










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Las predicciones de cambio climático para la zona Andina de la Región de Maule advierten para fines de siglo una disminución del 60 al 70 % en las precipitaciones anuales (Ppt) y un aumento en la temperatura de hasta 5º C, lo que sumado a la destrucción antrópica genera una condición de alta vulnerabilidad para los bosques caducifolios montanos septentrionales dominados por N. obliqua (Roble) que crecen en esta zona. (Donoso, 1982; Lara et al., 2012).
El déficit hídrico inhibe directa e indirectamente el crecimiento de los árboles (Kozlowski y Pallardi 1997, Bonan 2008, Aubry-Kientz y Moran 2017) al condicionar el balance energético que se produce entre la sobrevivencia, el crecimiento vegetativo y el reproductivo (Thabeet et al., 2009; Hulshof et al., 2012), el que sumado al impacto de la competencia inter especifica genera un problema complejo de modelar y cuantificar.
La dendrocronología es una valiosa herramienta que permite medir los impactos del ambiente sobre el crecimiento de los árboles y recuperar información de comportamientos pasados (Fritts, 1976; Babst et al., 2013). La medición histórica de crecimientos, junto a datos climáticos, permite establecer algunas deducciones básicas para evaluar el comportamiento de la especie frente a los efectos del cambio climático y realizar algunos alcances silviculturales elementales.
El prolongado déficit de precipitaciones ocurrido entre los años 2007-2012 en la zona central de Chile permite estudiar empíricamente el efecto que ha tenido sobre el Crecimiento Anual Corriente radial (CAC) de árboles, creciendo tanto en forma aislada como en competencia y constituye una aproximación de lo que podría ser una condición permanente en el futuro. El déficit prolongado de precipitaciones por seis años consecutivos constituye un escenario irrepetible para evaluar su impacto en el crecimiento de los bosques.
El propósito de este trabajo es cuantificar las relaciones existentes entre la precipitación anual, el crecimiento radial y el efecto de la competencia, así como medir la sensitividad de los anillos de crecimiento. Otro objetivo es determinar la extensión de tiempo previa al crecimiento anual que mejor explica la relación entre las precipitaciones y el crecimiento anual corriente, en un período climático normal y de sequía, en bosques secundarios de roble no manejados.
Materiales y métodos
Área de trabajo
La información fue colectada en el sector precordillerano de Monte Oscuro (19H 324862E-6113124S), localizado a 50 km al Sureste de la ciudad de Curicó, comuna de Romeral, provincia de Curicó, región del Maule, Chile. El área de estudio cubre una superficie aproximada de 1.300 hectáreas y corresponde a formaciones de bosques secundarios de los tipos forestales Roble-Hualo, Ciprés de la cordillera y Roble-Raulí-Coigüe. Esta superficie fue talada para extracción de madera aserrada y postes en la década de 1950. Posteriormente, el bosque ha sido retalado en pequeñas áreas para la producción de leña y carbón. De ahí su formación como mosaico de bosquetes coetáneos secundarios.
Desde el punto de vista morfogenético, el suelo es producto de materiales de depósitos de morrenas, acarreos fluviales y glaciofluviales, además de algunos depósitos lacustres y glaciolacustres que han sido cubiertos por capas de cenizas volcánicas (CONAF, 1999). Consecuentemente, el suelo es profundo, derivado de cenizas volcánicas y presenta una textura franco-limosa, alto contenido orgánico y baja disponibilidad de fósforo y nitrógeno (CONAF, 1999).
En cuanto a la vegetación, el área corresponde a bosques andinos de roble de altura, donde es posible encontrar la especie creciendo junto a Peumus boldus y Cryptocarya alba, propias del bosque esclerófilo, así como con Laurelia sempervirens, Aextoxicon punctatum, Nothofagus glauca y Astrocedrus chilensis, entre otras, dependiendo de la exposición en la que se encuentre (Corvalán et al., 2014) .
El clima es templado mediterráneo, según la clasificación climática de Köppen con estación seca de cuatro a cinco meses. Este se caracteriza por concentrar las precipitaciones durante los meses de invierno, mientras que en los meses de la estación seca (octubre a marzo) las precipitaciones estivales no superan los 40 mm mensuales. La zona precordillerana presenta nevazones ocasionales en invierno (Atmósfera, 2014).
