










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La soya (Glycine max (L.) Merr.) es un cultivo de gran importancia mundial por su significativo aporte en proteínas y aceites. En los últimos años hubo un notable incremento en la producción de soya en Cuba.1 Sin embargo, las condiciones climáticas tropicales han favorecido la incidencia de enfermedades en este cultivo, que han afectado los rendimientos. La roya asiática causada por el hongo Phakopsora pachyrhizi (Sydow & Sydow) es la enfermedad foliar más destructiva de la soya, causa defoliación temprana afectando el peso y la calidad de las semillas.2 El P. pachyrhizi, podría reducir los rendimientos en más del 80 % cuando las condiciones ambientales son favorables para el desarrollo de la enfermedad.3 La presencia de P. pachyrhizi en Cuba se informó en 2009.4 Actualmente no existen cultivares de soya resistentes a esta enfermedad, los fungicidas modernos no han controlado eficazmente este patógeno y han tenido un impacto negativo en el medioambiente.
Para contrarrestar la infección por hongos, las plantas desarrollaron un sistema de inmunidad innata que ha reconocido la presencia del patógeno y comenzado una respuesta de defensa. Las defensinas son pequeños péptidos antimicrobianos que juegan un papel fundamental en esa inmunidad innata de las plantas. La defensinas pueden inhibir el crecimiento de microorganismos e insectos fitopatógenos, también pueden estar involucrados en la adaptación al estrés abiótico.5 La inhibición del crecimiento de hongos patógenos por las defensinas vegetales no está asociada a toxicidad en células de mamíferos o plantas.6 Varios genes que codifican defensinas se aislaron de plantas silvestres y se transfirieron con éxito a cultivos de importancia económica como tabaco, tomate, papa, arroz, frijol, entre otros, produciendo resistencia a diferentes patógenos.7,8 las plantas de tabaco y papa transformadas con la defensina NmDef02 aislada de Nicotiana megalosiphon mostraron resistencia a Phytophthora infestans en condiciones de invernadero y campo.7
En Cuba no se dispone de una metodología de transformación genética de soya y hasta el momento en la literatura científica no se ha informado de la existencia de plantas transgénicas de soya resistentes a hongos patógenos mediante la producción de defensinas. Considerando el potencial antifúngico de la defensina NmDef02 y la importancia de obtener cultivares resistentes a la roya asiática, este trabajo tuvo como objetivo obtener plantas transgénicas de soya que expresen el gen de defensina NmDef02 y evaluarlas frente al hongo P. pachyrhizi bajo condiciones de infección natural en campo. También se evaluó la efectividad de la nodulación por Bradyrhizobium japonicum en plantas de soya que han portado la defensina, porque la asociación con esta bacteria es fundamental para la fijación de nitrógeno atmosférico.1
MÉTODOS
Material vegetal y plásmido utilizado
Se usaron semillas de soya (Glycine max (L.) cultivar DT-84 suministradas por el INCA. Se utilizaron ejes embriogénicos de semillas maduras como explantes para su transformación por bombardeo, según la metodología establecida en el laboratorio.9 El vector pCP4EPSPS-DEF que porta los genes cp4epsps y NmDef02 se utilizó para la transformación genética.10
Transformación, selección y regeneración de plantas
Se bombardearon 150 explantes con el vector pCP4EPSPS-DEF y se seleccionaron en medio MSB5 con 20 mg/L de glifosato. Los controles DT-84 sin bombardear se cultivaron en las mismas condiciones. Los brotes regenerados se transfirieron a MSB5 sin selección para el enraizamiento.9 Las plantas se pasaron a macetas con materia orgánica y zeolita en un invernadero para obtener las semillas de la generación T1 y las siguientes generaciones. Se aplicó glifosato (360 g/L) para seleccionar plantas resistentes.
Análisis de la integración del gen NmDef02 mediante reacción en cadena de la polimerasa (PCR)
Se aisló ADN genómico total de los clones transgénicos y del control no transgénico, utilizando el protocolo CTAB.11 La mezcla de la reacción, condiciones de PCR y cebadores para el gen de la defensina se describieron previamente.10 Los productos de la reacción se chequearon en gel de agarosa 2 %.
