que el número de repeticiones por tratamiento, número de individuos por UE y el número de individuos muestreados por UE afectan de manera significativa la actividad de GSH-Px en suero sanguíneo (Fig. 2, Fig. 3, Fig. 4). Se aprecia que a medida que aumenta el número de repeticiones por tratamiento la variable respuesta disminuye, mientras que cuando aumenta el número de individuos por repetición y el número de individuos muestreados por UE la variable evaluada aumenta. Los anteriores son elementos importantes a considerar al momento de realizar el diseño experimental para este tipo de trabajos, mostrando claramente que no solo se debe prever el número de repeticiones sino también el número de cerdos en cada repetición.
La capacidad de defensa antioxidante en el organismo está dada por el trabajo en conjunto realizado por enzimas y otros factores no enzimáticos como lo son las vitaminas (Halliwell, 1994). Superóxido dismutasa (SOD) es otra enzima que juega un papel importante en la cadena de reacciones químicas para controlar radicales libres o factores proxidantes, dicha enzima tiene como cofactores al Cu y Zn (Collins, 2016; Ighodaro y Akinloye, 2018). En este estudio se encontró que ambos minerales afectan de forma significativa la actividad de GSH-Px. Por cada unidad de Cu que se incremente en la dieta la actividad de GSH-Px disminuye (Coef. de Regresión= -0,076; P < 0,001), mientras que para el caso del Zn ocurre lo contrario por cada unidad de Zn que se incrementé la actividad enzimática aumenta (Coef. de Regresión= 0,0035; P < 0,001). Una situación similar ocurre para el caso de las vitaminas que tienen un rol antioxidante como lo son las vitaminas A (Chew, 1996) y E (Wang, Xu, Su, Shi y Shan, 2017). A medida que aumenta el nivel de vitamina A en la dieta aumenta la actividad de GSH-Px (Coef. de Regresión= 0,00006; P < 0,001) y lo contrario ocurre para la vitamina E, al aumentar su inclusión se reduce la actividad enzimática (Coef. de regresión= -0,011; P < 0,001). Este efecto encontrado probablemente se deba a que a mayor cantidad de vitamina E en la dieta mayor será el efecto protector sobre la membrana celular en contra de los factores proxidantes disminuyendo de este modo el trabajo para la enzima GSH-Px. En un estudio realizado en cerdas por Chen et al. (2016a, 2016b) se encontró que no existe interacción entre la vitamina E y el Se suplementados a través de la dieta sobre la actividad de GSH-Px. Sin embargo, Urso et al. (2015) en una investigación realizada en pollos encontró que vitamina E favorece la producción de GSH-Px. Es importante realizar estudios en los cuales se evalué la actividad antioxidante enzimática en cerdos empleando dietas con diversos niveles de suplementación de Cu, Zn, vitaminas A y E y determinar la existencia de posibles interacciones durante la absorción intestinal.
Respeto al sesgo de publicación encontrado (P < 0,05) se aprecia que solo se estaría dando preferencia a aquellos trabajos en los cuales se ha obtenido resultados positivos en la actividad de GSH-Px tras la suplementación de Se. Sin embargo, se debe tener presente que, si bien los animales del GC no fueron suplementados con Se, los macroingredientes de la dieta (maíz, soya, sorgo, etc.) aportan un determinado nivel de Se a la dieta basal (0,067 ± 0,027 ppm). Con lo anterior se estaría estimulando la actividad enzimática de GSH-Px. Debido a la limitación del software utilizado para este trabajo de investigación no se pudo evaluar el impacto de posibles estudios perdidos sobre el tamaño del efecto obtenido, tampoco fue posible corregir el efecto estimado, a través de la prueba de corte y relleno (trim and fill). Adicionalmente, existiría un sesgo relacionado al origen de los artículos utilizados para este trabajo pues todos pertenecen al mismo autor Mahan y colaboradores (Mahan y Parrett, 1996; Marin-Guzman, Mahan, Chung, Pate y Pope, 1997; Mahan et al., 1999; Mahan y Peters, 2004;). Sin embargo, debe considerarse que el uso de Se en cerdos es la línea de trabajo de este grupo de investigación de reconocido prestigio mundial. En trabajos futuros es importante que se evalúe el efecto de la suplementación de Se sobre la actividad de GSH-Px frente a desafíos sanitarios, puesto que esta selenoproteína es una de las armas de defensa del sistema inmune del cerdo (Dalgaard et al., 2018). Algunos autores plantean la hipótesis de que la suplementación puede potencialmente “estimular” la inmunidad celular, pues el Se puede aumentar la expresión de IL-2R (receptores interleucina 2) en las células T y mejorar las respuestas de las células T (McKenzie, Rafferty y Beckett, 1998). No existen estudios en los cuales se analice la actividad de GSH-Px y la relación con el nivel de glutatión, molécula reductora durante la reacción (Bansal y Simon, 2018).
Conclusiones
La suplementación dietética de selenio (Se) favorece la actividad de la enzima glutatión peroxidasa medida en suero sanguíneo. Existen varios factores relacionados al diseño del experimento que afectan la variable respuesta en estudio y que deben ser consideradas cuidadosamente antes de iniciar un trabajo de investigación. Otros nutrientes con función antioxidante como Cu, Zn, vitaminas A y E también están relacionadas con la actividad de GSH-Px por lo que es necesario mejorar el control de la calidad de las premezclas y alimentos concentrados por fábricas productoras de pienso