En cuanto a cobertura de suelos, se contó de un conjunto de datos LiDAR con una densidad de 4.64 puntos por m2 (p*m-2) y una imagen aérea VIS con una resolución espacial de 1 m, los que fueron adquiridos y ortorectificados en marzo del 2011. A partir de esta información, se dispuso de un detallado modelo de elevación digital de terreno y de copas que permitió la medición del modelo de alturas con muy alta resolución espacial (1 m). También existen en la zona fotografías aéreas métricas del Servicio Aerofotogramétrico de Chile de 1980 y 1995-1996, escalas 1:50 000 y 1:20 000, respectivamente y fotos color de 1991, escala 1:5 000 no métricas. También se dispuso de dos intensivos inventarios forestales realizados entre 2011 y 2013.
Medición de anillos de crecimiento radial
Las muestras dendrométricas que fueron colectadas corresponden a 51 árboles que crecen en forma aislada y 172 árboles que crecen en plena competencia. Las muestras fueron extraídas en distintas condiciones de tamaño y estado de desarrollo de los rodales. La selección espacial fue realizada usando el modelo de elevación de terreno y de cubierta de suelo y las imágenes aéreas e inventarios forestales (Figura 1). Los árboles en competencia provienen de 43 parcelas de inventarios forestales. En cada parcela se identificó la especie y se midieron los diámetros normales (DAP) a los árboles mayores a 10 cm de DAP. Una vez medidos fueron ordenados en valores crecientes de DAP y de cada cuartil de la población se seleccionó el individuo central al cual se le extrajo un tarugo de crecimiento en dirección al centro de la parcela con taladro marca JIM-GEM.
Los tarugos fueron secados al aire libre, montados sobre una base de madera y lijados con tres granos diferentes para facilitar su medición. Las lecturas fueron hechas con microscopio electrónico 400X CoolingTech. Se midió el crecimiento anual corriente en radio (mm) entre los años 2001 y 2012 a nivel del DAP. Con ello se conformó una matriz de 223 filas (51 árboles aislados más 172 árboles en competencia) por 12 columnas (años 2001 al 2012). Los datos de CAC a nivel de árbol fueron promediados anualmente.
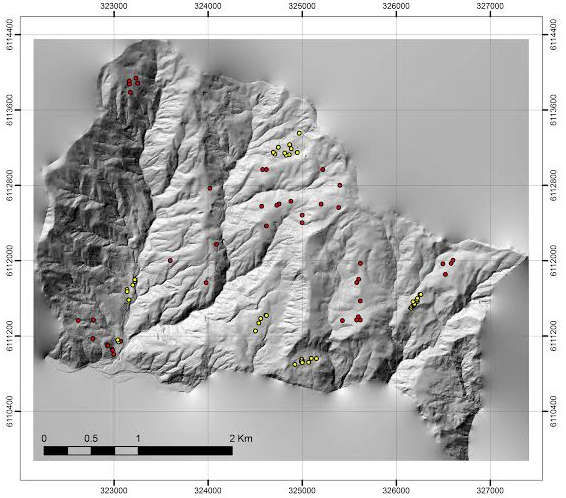
Precipitaciones anuales
Se utilizaron los datos de precipitaciones mensuales provenientes de la estación meteorológica "Monte Oscuro", perteneciente a la Dirección General de Aguas del Ministerio de Obras Públicas del Gobierno de Chile, ubicado a unos 3.5 Km del área de estudio en la misma cuenca, aguas abajo, entre los años 2001 y 2012 (DGA 2014). Los datos de precipitación anual fueron los acumulados mensualmente.
Autocorrelaciones y sensitividad
Para medir el nivel de autodependencia del CAC en DAP, se analizó la matriz de correlaciones históricas de los CAC promedios para los árboles aislados y los que crecen en competencia (Figura 2). La sensitividad se calculó como: (Ecuación 1), (Schweingruber 1988)
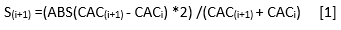
Relaciones crecimiento anual corriente vs precipitación años anteriores
Los datos de crecimiento anual corriente fueron promediados entre los años 2001-2006 (período normal) y comparados con el período 2007-2012 (período de megasequía). Se relacionó el crecimiento anual corriente con el promedio móvil de las precipitaciones ocurridas en "j" años anteriores (entre los años 1994 y 2012). Se ajustó un modelo lineal hasta encontrar el número de años que maximizara la variación del crecimiento explicado por la regresión (R2). El modelo fue el siguiente: (Ecuación 2)

Resultados y discusión
El crecimiento anual corriente en radio para el período de estudio se presenta en la tabla 1 (Tabla 1).
