










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La caracterización anatómica y morfológica de las acículas ha sido empleada ampliamente para distinguir adaptaciones dentro de especies a condiciones ambientales particulares (Köbölkuti et al., 2017; Jankowski et al., 2017; Kivimäenpää et al., 2017) y en algunos casos como caracteres para identificar o diferenciar especies de un mismo género, similares fenotípicamente, en particular en Pinus (García-Álvarez et al., 2009; Iglesias et al., 2012; Dörken y Stützel, 2012; Meng et al., 2018).
Schoettle y Rochelle, (2000) en Pinus flexilis reportaron una baja variación morfológica en variables del crecimiento del árbol (DAP, H,) longitud, incrementos anuales y foliares) entre sitios ubicados entre 1 630 y 3 330 msnm y una marcada variación en variables anatómicas de la densidad estomática y el número de hileras de estos, asociándolo a una alta plasticidad fisiológica o una amplia tolerancia ecofisiológica. Reinhardt et al., (2011), en un estudio ecofisiológico en dos procedencias de esta especie, comprobaron un efecto significativo de la altitud sobre variables fisiológicas relacionadas con el intercambio gaseoso y la fotosíntesis y su relación con la variación en la anatomía del aparato estomático.
En especies con distribución euroasiática, P. sylvestris y P. uncinata (Boratyñska y Pashkevich, 2001; Boratyñska y Bobowicz, 2001), se encontraron variaciones anatómicas a nivel de la epidermis de la acícula que facilitaban la diferenciación entre ambas especies en hábitats simpátricos. Por su parte, Boratyñska et al., (2003) emplearon la morfología y anatomía de las acículas para comparar dos poblaciones de Pinus uliginosa y cuantificar el grado de hibridación del complejo P. mugo x P. sylvestris en la definición taxonómica de P. uliginosa, a través de variables como número de estomas y de canales resiníferos.
García-Álvarez et al., (2009) resaltan la importancia de las características de la estructura foliar en la identificación y clasificación especies del género Pinus del Mediterráneo Ibérico, a partir de láminas palinológicas con el uso de características de la cutícula y estomas.
Otros estudios muestran que variables asociadas a los canales resiníferos son factibles para la definición taxonómica en coníferas (Dörken y Stützel, 2012; Sheue et al., 2014; Ghimire et al., 2014) y entre poblaciones de una especie como rasgo adaptativo a la altitud y la exposición (Tiwari et al., 2013; Köbölkuti et al., 2017; Jankowski et al., 2017).
Ju-Kyeong y Jung-Oh, (2013)en un estudio anatómico de las acículas en 289 individuos de Abies koriana, detectaron una variación clinal en cuanto al número, tipo de canales resiníferos, ancho de la acícula y grosor de esta, lo cual estaría relacionado con la adaptación a ambientes más extremos. En estudios más recientes, Èortan et al., (2017) demuestran que los estomas son rasgos genéticamente controlados, por lo que pueden ser usados para estudios de diferenciación entre poblaciones de Populos nigra en tres valles del río Danubio. De esta manera, variables anatómicas como longitud, ancho, forma de las estomas, ancho de poros, densidad estomática, coeficiente de forma y área de las estomas mostraban diferencias significativas entre poblaciones.
Pinus tropicalis Morelet es una especie endémica del occidente cubano y constituye un relicto del linaje euroasiático en Centro América y el Caribe (Geada-López et al., 2002; Geada-López et al., 2004; Gernandt et al., 2005). La especie forma pinares puros continuos o en simpatría con Pinus caribaea Morelet var. caribaea Barret y Golfari, sobre edátopos muy extremos: arenas blancas silíceas y pizarras (López-Almiral, 1982 y Geada-López et al., 2004) reportaron diferencias morfológicas en el cono y las acículas y a nivel de ADN cloroplástico entre poblaciones que corresponden a edátopos de arenas blancas y pizarras.
De acuerdo a los aspectos antes mencionados y teniendo en cuenta que P. tropicalis habita los edátopos más extremos por sus características hídricas y nutricionales, esta investigación tiene como objetivo caracterizar la variación anatómica en acículas de la especie en tres localidades, diferenciar las poblaciones de acuerdo a las variables anatómicas y determinar la o las variables que más contribuyen a esta diferenciación. Todo esto fundamentará los planes para la selección de diferencias de fuentes semilleras que logren mejor establecimiento y productividad en función del sitio a plantar.
Materiales y métodos
Muestreo: el estudio se realizó en tres localidades correspondientes a poblaciones naturales de P. trpoicalis: Petriles, San Ubaldo y Cayo la Mula de la provincia de Pinar del Río (Tabla 1). Se colectaron acículas de la porción media de cada árbol a razón de al menos 30 árboles por localidad.
Tabla 1 Georreferenciación de las localidades de colecta de acículas de P. tropicalis.
| Localidad | Coordenadas geográficas | Edátopo | |
| Latitud | Longitud | ||
| Pretiles | 22° 23' 53'' | 84° 19' 24'' | arenas blancas |
| San Ubaldo | 22° 03' 52'' | 84° 01' 02'' | arenas blancas |
| Cayo la Mula | 22° 38' 56'' | 83° 35' 10'' | pizarra |
Variables anatómicas medidas
Se realizaron cortes transversales a tres acículas de cada árbol, se midieron las siguientes variables anatómicas: número de estomas, número de canales, grosor de la cutícula, grosor de la epidermis, grosor de la hipodermis, grosor del parénquima clorofílico, grosor del parénquima de transfusión, distancia al canal derecho, distancia al canal izquierdo. Los valores fueron promediados para los análisis estadísticos. Adicionalmente, se clasificó el tipo del canal resinífero de acuerdo a Sheue et al., (2014). Las mediciones se realizaron con un microscopio óptico modelo NLCD-307B.
Análisis estadístico
Análisis de componentes principales: se empleó con el objetivo de revelar las relaciones entre variables, a fin de reducir la dimensionalidad del análisis y explicar las relaciones con un número mínimo de dimensiones y expresar la variabilidad conjunta de todas las variables. Se consideraron los componentes cuyos autovalores fueran superiores a uno; la matriz de componentes fue rotada de acuerdo al método Varimax. Para este análisis no se tuvo en cuenta el efecto del factor localidad.
Comparación entre localidades: se empleó la prueba de Kruskal-Wallis (k muestras independientes) para establecer si existían diferencias entre las localidades para cada una de las variables anatómicas estudiadas; y la prueba de U de MannWhitney (dos muestras independientes), ambas para p = 0,05 para comprobar si había un efecto significativo del factor localidad.
Clasificación de las localidades: se empleó la técnica multivariante de análisis de discriminante con el fin de maximizar las diferencias entre localidades, consideradas estas como variables dependientes o variables de clasificación. Como variables independientes se consideraron todas las variables anatómicas. Otro objetivo de este análisis fue el de identificar la(s) variables que más contribuyen a diferenciar las localidades.
Resultados y discusión
Características anatómicas de las acículas de P. tropicalis
La figura 1 representa la estructura equifacial de un corte transversal de una acícula de P. tropicalis. En la parte externa se aprecia una cutícula definida como modificación del tejido epidérmico que es uniestratificado. Hacia el interior se ubica un tejido hipodérmico formado por varias capas de células no uniformes; seguido a estos se localiza el parénquima clorofílico muy bien definido, formando paquetes delimitados por los amplios canales. Al interior se encuentra el parénquima de trasfusión muy amplio y en el centro los haces vasculares xilema y floema (Figura 1).
Los canales resiníferos variaron en su número, pero fueron clasificados de tipo marginal (Sheue et al., 2014); estos pueden ocupar varios tejidos, desde el epidérmico hasta el parénquima clorofílico. Este carácter de tipo marginal es considerado ancestral para el género (Dörken y Stützel, 2012; Seong y Offner, 2013; Sheue et al., 2014) y concuerda con la posición basal dentro del linaje de la sección Pinus: subsección Pinus de esta especie en la filogenia molecular (Gernandt et al., 2005; Gernandt et al., 2008); Keeley, 2012).

Relación entre las estructuras anatómicas de acículas de P. tropicalis
De acuerdo a los resultados del análisis de componentes principales, las relaciones entre las variables de la anatomía foliar de P. tropicalis se pueden explicar mediante tres dimensiones, cuyos autovalores son superiores a uno y que expresan el 55 % de la variabilidad presente (Tabla 2).
Tabla 2 - Análisis de componentes principales realizado sobre la matriz de correlación de las variables anatómicas de acículas de P. tropicalis.
| Componente | ||||
| Variable anatómica | Comunalidad | 1 | 2 | 3 |
| Grosor de hipodermis | 0,710 | 0,826 | -0,149 | -0,071 |
| Número de canales | 0,687 | 0,692 | 0,242 | 0,070 |
| Grosor parénquima clorofílico | 0,623 | 0,609 | -0,295 | 0,478 |
| Distancia canal izquierda | 0,621 | 0,593 | 0,233 | -0,182 |
| Distancia canal derecha | 0,582 | 0,091 | 0,762 | -0,179 |
| Número de estomas | 0,542 | 0,053 | 0,742 | 0,169 |
| Grosor parénquima transfusión | 0,444 | 0,007 | 0,540 | 0,067 |
| Grosor de cutícula | 0,439 | -0,213 | 0,065 | 0,758 |
| Grosor de epidermis | 0,296 | 0,087 | 0,059 | 0,658 |
| Autovalor | 1,987 | 1,629 | 1,329 | |
| Varianza explicada | ||||
| % de Variación | 21,656 | 18,350 | 14,939 | |
| Acumulado % | 21,656 | 40,006 | 54,945 | |
De acuerdo al valor de las comunalidades, las variables que más contribuyeron a la segregación de los componentes fueron en orden del valor de extracción (mayor de 0,5): grosor de hipodermis, número de canales, grosor parénquima clorofílico, distancia canal izquierda, distancia canal derecha y número de estomas.
El primer componente explica la relación entre grosor de hipodermis, número de estomas y número de canales; las tres variables se correlacionan significativamente (p<0,01) y de forma positiva. La relación entre estas tres variables podría estar asociada con el ambiente hídrico en que se desarrolla la especie.
El grosor de la hipodermis, con hasta cuatro capas de células, pudiera ser resultado de la adaptación al control a la pérdida de agua. El número de estomas, aunque contradictoriamente alto para el ambiente donde se desarrolla la especie, está muy hendido, llegando en ocasiones al parénquima clorofílico. Este comportamiento ha sido reportado por García-Álvarez et al. (2009); Tiwari et al., (2013) y Köbölkuti et al., (2017) para pinos que crecen en ambientes muy secos.
El segundo componente, que explica la relación de las distancias de los canales a la epidermis, muestra la alta simetría a ambos lados para todas las localidades; esto coincide con lo descrito para muchas especies filogenéticamente emparentadas donde se describe una disposición marginal de estos canales (Seong y Offner, 2013; Sheue et al., 2014).
El grosor parénquima clorofílico, aunque contribuye a la varianza total en el análisis, es una variable que no se correlaciona significativamente con el resto, por tanto, no es útil a la hora de explicar las relaciones entre estas.
Comportamiento de las variables anatómicas por localidades
En la tabla 3, se presentan los valores promedios de las variables estudiadas por localidades, de acuerdo a la prueba de Kruskal-Wallis, comprobándose un efecto significativo del factor localidad solamente sobre la variable número de estomas. (Tabla 3)
Tabla 3 - Comportamiento promedio de las variables estudiadas por localidad.
| Localidad | Número de estomas | Número de canales | Grosor de cutícula | Grosor de epidermis | Grosor de hipodermis | Grosor parénquima clorofílico | Grosor parénquima transfusión | Distancia canal derecha | Distancia canal izquierda |
| Pretiles | 12,93 |
7,58 | 0,01 | 0,20 | 2,62 | 1,57 | 1,63 | 3,22 | 3,12 |
| San Ubaldo | 17,07 |
8,20 | 0,06 | 0,20 | 3,73 | 1,26 | 1,61 | 3,23 | 3,35 |
| Cayo La Mula | 13,73 |
6,27 | 0,02 | 0,21 | 2,00 | 1,38 | 1,61 | 3,08 | 3,13 |
Letras desiguales indican diferencias significativas p<0,05 prueba U de MannWhitney
El desarrollo y la diferenciación estomática están determinados por factores genéticos (He et al., 1998), pero la morfología y los tipos anatómicos de las acículas de pino dependen, además, de los factores abióticos (Schoettle y Rochelle, 2000); Zhang et al., 2012 y Jankowski et al., 2017).
Por esto se deduce que el mayor número promedio de estomas en las muestras de la localidad de San Ubaldo pueden estar relacionados con las características edáficas, que se caracterizan por presentar suelos extremadamente arenosos, con muy poca retención de agua, y una alta reflexión de la luz por el color claro de su superficie.
Jinxing et al., (2001) señalan que los parámetros estomáticos están afectados por factores tales como la altitud, concentración atmosférica de CO2(Loumala et al., 2005; Hartikainen et al. 2009), la temperatura, la intensidad de la luz y la reflexión lumínica (Kivimäenpää et al., 2017). Tiwari et al., (2013) y Köbölkuti et al., (2017) reportan que tanto para P. roxburguii y P. sylvestris la organización estomática tiene un importante rol en el ajuste de las plantas a diferentes altitudes, detectando que la densidad estomática se incrementa con la elevación, debido a una menor dispersión de la luz y una mayor tasa de transpiración. En cambio, Schoettle y Rochelle (2000), en la especie Pinus flexilis encuentran que en las montañas ocurre una disminución significativa en la densidad estomática con la elevación, relacionándolo con una respuesta de aclimatación para restringir la pérdida de agua a grandes altitudes.
Aunque estadísticamente no son significativos, tanto las características anatómicas de las acículas de P. tropicalis en la localidad de San Ubaldo, como el número de canales de resina, grosor de cutícula y grosor de hipodermis son superiores al resto de las localidades y el grosor del parénquima clorofílico es inferior. Estas características también pueden ser ajustes anatómicos a condiciones edáficas y microclimáticas singulares de estos ecótopos.
Ju-Kyeong y Jung-Oh. (2013) y Tiwari et al., (2013), en un estudio comparativo de acículas de Abies koreana y P.roxburguii, comprobaron que las características de los canales de resina tienen una variación clinal y altitudinal. Por otro lado, O'Neill, et al., (2002), en un estudio del efecto de la variación geográfica sobre las características de los canales de resina en plántulas de abetos, encontraron que el tamaño de los canales varió entre localidades y lo asocian a la extensión del verano, el índice de aridez y al efecto de la continentalidad.
Finalmente, P. tropicalis tiene canales marginales y su número varía en función de la disponibilidad de agua en las localidades, pero estudios más amplios son necesarios para explicar esta relación.
Clasificación de las localidades de acuerdo a las características anatómicas de las acículas
En la tabla 4, se presenta la contribución de cada una de las variables empleadas para la clasificación. (Tabla 4)
Tabla 4 - Prueba de igualdad de medias entre grupos.
| Wilks' Lambda | F | df1 | df2 | Sig. | |
| Número de estomas | 0,609 | 32,792 | 2 | 102 | 0,00 |
| Número de canales | 0,735 | 18,424 | 2 | 102 | 0,00 |
| Grosor de cutícula | 0,828 | 10,619 | 2 | 102 | 0,00 |
| Grosor de epidermis | 0,993 | 0,376 | 2 | 102 | 0,687 |
| Grosor de hipodermis | 0,305 | 116,255 | 2 | 102 | 0,00 |
| Grosor parénquima clorofílico | 0,765 | 15,685 | 2 | 102 | 0,000 |
| Grosor parénquima transfusión | 1,000 | 0,023 | 2 | 102 | 0,978 |
| Distancia canal derecha | 0,997 | 0,160 | 2 | 102 | 0,852 |
| Distancia canal izquierda | 0,994 | 0,294 | 2 | 102 | 0,746 |
Se comprobó que las variables número de estomas, número de canales, grosor de cutícula, grosor de hipodermis y grosor del parénquima clorofílico tienen promedios desiguales en las variables discriminantes, por lo que contribuyen a diferenciar las localidades. De acuerdo a la prueba Wilks' Lambda, las dos funciones obtenidas son capaces de discriminar (p<0,05) entre los grupos.
La primera función consigue explicar el máximo de diferencias entre las localidades, de acuerdo a los valores de los centroides de las funciones discriminantes, en particular entre la localidad de San Ubaldo con respecto a los otros dos grupos (Tabla 5). La variable que más contribuye a distinguir este grupo, de acuerdo a los valores estandarizados de los coeficientes de las funciones discriminantes, es el grosor de hipodermis, seguida de número de estomas y de canales. La segunda función discrimina las dos localidades restantes, que se caracterizan por presentar un menor número de estomas, menor número de canales, menor grosor de cutícula y mayor grosor de parénquima clorofílico con respecto a San Ubaldo (Tabla 6).
Por la importancia que tiene el número de canales para la discriminación, es válido señalar que se han reportado variaciones intraespecíficas en Picea mexicana(Flores-López et al., 2013), asociadas a condiciones de mayor humedad y menor aridez, y en Picea sitchensis y Picea glauca asociadas a una disminución por el efecto de la continentalidad en poblaciones al interior de su área de distribución (O'Neill et al., 2002). De igual manera, Tiwari et al., (2013) encontraron variaciones a lo largo de un gradiente latitudinal con una disminución de estas estructuras. Jankowski et al., (2017), en un estudio realizado a P. sylvestris obtuvo que la adaptación al frío trajo consigo variaciones en la anatomía, demostrándose que existe una relación entre la temperatura, el número y volumen de canales positiva.
Sin embargo, para un pino insular con pequeño areal de distribución como P. tropicalis esta es la primera vez que se encuentra marcada diferencia en la anatomía de la acícula.
Tabla 5 - Centroides de las funciones discriminantes por grupos (localidad).
| Localidad | Función | |
|---|---|---|
| 1 | 2 | |
| Pretiles | -0,892 | 0,821 |
| San Ubaldo | 3,080 | -,217 |
| Cayo La Mula | -1,742 | -1,015 |
Tabla 6 - Coeficientes estandarizados de las funciones discriminantes canónicas.
| Función | ||
| 1 | 2 | |
| Número de estomas | 0,500 | -0,704 |
| Número de canales | 0,315 | 0,518 |
| Grosor de cutícula | 0,197 | -0,387 |
| Grosor de epidermis | -0,136 | 0,070 |
| Grosor de hipodermis | 0,804 | 0,416 |
| Grosor parénquima clorofílico | -0,507 | 0,502 |
| Grosor parénquima transfusión | -0,010 | -0,049 |
| Distancia canal derecha | 0,013 | 0,212 |
| Distancia canal izquierda | 0,111 | 0,227 |
En la figura 2, se representan los centroides de cada uno de los grupos o localidades y la ubicación, de acuerdo a los valores obtenidos de las funciones discriminantes, de cada uno de los casos. Se observa una mayor agrupación en las localidades de Pretiles y Cayo la Mula, que además tienen alrededor de un 20 % de solapamiento; o sea, casos de una u otra localidad que presentan características de la otra (Tabla 7).
Esto corrobora la singularidad desde el punto de vista edáfico de la localidad de San Ubaldo, que como se señaló anteriormente deben determinar las características anatómicas que prevalecen en esta localidad (Figura 2).
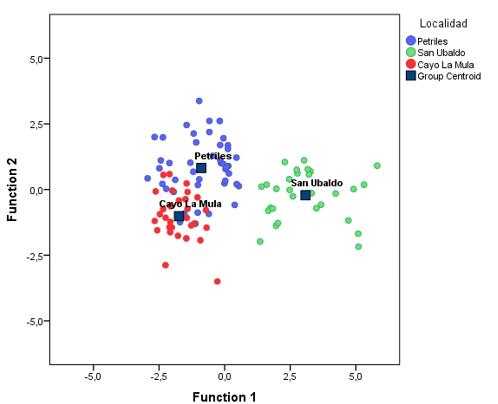
Fig. 2 - Distribución de los casos y los centroides de cada localidad en base a las dos primeras funciones discriminantes.
Las funciones discriminantes logran clasificar el 100 % de los casos de la localidad de San Ubaldo, distinguiéndola claramente del resto de las localidades. En la tabla 6, se presenta un resumen del análisis, a través del cual se logró clasificar correctamente el 86,7 % de los casos de acuerdo al grupo de pertenencia original. Por lo tanto, es posible clasificar, a partir de las variables anatómicas, las localidades; esto pudiera indicar que, en determinadas poblaciones, hubiera cierto grado de adaptación o aclimatación al ambiente.
Tabla 7 - Resultados de la clasificación.
| Localidad | Grupo de pertenencia pronosticado | Total | ||||
| Pretiles | San Ubaldo | Cayo La Mula | ||||
| Original | Pretiles | 80 | 0 | 20 | 100 | |
| San Ubaldo | 0 | 100 | 0 | 100 | ||
| Cayo La Mula | 16,7 | 0 | 83,3 | 100 | ||
Varios investigadores han reconocido que el análisis de las características morfológicas, debe ser realizado en una situación bien concebida, lo que quiere decir que las colectas deben ser a través de toda el área de distribución de la especie.
La anatomía de las acículas de P. tropicalis corresponde a una estructura equifacial semicircular con grandes canales resiníferos de tipo marginal, estomas hendidos, parénquima de transfusión y clorofílico bien definidos.
Existe una gran variación en las variables anatómicas entre las localidades y una relación significativa entre las variables grosor de hipodermis, número de estomas y número de canales, aspecto que pudiera estar relacionado con las condiciones hídricas donde se desarrolla la especie.
Las variables anatómicas número de estomas, número de canales, grosor de cutícula y grosor de hipodermis logran clasificar correctamente las tres localidades.