Expresión relativa del gen NmDef02 mediante qRT-PCR
Se extrajo ARN total de 6 clones transgénicos y del control por protocolo Tri-Reagent (Sigma-Aldrich, EEUU). El ADNc se sintetizó por el kit Super-Script III (Invitrogen, EEUU). Las reacciones y condiciones para determinar la expresión relativa de la defensina mediante qRT-PCR, se describieron previamente.10
Se utilizó la β-actina de soya como gen de referencia.10 Se usó un PCR Rotor-Gene 3000 (Corbett, Sydney, Australia) y los valores se analizaron con el programa Q-Gene 96.12
Análisis por Southern blot
La transferencia del Southern y la hibridación se hizo según Sambrook et. al. (1989).13 El ADN genómico (15 μg) se extrajo por CTAB de las plantas (T5) seleccionadas con glifosato. Se digirió con enzima EcoRV y se transfirió a membrana de nylon (Hybond N, Amersham Biosciences). La hibridación se realizó con el gen cp4epsps como sonda, marcada con [32P] -dATP, utilizando el kit de marcaje de cebadores aleatorios de ADN (Promega, EEUU). La sonda se obtuvo por PCR con cebadores del gen cp4epsps y dio un fragmento de 887 pb. El producto de la reacción se purificó por el kit Gel SV Sistema Clean-Up Wizard (Promega).
Evaluación de la nodulación en plantas transgénicas de soya inoculadas con Bradyrhizobium japonicum
Se evaluó la eficiencia de la nodulación por B. japonicum (cepa Semia 5080) en plantas transgénicas de soya que portan la defensina. El procedimiento utilizado se describió previamente por Soto et al.10 Se puso control no transgénico (DT-84) y control sin inocular.
Evaluación de clones transgénicos frente a la incidencia del hongo P. pachyrhizi en condiciones de infección natural, en campo
Clones transgénicos obtenidos por autopolinización (generaciones T3, T4, T5 y T6) y el control no transgénico (DT-84) se cultivaron en parcelas del CIGB (Habana) durante los años 2013-2014, 2014-2015, 2016-2017 y en el INCA (Mayabeque) durante los años 2013-2014, 2014-2015, 2015-2016 para su evaluación frente al hongo P. pachyrhizi. Los experimentos de campo fueron autorizados por el Centro Nacional de Seguridad Biológica de Cuba con licencia: LH47-L(95)13. Las semillas inoculadas con B. japonicum se sembraron usando un diseño de bloques al azar, descrito previamente por Soto et al.10 Las hojas con síntomas de roya asiática se analizaron con el kit Envirologix QuickStixTM (Envirologix, EEUU) para confirmar presencia de P. pachyrhizi. Se calculó la severidad de la enfermedad (área de la hoja afectada por roya) según protocolo descrito.10 Se hicieron evaluaciones con 20-30 plantas.
Se usó PCR cuantitativa (qPCR) para medir biomasa de P. pachyrhizi y el procedimiento usado se describió previamente.14 Se calculó el porciento de defoliación a los 36 y 60 días. Se evaluó la afectación en los rendimientos.10
RESULTADOS
Transformación y regeneración de plantas
Después de 15 días en medio de selección con glifosato, 19 explantes bombardeados desarrollaron brotes para una frecuencia de transformación de 10,7 %. Se obtuvieron 22 clones resistentes al herbicida con una elevada eficiencia de transformación (13,5 %)9 que se transfirieron a invernadero hasta obtener semillas T2. Las plantas transformadas mostraron un crecimiento similar al control no transformado. Se demostró la expresión de la proteína CP4 EPSPS en las 22 líneas enraizadas (T0) utilizando el kit de inmunodetección Roundup Ready®.10
Integración del transgen y expresión relativa de la defensina NmDef02 en plantas transgénicas
El ADN de los clones de soya resistentes a glifosato (T1) se analizó por PCR y se detectó el fragmento esperado de 140 pb, lo que indicó la presencia del gen NmDef02 en las plantas transgénicas de soya. Se confirmó la integración estable del fragmento del plásmido que contiene los genes cp4epsps y NmDef02 (5,3 Kb) en el genoma de los clones transgénicos (T3) mediante Southern blot. El ADN de plantas no transformadas no mostró señal de hibridación.10
El análisis mediante qRT-PCR mostró que los clones transgénicos presentaron diferentes niveles de expresión relativa de la defensina NmDef02 (figura 1C). Los clones Def1, 17 y 18 mostraron diferencias significativas (p ≤ 0,001) con el control no transgénico, siendo superior en el clon Def1.
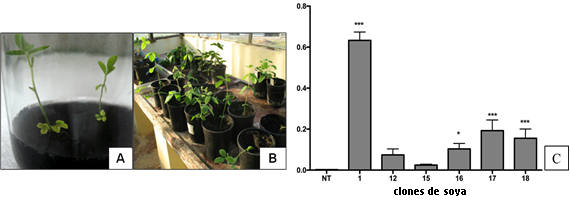
Fig. 1 Transformación de soya con el plásmido pCP4EPSPS-DEF A) Plántulas en medio de enraizamiento. B) Plantas transgénicas adaptadas a invernadero. C) Expresión relativa del gen NmDef02 en plantas transgénicas de soya. 1-18: clones transgénicos. NT: control no transgénico. Las barras representan la media (n> 9) y el error estándar. Los asteriscos indican diferencias significativas usando ANOVA por la prueba de rango múltiple de Tukey con respecto al control (*p ≤ 0,05, ***p ≤ 0,001). Los análisis moleculares mostraron la presencia del transgen en la progenie de estos clones; donde el clon Def1 acumuló los niveles más altos de expresión de la defensina y mostró el grado más alto de resistencia a ambas enfermedades.