Tabla 1 - Precipitación, crecimiento anual corriente y sensitividad del período de análisis.
| Tipo de árboles | |||||
|---|---|---|---|---|---|
| Aislados | En competencia | Aislados | En competencia | ||
| Período | Ppt promedio (mm*año-1) | CAC promedio (mm*año-1) | Sensitividad promedio (%) | ||
| 2001-2006 | 1 630 | 3.811 | 1.812 | 0.398 | 0.351 |
| 2007-2012 | 1 062 | 3.241 | 1.407 | 0.364 | 0.384 |
| Variación (%) | 0.348 | -0.176 | -0.288 | -8.4 | 9.4 |
De la tabla 1 se deduce que una baja en la precipitación desde 1 630 mm*año-1 a 1 062 mm*año-1, (34.8 %) reduce en promedio la tasa de crecimiento desde 3.811 mm*año-1 a 3.241 mm*año-1 en los árboles que crecen aislados (17.6 %) y desde 1.812 mm*año-1 a 1.407 mm*año-1 en árboles que crecen en competencia (28.8 %).
De acuerdo a lo señalado por Donoso et al., (2010), el crecimiento radial de un bosque marginal de Nothofagus macrocarpa ubicado en Chile central es de 1.15 mm*año-1, valor muy inferior al encontrado en este estudio, tanto en árboles aislados como en competencia. Donoso (1988), estudiando renovales de roble, en un área cercana a los de este estudio, determinó crecimientos anuales radiales para períodos de 10 años entre 1.75 y 3.45 mm*año-1, valores superiores a los encontrados en este estudio en árboles en competencia. En una zona más al sur, De la Maza y Gilchrist, (1983) encontraron valores de crecimiento radiales entre 2.5 y 4 mm*año-1, superiores a de este estudio. Los resultados reportados son consistentes con el gradiente de precipitaciones que se encuentra a lo largo de la distribución latitudinal en Chile. No se reporta en la literatura un estudio semejante de la misma especie que permita comparar el impacto de la declinación de las precipitaciones, aun cotejando los datos de CAC promedio de este estudio con los señalados por Adams y Kolb, (2005), quienes al estudiar la razón de crecimiento en años húmedos versus años secos en Pinus ponderosa, Pseudotsuga menziesii, Pinus flexilis y Picea engelmannii, en el norte de Arizona reportaron valores semejantes en todos los sitios, especies y comunidades con el de este estudio (Adams y Kolb, 2005).
En la tabla 1 se presentan los valores promedios de sensitividad, dando cuenta de una disminución de la misma en árboles aislados en período de sequía y, en sentido contrario, un aumento de la misma en árboles creciendo en competencia. La disminución de la sensitividad de los árboles aislados frente a la sequía se explicaría por una disminución paulatina de las reservas de agua en el suelo profundo, haciendo cada vez más homogéneo el crecimiento anual en estas condiciones. En sentido contrario, el aumento de la sensitividad del crecimiento en árboles que crecen en competencia se explica por el efecto de la sequía entre las diferentes condiciones sociales en que crecen los individuos. La diferencia en crecimiento entre los individuos de diferentes percentiles (de DAP) se acentúa en períodos secos, haciendo más sensitivos los años más secos. Este argumento estaría indirectamente de acuerdo con lo señalado por Suárez et al., (2004), quienes estudiando Nothofagus dombeyi(coihue) en el Parque Nacional Nahuel Huapi en el noreste Patagónico, en un año de extrema sequía, encontraron que los individuos juveniles presentaron patrones de crecimiento más sensitivos y propensos a la mortalidad que los árboles más viejos (Suárez et al., 2004).
En la figura 2, se aprecia la variación de la precipitación anual y las tendencias de CAC, tanto de árboles aislados como en competencia. Se observa que existe un efecto de retardo del impacto de las precipitaciones sobre el patrón de crecimiento de los árboles, donde claramente los árboles aislados crecen más por no tener efecto de competencia. Las precipitaciones sobre el patrón de sensitividad con un efecto de retardo de entre uno y dos años de tendencias de crecimiento en ambos casos son similares. También se observa un efecto de desfase (Figura 2).
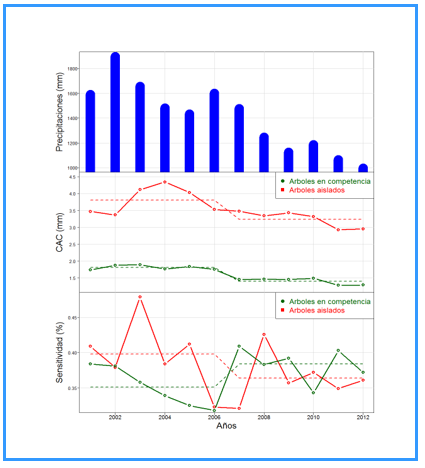
Fig. 2 - Precipitación anual (mm), crecimiento anual corriente en radio (mm/año) y sensitividad anual (%) del CAC en radio para árboles aislados y en competencia.

