










 Servicios personalizados
Servicios personalizados
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
La interacción genotipo-ambiente (IGA) es una de las complicaciones que se puede presentar en los procesos de selección de los animales y tiene implicaciones que puede tener importancia en este proceso. La IGA implica que existe un cambio de orden de mérito en diferentes condiciones ambientales; en otras palabras, los sementales o genotipos que se consideren como los mejores pueden no serlo bajo otras condiciones y esta situación puede ser importante desde el punto de vista económico y práctico. Menéndez-Buxadera y Mandonnet (2006) mostraron evidencias en ganado bovino lechero en diferentes países. Suárez, Zubizarreta y Pérez (2009) en sus resultados encontraron IGA en el ganado cubano Siboney de Cuba para producción de leche e intervalo entre partos considerando dos niveles de manejo y más recientemente Rodríguez y Guerra (2013) reportan IGA en ganado Cebú cubano. Igualmente, de Souza et al. (2003) encontraron IGA en rebaños de la raza Nellore en tres regiones en Brasil.
Con base en lo anterior, el objetivo del presente trabajo fue evaluar la posible existencia de IGA en ganado Santa Gertrudis en Cuba mediante la estimación de los parámetros genéticos evaluados en pruebas de comportamiento bajo condiciones de pastoreo considerando dos niveles de producción.
Materiales y métodos
Se utilizó la información referente a 4 529 animales machos y hembras que incluyeron 3 133 pedigrís hijos de 161 sementales que nacieron entre los años 1981 y 2016 de tres empresas genéticas y que tenían registrado el peso a 18 meses de edad y este se ajustó a 540 días.
Los animales incluidos en las pruebas de comportamiento, fueron destetados entre los 6 y 7 meses de edad y seleccionados para la prueba en condiciones de pastoreo, fundamentalmente a base de pastos de Cynodon nlemfuensis y Panicum máximum, y se les proporcionó sales minerales ad libitum, durante un período aproximado de 12 meses. Los animales pertenecieron a las empresas: Turiguanó, Camilo Cienfuegos y Vallina.
La información se dividió en dos conjuntos de datos, aquellos que se encontraban por encima de la media para el carácter estudiado y los que estaban por abajo, que se denominaron peso ajustado a 540 días alto (PAA) y peso ajustado a 540 días bajo (PAB) y se consideraron como dos caracteres distintos para contrastar solamente los valores extremos. Un procedimiento similar fue utilizado por Rodríguez y Guerra (2013) en ganado Cebú, pero utilizando la ganancia. Se depuró la base de datos eliminando aquellos sementales con menos de 10 hijos y grupos contemporáneos menores de tres. La Tabla 1 muestra una descripción de cada subconjunto de datos con la información fundamental.
Tabla 1 Composición genética para los dos ambientes
| Ambiente | n | No. GC | No. padres | No. madres |
|---|---|---|---|---|
| Alto | 1 489 | 134 | 132 | 1 258 |
| Bajo | 2 002 | 144 | 161 | 1 664 |
El peso se ajustó a los 18 meses de edad (540 días) de conformidad con la siguiente fórmula(Fig. 1)
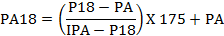
Donde:
PA=peso registrado al año
P18=peso registrado a los 18 meses
IPA-P18=intervalo en días desde la fecha en que se registró el peso al año hasta la fecha que se registra el peso a los 18 meses
Se utilizó un modelo unicarácter para cada uno de los dos caracteres (ambientes); donde se consideraron los efectos fijos de grupo de contemporáneo (rebaño-año-época de nacimiento como cuatrimestre) y sexo, y los efectos aleatorios de efecto directo del animal y el residuo. En notación matricial tenemos:
y = Xb + Za + e
Donde: y = vector de registros de peso ajustado
X y Z= matrices de incidencia
b= vector de efectos fijos (niveles de grupos contemporáneos y sexo)
a=vector de efectos aleatorios del animal
e = vector de residuos
Posteriormente se utilizó un modelo multicarácter, incluyendo ambos caracteres PAA y PAB.
En notación matricial quedaría como:
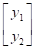 =
= 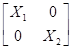
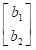 +
+ 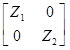
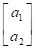 +
+ 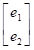
Donde:
y i = vector de las observaciones para el i-ésimo rasgo.
b i = vector de los efectos fijos para el i-ésimo rasgo.
a i = vector de los efectos aleatorios del animal para el i-ésimo rasgo.
e i = vector de los efectos residuales aleatorios para el i-ésimo rasgo.
X i y Z i son matrices de diseño que relacionan los datos con los efectos fijos y los animales, respectivamente.
En todos los casos se estimaron los componentes de varianza y covarianza, así como los parámetros genéticos, heredabilidad (h2) y correlaciones genéticas para el multicarácter. Los componentes de varianza y covarianza, los parámetros genéticos y los valores genéticos fueron estimados usando el procedimiento de máxima verosimilitud restringida (REML) y el algoritmo libre de derivadas usando el software MTDFREML (Boldman, et al. 1995).
Se calcularon las correlaciones de Pearson y Spearman entre los valores genéticos de los padres evaluados en ambos ambientes como un posible indicador de la presencia de interacción genotipo-ambiente utilizando el procedimiento PROC CORR del SAS 9.4 (2013).
Resultados y discusión
En la Tabla 2 se presentan los estadígrafos generales para PAA y PAB. Hubo mayor variabilidad para PAB y las medias generales para ambos grupos de caracteres fueron para PAA 390,99 ± 64,68 y 286,15 ± 53,05 kg considerando machos y hembras. En PAA los machos pesaron 74 % más que las hembras y en PAB 66 %, indicadores lógicos del mayor potencial de crecimiento de los machos y se amplían las diferencias a medida que el nivel de “manejo” fue superior.
Tabla 2 Estadígrafos generales para los dos ambientes
| Ambiente | n | Media (kg) | ± DE | CV (%) | Min (kg) | Max (kg) |
|---|---|---|---|---|---|---|
| Alto | 1 489 | 390,99 | 64,68 | 16,54 | 219 | 633 |
| Bajo | 2 002 | 286,15 | 53,05 | 18,54 | 112 | 340 |
Aranguren-Méndez et al. (2006) en animales mestizos doble propósito de Brahman y Holstein reportaron pesos ajustados a 540 días de 283,5 ± 38,2 kg para machos y 251,7 ± 34,1 kg para hembras, inferiores a los reportados en este trabajo considerando los dos sexos. Rodríguez Calvo et al. (2017) en ganado Cebú cubano encontraron en 6 hatos un rango de peso a los 18 meses que se movió entre 255-327 kg por debajo de los valores medios para el Santa Gertrudis. De Souza et al. (2018) con ganado Nellore en Brasil para demostrar la existencia o no de la interacción genotipo-ambiente reportan pesos ajustados a los 550 días en tres regiones de Brasil que oscilaron entre 301,1 ± 51,3 y 315,3 ± 53,2 kg que en comparación con el nivel considerado alto en nuestro trabajo están por debajo, aunque los coeficientes de variación se movieron alrededor de 17 % muy similares a los nuestros. Diferencias en el comportamiento de estos indicadores bajo diferentes sistemas de producción en la propia raza Nellore también fueron encontrados por Souza et al. (2003) y Conceição et al. (2005). Guillén Trujillo et al. (2012) con ganado Cebú Bermejo en Cuba reportaron pesos finales en pruebas de comportamiento en varias provincias del país de 327,7 ± 52,3 kg que resultó inferior al reportado en Brahman de Sudáfrica (360,8 kg) por Pico (2004). Sin embargo, fue superior a los valores mencionados en Brahman blanco de Venezuela por Plasse et al. (2002) de 297,4 kg y de Cuba por Espinoza-Villavicencio et al. (2008) con 326,5 kg.
En razas criollas en Colombia en rebaños en conservación como parte del programa de recursos genéticos animales, en un rebaño Romosinuano Ossa, Suárez y Pérez (2007) reportaron para peso a los 16 meses, 247,2 ± 33,8 kg y Martínez et al. (2009) en animales criollos Sanmartinero reportaron valores medios de 222,58 ± 39,6 kg, todos inferiores a los resultados en PAB, pero con menor edad.
En la Tabla 3 se presentan los componentes de varianza y las heredabilidades para los análisis univariados.
Posiblemente la mayor variabilidad encontrada en el nivel bajo incidió en los mayores componentes de varianza, y precisamente esa mayor variabilidad fenotípica y ambiental determinó un estimado de h2 sensiblemente menor que en el nivel alto.
Tabla 3 Componentes de varianza y heredabilidades (± EE) para PAA y PAB
| Estimados | PAA | PAB |
|---|---|---|
| σ2 A | 440 | 32 019 |
| σ2 E | 755 | 196 688 |
| σ2 P | 1 196 | 228 707 |
| h2 ± EE | 0,37 ± 0,09 | 0,14 ± 0,04 |
En análisis unicarácter, Rodríguez y Guerra (2013) estudiando el peso final en pruebas de comportamiento en ganado Cebú cubano, encontraron heredabilidades de 0,22 ± 0,04 en nivel alto y 0,15 ± 0,03 en nivel bajo. La misma tendencia que la encontrada en este trabajo, aunque la diferencia entre los extremos fue menor. Los autores plantearon la existencia de interacción genotipo-ambiente. También en Cuba y en ganado Cebú, Espinoza-Villavicencio et al. (2008) y Guillén Trujillo et al. (2012) en análisis univariados reportaron heredabilidad de 0,28, más semejantes a los encontrados en el nivel alto en Santa Gertrudis y, Rodríguez Calvo et al. (2017) en modelos multicarácter reportaron heredabilidad de 0,33 ± 0,07 para peso a los 18 meses.
En determinaciones unicarácter Aranguren-Méndez et al. (2006) en bovinos mestizos doble propósito en Venezuela reportaron un valor similar al nuestro en el nivel alto (0,38 ± 0,10). Se puede considerar que esta población era heterogénea desde el punto de vista genético y eso influye en la variabilidad genética que fue superior a los reportados en Brahman venezolano (Plasse et al. 2002) o para ganado Tabapuá (Ferraz et al. 2004) pero similar al Hereford (Meyer et al. 1993). Para Nellore en Brasil, Barbosa et al. (2017) considerando el peso ajustado a 550 días reportaron un valor de h2 = 0,26 que es intermedio a los valores nuestros.
En ganado criollo colombiano Romosinuano, Martínez, Pérez y Herazo (2006) y Ossa, Suárez y Pérez (2007) considerando el peso a 16 meses de edad reportaron heredabilidades de 0,33 ± 0,07 y 0,13 ± 0,05 con la misma población; mientras Martínez et al. (2009) con ganado Sammartinero en los llanos orientales de Colombia reportaron un valor de h2 = 0,18.
En la Tabla 4 aparecen los resultados obtenidos en el análisis bicarácter considerando los dos conjuntos de datos como rasgos independientes según lo planteado por Falconer y Mackay (2006) como una vía para determinar posible interacción genotipo-ambiente.
Tabla 4 Componentes de varianza y covarianza, parámetros genéticos y sus errores estándar para PAA y PAB
| Estimados | PAA | PAB |
|---|---|---|
| σ2 A | 321 | 136 |
| σa (AB) | 171 | |
| σ2 E | 1117 | 1098 |
| σe (AB) | -99,6 | |
| σ2 P | 1439 | 1233 |
| σp (AB) | -26,6 | |
| h2 ± EE | 0,22± 0,09 | 0,11± 0,04 |
| rg | 0,82 | |
| re | -0,09 | |
| rp | -0,02 |
Los estimados de heredabilidad fueron más bajos que los obtenidos para las estimaciones unicarácter y los componentes de varianza consecuentemente fueron más bajos. La correlación genética fue alta y positiva, pero la correlación residual y la fenotípica, aunque de signo negativo fueron prácticamente cero.
En análisis multicarácter y en trabajos tratando de encontrar evidencias de una posible interacción genotipo-ambiente, Menéndez-Buxadera et al. (2006) en Cebú de Cuba reportó heredabilidades entre 0,21-0,34. Guillén Trujillo et al. (2012) analizando el peso final en pruebas de comportamiento en ganado Cebú en Cuba reportó heredabilidad de 0,28 ± 0,10 planteando que las estimaciones multivariadas son mayores que las univariadas y que aquellas son más útiles cuando la diferencia entre las correlaciones genéticas y residuales es mayor a 0,50, lo que fue planteado por Bennett y Gregory (1996). En nuestros resultados los estimados bivariados fueron menores que los univariados y se cumple lo planteado en las diferencias de las correlaciones para validar en alguna medida estos resultados.
Verde (2010) en ganado Romosinuano en Venezuela analizando entre otros caracteres el peso a 548 días encontró interacción genotipo-ambiente desde el punto de vista estadístico.
Más recientemente, de Souza et al. (2018) trabajando con la raza Nellore en tres localidades brasileñas y analizando el peso a 550 días de edad encontraron valores altos de heredabilidad en un rango entre 0,60-0,62, confirmando los resultados de Santos et al. (2012) en el norte de Brasil y reportaron existencia de interacción genotipo-ambiente. De Lira et al. (2013) en la misma raza en tres estados brasileños encontraron heredabilidades de 0,80 en dos de ellos y 0,24 en la región de Mato Grosso que es similar al encontrado en el nivel alto en Santa Gertrudis.
Souza et al. (2003) plantearon que correlaciones genéticas menores a 0,80 en ambientes distintos para el mismo carácter es indicador de interacción genotipo-ambiente, lo que fue ratificado por Nepomuceno et al. (2013). Aunque ese es un criterio importante no se puede considerar el único y hay que valorar también el cambio en el orden de mérito entre los sementales atendiendo a sus valores genéticos. En nuestro trabajo, el hecho de que la correlación genética fuese 0,82 indica que ambos caracteres están determinados por los mismos genes que actúan aditivamente, pero necesariamente no se puede asegurar que no exista interacción genotipo-ambiente, la que se puede valorar mejor a partir de las correlaciones de Pearson y Spearman atendiendo a los valores genéticos encontrados para ambas situaciones ambientales para aquellos sementales con hijos representados en ambos ambientes.
En la Tabla 5 se presentan los resultados de las correlaciones de Pearson y de Spearman que se estimaron para los 48 toros representados en ambos ambientes y demuestran que no hay asociación entre los valores genéticos de los toros representados en los dos diferentes ambientes, es decir que los mejores en el ambiente alto, necesariamente no son los mejores bajo las otras condiciones.
Tabla 5 Correlaciones de Pearson y Spearman para los valores genéticos de los toros representados en ambos ambientes
| VGA | VGB | |
|---|---|---|
| VGA | - | 0,04 |
| VGB | 0,18 | - |
Rodríguez y Guerra (2013) reportaron en ganado Cebú en Cuba correlaciones de Pearson y de Spearman de 0,46 y 0,39, respectivamente, concluyendo que existe interacción genotipo-ambiente entre los dos ambientes extremos utilizados. Estos valores fueron superiores a los de este trabajo. También, de Souza et al. (2018) concluyeron que en correspondencia con las correlaciones de Spearman encontradas para los valores genéticos en toros de la raza Nellore en Brasil en tres estados que variaron entre 0,28 y 0,31, es suficiente evidencia de interacción genotipo-ambiente.
En la Fig. 2 aparecen para 10 toros escogidos al azar sus valores genéticos en los dos ambientes y demuestra la variabilidad y el cambio del orden de mérito de los sementales ratificando los aspectos planteados anteriormente. Por ejemplo, el 619B, 673 A, 678 A, 621B se comportaron mejor en las peores condiciones.
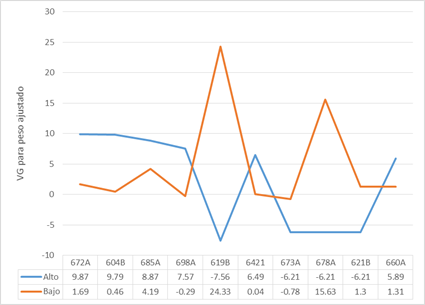
Fig. 2 Variaciones en el valor genético (VG) para una muestra de 10 toros evaluados bajo condiciones contrastantes
Ambrosini et al. (2016) plantearon que el cambio en el orden de mérito de los sementales como evidencia de interacción genotipo-ambiente debe ser especialmente considerado en ambientes desfavorables, donde es más difícil identificar los mejores animales.
Conclusiones
Las heredabilidades para peso ajustado a 540 días de edad fueron moderadas y siempre superiores para las mejores condiciones independientemente de estimarse por un modelo unicarácter o bicarácter, resultando en este último caso inferiores.
Se demuestra que bajo esta metodología ambientes contrastantes presentan evidencia de interacción genotipo-ambiente al producirse cambios importantes en el orden de mérito de los sementales