Formación de nódulos en plantas transgénicas inoculadas con B. japonicum
Las plantas transgénicas inoculadas con B. japonicum mostraron un desarrollo fenotípico similar a las plantas no transformadas utilizadas como control. Se obtuvieron nódulos de diferentes tamaños en todas las plantas inoculadas y mayor cantidad de nódulos en las plantas transgénicas sin diferencias significativas en el peso fresco promedio de los nódulos. Los nódulos tuvieron coloración roja interna debido a la presencia de led-hemoglobina.10 Esto es un indicador indirecto del proceso de fijación de nitrógeno atmosférico por el bacteroide.1,15 Por tanto, se demuestra que la defensina NmDef02 no ha afectado la nodulación inducida por B. japonicum.
Resistencia de los clones transgénicos al hongo P. pachyrhizi en campo
La presencia del hongo P. pachyrhizi se confirmó mediante tiras de inmunodetección Envirologix QuickStixTM, en experimentos de La Habana y Mayabeque. En todos los experimentos, las plantas no transgénicas tuvieron un 100 % de afectación por roya asiática, lo que demostró la elevada susceptibilidad de este cultivar al hongo. Aunque los clones transgénicos mostraron pústulas en las hojas cercanas al suelo, tuvieron una severidad por roya significativamente menor que el control DT-84 (p < 0,05) siendo menor el número de pústulas en la zona superior de las plantas evaluadas en CIGB, en los clones Def 1, 12, 17 y 18 (figura 2A). La baja severidad por roya, también fue evidente en las plantas evaluados en el INCA donde los clones transgénicos tuvieron una severidad significativamente menor que el control Dt-84. Los resultados de los 3 años se muestran en la figura 2B. Los clones Def 1 y 17 mostraron mayor resistencia al hongo, que se mantuvo estable hasta la generación T5.

Fig. 2 Afectación por roya asiática en hojas de soya. A) Severidad por roya en la zona superior, media o inferior de plantas evaluadas en CIGB (n= 20). B) Severidad por roya en zonas superior e inferior de plantas transgénicas de soya (T3, T4 y T5) evaluadas en el INCA durante 3 años (n= 50). Diferencias significativas con el control DT-84 según prueba de Tukey.
Se demostró por qPCR que los clones transgénicos han tenido una cantidad significativamente menor de biomasa fúngica que las plantas no transgénicas (p< 0,05), siendo menor en los clones Def1, 17 y 18 (figura 3). Estos resultados han demostrado que la expresión constitutiva del gen NmDef02 produjo una disminución en el número de pústulas en las líneas transgénicas, que condujo a una menor biomasa fúngica.
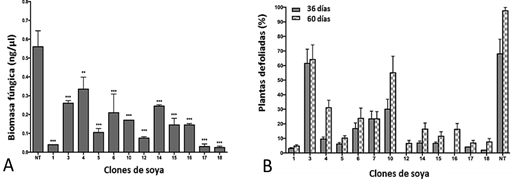
Fig. 3 Evaluación de clones transgénicos en parcela del CIGB. A) Cuantificación de biomasa de P. pachyrhizi en plantas transgénicas de soya. (n= 3). B) Defoliación en plantas de soya por incidencia de roya asiática. Evaluación a los 36 y 60 días después de la aparición de los síntomas. 1-18: clones transgénicos; NT: control no transgénico. Diferencias significativas con el control DT-84 según prueba de Tukey.
La defoliación precoz fue mayor en plantas no transgénicas a los 60 días después de la infección por roya. Contrariamente, algunos clones transgénicos mostraron una defoliación menor del 10 % en ese mismo tiempo. En este estudio, los clones transgénicos que expresaron la defensina mostraron diferentes niveles de resistencia a P. pachyrhizi, siendo mayor en los clones Def1, 12, 17 y 18. La reducida esporulación del hongo observada en algunos clones ha sido evidencia de la resistencia a P. pachyrhizi,8 en estos experimentos bajo condiciones de infección natural. Las hojas de plantas transgénicas permanecieron verdes, aunque estuvieran infestadas, contrastando con las plantas no transgénicas que mostraron clorosis intensa en sus hojas seguida de defoliación precoz (figura 4).

Fig. 4 Afectación por P. pachyrhizi en campo. A) Plantas no transgénicas (DT-84) con síntomas de roya asiática. Plantas clon Def1 resistente al hongo. B) Defoliación en plantas no transgénicas (DT-84) por maduración precoz.
La enfermedad afectó el número de ramas, vainas y semillas, así como el peso de semillas por planta en el control no transgénico. Por el contrario, la mayoría de las líneas transgénicas fueron significativamente superiores al control en todos los parámetros relacionados con el rendimiento (tabla 1).
Tabla 1 Caracterización morfo-agronómica de plantas transgénicas (Def1-Def18) y no transgénicas (DT-84) afectadas por roya asiática
| Clones | Número de ramas | Número de vainas | Número de semillas | Peso de semilla fresca/planta (g) |
|---|---|---|---|---|
| Def1 | 2,9 abc | 45,5 ab | 70,7 ab | 9,5 abcd |
| Def12 | 3,6 a | 39,2 abcde | 73,8 a | 8,1 bcde |
| Def16 | 3,5 ab | 38,0 abcde | 60,5 abc | 10,8 ab |
| Def17 | 3,4 ab | 47,2 a | 81,5 a | 11,7 a |
| Def18 | 3,1 abc | 43,5 abc | 79,2 a | 11,0 ab |
| DT-84 | 0,5 e | 22,0 f | 31,2 d | 4,7 f |
La resistencia de los clones transgénicos (Def1 y Def 17) a la roya asiática también se observó en las generaciones T5 y T6, sembradas en parcela. La presencia de pústulas se observó en todas las hojas de plantas transgénicas y no transgénicas. El control Dt-84 sufrió clorosis y defoliación precoz. Aquí se evaluó la defoliación a los 60 días, donde el control con 907 plantas sembradas tuvo un 95,2 % de plantas defoliadas totalmente. Los clones evaluados mostraron resistencia al hongo con 1,95 % de defoliación en 868 plantas sembradas (Def1) y 0,95 % de defoliación en 838 plantas sembradas (Def17). Como el hongo apareció en la etapa de llenado y maduración del grano, no hubo afectación en los rendimientos.
DISCUSIÓN
La producción de plantas transgénicas que expresan genes antimicrobianos ha proporcionado una amplia resistencia contra diferentes patógenos, al tiempo que ha reducido el uso de pesticidas químicos.
En el presente estudio, obtuvimos la primera evidencia de resistencia al hongo P. pachyrhizi en plantas de soya, transformadas con el gen de defensina NmDef02 bajo el promotor constitutivo 35S.10) Esto fue posible mediante una metodología de bombardeo de ejes embriogénicos de semillas maduras, optimizada con genotipos cubanos de soya.9
Entre los promotores constitutivos ha destacado el promotor 35S (CaMV35S) usado en este trabajo para la transformación de soya con el gen NmDef02, ha tenido un amplio uso para la expresión de genes de defensinas.16 Los promotores constitutivos han permitido la expresión en la mayoría de los tejidos de la planta y han sido útiles cuando el objetivo ha sido expresar un transgen en toda la planta, como sucedió en este trabajo. Aquí encontramos una alta correlación entre la expresión relativa de NmDef02, determinada por qRT-PCR y la alta resistencia a P. pachyrhizi en los clones Def1, 17 y 18, evaluados en parcelas de La Habana y Mayabeque, con condiciones climáticas diferentes. Aunque todos los clones transgénicos evaluados expresaron signos de roya asiática, tuvieron menor severidad en comparación con el control no transgénico (Dt-84) durante 5 generaciones.
Nuestros datos han apoyado la idea de que la expresión constitutiva del gen NmDef02 produjo una disminución en el número de pústulas en los clones transgénicos y esto condujo a una menor biomasa fúngica,10 lo que ha demostrado la resistencia de los clones al hongo, quizás por la reducción de la esporulación.17 La protección de las plantas transgénicas ha sido evidente porque las hojas han conservado su color y vitalidad, evitándose la defoliación prematura.
Se demostró que el cultivar DT-84 utilizado como control no transgénico fue altamente susceptible a este hongo y las plantas de soya sufrieron una defoliación completa antes de concluir su ciclo de maduración.10 La defoliación temprana de estas plantas redujo la productividad al interferir con sus procesos fisiológicos, lo que resultó en menos vainas normales, menos semillas por vaina y menor peso de granos.18
Conclusiones
Este ha constituido el primer reporte de transformación genética de soya con un gen de defensina para la resistencia a patógenos fúngicos. En este estudio se obtuvo la primera evidencia de resistencia al hongo P. pachyrhizi, en plantas de soya transformadas con el gen de defensina NmDef02. También se demostró que la expresión del gen de defensina NmDef02 en plantas de soya, no ha tenido un efecto negativo sobre la nodulación inducida por B. japonicum, bacteria que ha jugado un papel fundamental en la tecnología de este cultivo en Cuba. Evidentemente, el uso de plantas transgénicas que expresen esta defensina, reduciría el número de aplicaciones de fungicidas químicos en campo, para un manejo integral de patógenos en soya, con un mínimo impacto ambiental.