Fig. 3 - Matriz de correlaciones de CAC en radio entre diferentes años para el período normal de precipitaciones (2001-2006) y para el período seco (2007-2012).
Uno de los efectos más notables de la precipitación sobre el crecimiento de los árboles se observa en la figura 3, donde se muestra la estrecha autocorrelación de los crecimientos en el período estudiado. Los árboles que crecen en competencia muestran un mayor nivel de interdependencia entre crecimientos sucesivos que los árboles que crecen aislados. Este fenómeno está bien descrito por Frits, (1976), quien señala que en los árboles existe un balance energético dinámico en el crecimiento vegetativo. Las precipitaciones de un año condicionan el crecimiento de años posteriores (Fritts, 1976; Kozlowski y Pallardi 1997). El déficit hídrico inhibe directa e indirectamente el crecimiento de los árboles al condicionar el balance energético que se produce entre la sobrevivencia, el crecimiento vegetativo y el reproductivo (Thabeet et al., 2009; Hulshof et al., 2012) (Figura 3).
En la tabla 2, se muestran los parámetros de ajuste del modelo [2] de crecimiento anual corriente vs precipitación promedio en "j" años (Tabla 2).
Tabla. 2 - Crecimiento anual corriente vs precipitación promedio en "j" años anteriores.
| Tipo de árboles | Aislados | En competencia | ||||
| Número de años (j) | a | b | R2 (%) | a | b | R2 (%) |
| 1 | 3.2865 | 0.0001 | 1.3 | 1.5214 | 0.0002 | 9.5 |
| 2 | 3.0700 | 0.0003 | 4.6 | 1.1831 | 0.0004 | 30.1 |
| 3 | 2.6310 | 0.0006 | 12.9 | 0.9029 | 0.0006 | 41.7 |
| 4 | 2.1015 | 0.0013 | 30.8 | 0.9056 | 0.0007 | 34.3 |
| 5 | 1.7253 | 0.0017 | 46.4 | - | - | - |
| 6 | 1.6525 | 0.0014 | 57.3 | - | - | - |
| 7 | 1.5741 | 0.0014 | 22.0 | - | - | - |
Se puede observar que, bajo este simple modelo lineal, la precipitación promedio disminuye los errores de ajuste al incorporar mayor cantidad de años, llegando a explicar un 57.3 % de la varianza total del error en el caso de los árboles aislados, al usar una venta móvil de seis años y el 41.7 % en el caso de los árboles en competencia al usar tres años. Esta información resulta de interés si se relaciona con la matriz de autocorrelaciones señalada en la figura 3. En ambos casos, puede ser interpretado como un efecto positivo de almacenamiento de agua en el suelo. Algunos estudios que relacionan el crecimiento con la precipitación han sido especialmente desarrollados en zonas tropicales. Brienen y Zuidema (2005) encuentran un coeficiente de determinación de 7.9 % para Cedrelinga catenaeformis, una leguminosa que crece en un bosque lluvioso en Bolivia, al estimar el ancho del anillo de crecimiento estandarizado como función de la precipitación total. Devall et al., (1995) encuentran en Panamá Central coeficientes de determinación entre 28 % y de 44 % para Pseudobombax septenatum, Cordia alliodora y Annona spraguei. No se dispone de información que permita comparar el crecimiento con el efecto acumulado en el tiempo del almacenamiento de agua en el suelo.
Una disminución de 34.8 % en la precipitación promedio anual (2006-2012) (desde 1 630 a 1 062 mm*año-1) respecto del período (2001-2006) genera una caída de:
El 17.6 % de crecimiento anual corriente en radio al DAP en árboles aislados (desde 3.8 mm*año-1 a 3.2 mm*año-1)
El 28.8 % de crecimiento anual corriente en radio al DAP en árboles en competencia (desde 1.8 mm*año-1 a 1.4 mm*año-1)
La precipitación promedio anual como variable predictor del CAC, es capaz explicar que:
El 57.3 % en los árboles aislados, al promediar los últimos seis años de precipitación anual.
El 41.7 % en los árboles en competencia, al promediar los últimos tres años de precipitación anual.
Las autocorrelaciones del CAC son significativas (al 95 % de confiabilidad) y alcanzan a todo el período de análisis (12 años), tanto para los árboles aislados como para los árboles en competencia, abarcando tanto los años normales de precipitación como los años secos.
En el período seco, respecto del período normal:

