












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
PermalinkRevista Cubana de Ciencias Forestales
ISSN 2310-3469
01--2020
Original article
Phenology of flora components in the use of natural and traditional medicine in the community of Verraco, Santiago de Cuba, Cuba
1Centro Oriental de Ecosistemas y Biodiversidad, Santiago de Cuba. Cuba.
Based on ethnobotanical studies carried out in the community of Verraco, belonging to the Baconao Biosphere Reserve, province of Santiago de Cuba, the components of the flora used in natural and traditional medicine were identified in situ, and the reproductive phenology of each of the identified species was determined. The objective of the work was to present the flowering and fruiting patterns of flora species that are components of natural and traditional medicine in the coastal community of Verraco, which belongs to the Baconao Biosphere Reserve. A total of 82 species of flowering plants used by the community for natural and traditional medicine were recorded, which are grouped into 75 genera and 49 botanical families. Of the total species studied, 46 % (38) were recorded with flowers and fruits, while only 12 % (10) and 39 % (32) were determined in the flowering and fruiting stages, respectively. Of the total species, only four are endemic and 24 % are naturalized, while 19 % correspond to non-indigenous but possibly naturalized species. Forty species flowers, coinciding with the rainy season, while 36 bear fruit during the same period. In the dry or low rainfall stage, a total of 57 species were recorded to flower in this period. A total of 27 species present a flowering stage of more than four months, while 21 species bear fruit in a similar stage of time. Only one species was recorded that flowers and bears fruit all year round: Heliotropium angiospermum. A total of 21 species present synchrony, as both phenological periods coincide in the same reproductive stage.
Key words: Ethnobotanical perception studies; Reproductive phenology; Medicinal plants.
Introduction
The phenology of plant species is one of the main studies to be carried out to understand the flora of an ecosystem (Ortiz, 1990). Phenological studies in general, and especially the flowering and fruiting phases, are essential to determine the optimal time to collect seeds and thus guarantee the management of medicinal, forest, food, melliferous and other species, contributing to possible conservation programs (Albert-Puentes et al., 1995).
Several ethnobotanical studies have been carried out in various coastal communities in the country, with emphasis on the use of traditional medicine in Cuba, including those of Rosette et al. (2019) and Polanco et al., (2011), for the Guanahacabibes and Baconao Biosphere Reserves, respectively; among others Vilamajó (1984) , Vilamajó (1985), Vilamajó and Menéndez (1988), Figueredo et al., (2010), Figueredo et al., (2013). The aim of the work was to publicize the flowering and fruiting patterns of flora species that are components of natural and traditional medicine in the coastal community of Verraco, part of the Baconao Biosphere Reserve.
The population's knowledge and monitoring of the phenology of flora components in the use of natural and traditional medicine enables them to make appropriate use of plants for the treatment and cure of different conditions, and allows them to draw up strategies for the appropriate sowing and cultivation of these species.
Materials and methods
Based on the ethnobotanical studies carried out in the community of Verraco by Polanco et al., (2011), and on the floristic list of Figueredo et al., (2013), the components of the flora that are used in natural and traditional medicine were selected and their phenology was determined in situ, especially the flowering and fruiting phases of each of the identified species, according to the methodology of Albert-Puentes et al., (1993ª).
Monthly monitoring was carried out between 2015 and 2016 in the community of Verraco, located in the geographical coordinates X: 639 080, Y: 139 020, belonging to the Baconao Biosphere Reserve, province of Santiago de Cuba, and the phenological field observations were complemented with reviews of materials belonging to the Spermatophyte Section of the Herbarium BSC "Dr. Jorge Sierra Calzado" of the Centro Oriental de Ecosistemas y Biodiversidad of Santiago de Cuba, noting the phenological data.
As for the duration of flowering and fruiting, the species were classified according to Castillo and Carabias (1982) as:
Short = species whose flowering or fruiting period is < 4 months.
Long = species whose flowering or fruiting period is ≤ 4 months.
According to the flowering period, the criteria of Sarmiento and Monasterio (1983) were followed, where:
Species with continuous flowering, are those that bloom all year round.
Early flowering species, those that bloom at the beginning of the rainy season (spring).
Delayed-flowering species are those that begin in the second half of the rainy season, and extend through the end of that season.
Late-blooming species are those that flower in the dry season.
Opportunistic species: those that are able to flower in any period, as long as the environmental conditions are favorable to them.
For the analysis of the distribution of plant species and the updating of taxonomic nomenclature, the criteria of Greuter and Rankin were followed (2017).
Results and discussion
According to the ethnobotanical perception studies carried out in the community of Verraco, a total of 82 species of flowering plants (angiosperms) were recorded, which are used by the community for the use of natural and traditional medicine, which are grouped into 75 genera and 49 botanical families (Table 1).
Table 1 - Phenology of the species with medicinal use by the Verraco community
| Family | Scientific name | Common name | Distribution | Phenology | |
| Flowering | Fructification | ||||
| Acanthaceae |
|
árnica | Cu - Esp, Ja, PRc | Jan-Sep | Jan-Sep |
|
|
carpintero | PCu - PEsp, PJa | Ap | ||
| Aizoaceae |
|
espinaca | NCu | May | |
| Amaranthaceae |
|
apasote | PCu - PEsp PJa | Ap-June | |
| Anacardiaceae |
|
mango | NCu - PEsp PJa | Jan-Ap | Ap-Aug |
| Annonaceae |
|
guanábana | PCu - PEsp PJa | Nov | |
|
|
anón | CCu - CEsp CJa | Jun | Aug-Sep | |
| Apiaceae |
|
perejil | NCu | Jun | Aug-Sep |
|
|
anís | NCu | Jun | Aug-Sept | |
| Arecaceae |
|
corojo | ˚Cu | Jun | |
|
|
coco | PCu - PEsp PPRc | Nov-Mar | Nov-Mar | |
| Asteraceae |
|
romerillo | Cu - Esp Ja PRc | Mar | Oct |
|
|
manzanilla | Cu - Ja AmN AmC AmS | Feb | ||
|
|
salvia | Cu - Esp Ja PRc | Jan-Jun | Oct-Nov | |
|
|
rompezaragüey | ˚Cu | Jan, Mar/Nov | ||
|
|
guisazo de Baracoa | PCu - AmN NVM | Mar/May/Jul | ||
| Boraginaceae |
|
alacrancillo | Cu - Esp Ja PRc | Whole a year | Whole a year |
| Brassicaceae |
|
berro | NCu(CuW(Art May | May-Sep | Feb |
| Bromeliaceae |
|
piña ratón | NCu - Esp Ja PRc | May-Oct | Nov-Feb |
| Burseraceae |
|
sasafrás | PCu - AmN AmC | Dec-March | Dec-Mar |
|
|
almácigo | Cu - Esp Ja PRc | Feb-Sep | October-Mar | |
|
|
copal | ˚Cu | Apr | ||
| Cactaceae |
|
tuna | NCu - PEsp PJa | Dec-Mar | Oct |
| Canellaceae |
|
cúrbana | Cu - Esp Ja PRc | Jun-Sep | |
| Capparaceae |
|
raíz de berraco | Cu (CuE(Ho Gu)) - | Jun | Aug |
| Caricaceae |
|
fruta bomba, papaya | CCu - CEsp CJa | Feb-Mar | |
| Commelinaceae |
|
barquito, cordobán | NCu - PEsp PPRc | Ap-Jun | |
| Costaceae |
|
caña mexicana | NCu - PEsp PJa | May | |
| Crassulaceae |
|
hoja de aire | NCu - PEsp PJa | Jan-Mar | Ap-Jun |
| Cucurbitaceae |
|
cundeamor | NCu - PEsp PJa | Mar-Jun | Mar/Sep-Oct |
| Cyperaceae |
|
caramaná | PCu - PEsp PJa | May-Oct | May-Oct |
| Erythroxylaceae |
|
jibá | Cu - Esp Men AmS | Jan-Ap | Mar-May |
| Euphorbiaceae |
|
coronilla | Cu - Esp Ja PRc | Ap | |
|
|
túa túa | Cu - Esp Ja PRc | May-Sep | Jun-Oct | |
|
|
chaya | NCu | Nov | ||
| Fabaceae |
|
palo de brasil | Cu (CuW(Mat IJ) | Apr-Jun/Sep-Dec | |
|
|
cañándonga | Cu(CuW(Art) AmS | Feb-May | May-Aug | |
|
|
amor seco | Cu - Esp Ja PRc Men Cay AmN AmC AmS | Jul | Jul | |
|
|
platanillo | DCu(CuW(PR* Art Hab* May Mat IJ) | Feb-Ap | Feb-Ap | |
|
|
tamarindo | PCu(CCuW CCuC | Dec-Mar | Mar-May | |
| Lamiaceae |
|
hierba buena | CCu(CuW(Hab*)) - | Mar | |
|
|
albahaca morada | NCu(CuW(PR* Art Hab* Mat) | Ap | ||
|
|
mejorana | PCu(CCuW(Hab*) | Feb-Ap | ||
|
|
orégano | PCu(CCuW(PR* Art Hab* May Mat IJ) | Ap | ||
|
|
meprobamato | Mar | Jun | ||
|
|
romero | PCu(CCuW CCuC | Mar | ||
|
|
vencedor | C Cu(CuW(Hab*) | Nov | ||
| Lauraceae |
|
aguacate | PCu(CuW(Art Mat) CuC(SS) CuE(Ho | Mar-May | Jun-Sept |
| Lythraceae |
|
resedá | PCu - pHab* CEsp | Ap and Sep | |
|
|
malva bruja | Cu(CuW(PR* Art Hab* May Mat IJ) | Nov | ||
| Malvaceae |
|
algodón | -Cu - VM | Feb/Ap/Jun/ Nov/Dec | |
|
|
guásima | Cu(CuW(PR* Art Hab* May IJ) | Oct-Dec | Oct-Feb | |
|
|
marpacífico rojo | NCu | Mar-Jun | ||
|
|
majagua | NCu | Ap-Jun | ||
| Meliaceae |
|
árbol del nim | PCu(CuW(Hab*)) - | Sep | |
|
|
jubabán | Cu(CuW(PR* Art Hab* May Mat | Jan-June | Jan/Ab/Jun/May/Dec | |
| Moringaceae |
|
moringa | NCu(PCuW(Art cHab*) CCuC(Ci SS) | Mar | May |
| Myrtaceae |
|
eucalipto | Jan/May | Jan-Feb/Oct | |
|
|
guayaba | DCu - Esp Ja PRc | May/Jun | Ap-Jul | |
| Oleaceae |
|
jazmín de 5 pétalos | NCu(CuW(Mat) CuC(VC LT) | Jan/Sept Aug | |
| Papaveraceae |
|
cardo santo | DCu(CuW(PR* Hab* Mat) CuC(VC Ci) | Feb-Ap | |
| Pasifloraceae |
|
marilope o tapón | PPRc Men Bah Cay | Mar/Ap | Mar/Nov |
| Phyllanthaceae |
|
fruta o huevo escondido | ˚Cu | Feb-Ap | Feb-June |
| Phytolaccaceae |
|
anamú | Cu(CuW(PR* Hab* May IJ) CuC(VC Ci SS | Ab/Jun/Ago/ Nov/Dec | |
| Piperaceae |
|
anizón | DCu(CuW(Hab*) CuC(Ci SS) CuE(Gr | Ap-June | Jun-Aug |
| Plantaginaceae |
|
llantén | NCu(CuW(PR* Art Hab* May Mat) | Feb-Jun | Feb-Jun |
| Poaceae |
|
caña santa, hierba de calentura | CCu - CEsp CJa CPRc CMen CBah | Oct | Dec |
| Polygonaceae |
|
uva caleta | Cu - Esp Ja PRc | Feb-Ap | |
| Punicaceae |
|
granada | C Cu - C Esp C Ja C PRc C Men | Oct-Dec | |
| Rhamnaceae |
|
carbonero | Cu - Esp Ja PRc Men Bah Cay AmN | Ap-Jun | |
| Rutaceae |
|
naranja agria | NCu(CuW(Hab* Mat) CuC(Ci Cam) | Jan-May | |
|
|
limón | CCu - CEsp CJa | Jan/Jun/Oct/ Nov | ||
|
|
mandarina | CCu - CEsp CJa CPRc CAmN | Dec-Jan | ||
|
|
ruda | (C)Cu - CEsp CPRc CMen VM | Mar | Jun | |
| Sambucaceae |
|
sauco blanco | NCu | Feb-May | |
| Solanaceae |
|
yerba mora | Cu(CuW(PR* Art Hab* May Mat IJ) | Mar-May/Oct | Mar-May/Jul/Oct-Nov |
|
|
prendejera | Cu (CuW(PR* Art Hab* May Mat IJ) CuC(VC Ci SS | May/Nov | Jan/May Oct/Nov | |
| Urticaceae |
|
lloviznita | CPRc CMen CAmC CAmS Cu - Esp Ja | Ap | |
| Verbenaceae |
|
menta americana | Cu (CuW(PR* Art Hab* May Mat) | Ap/Jun-Jul | |
|
|
verbena | Cu (CuW(PR* Art Hab* May Mat IJ) | Feb-May | Jan/Jul/Oct-Nov | |
| Vitaceae |
|
bejuco ubí | Cu - Esp Ja | Ap | |
| Xanthorrhoeaceae |
|
sábila | PCu - PEsp PJa | Feb-May | Ap-Jun |
Legend: AmC: Central América, AmN: North América (including México), AmS: South América (including Trinidad, Tobago, Curazao, Margarita, etc.), Art: Artemisa province, Bah: Bahamas, CA: Ciego de Ávila province, Cam: Camagüey province, Cay: Islas Caymán, Ci: Cienfuegos province, CuC: central Cuba, CuE: Easter Cuba, CuW: Western Cuba, Esp: Hispaniola, Gr: Granma province, Gu: Guantánamo province, Hab*: La Habana province, Ho: Holguín province, IJ: Pine Island, Ja: Jamaica, Mat: Matanzas province, May: Mayabeque province, Men: minor Antillas (including Virgens Island), PR*: Pinar del Río province, PRc: Puerto Rico, SS: Sancti Spíritus province, VC: Villa Clara province, VM: Old World (including Australia e Pacific Island), Endemic in Cuba, pointing by error, D: Present but doubtfully indigenous, N: naturalized, P: no indigenous but likely naturalized, C: widely cultivated, (C): Occasionally Cultivated or currently not cultivated, Jan: January, Feb: February, Mar: March, Ap: April, May: May, Jun: June, Aug: August, Sep: September, Oct: October, Nov: November, Dec: December.
These results are congruent with Polanco et al., (2011), Rosete et al., (2019), and Nina (2017), who in different ethnobotanical studies on the use of biodiversity for medicinal purposes report that community members use mainly curative and aromatic plant species to address certain health conditions, using not only those available in the natural environment, but also those grown in home gardens and farms, largely using arvenous species.
From the total number of species studied, 46 % (38) were recorded with flowers and fruits, while only 12 % (10) and 39 % (32) were determined in the flowering and fruiting stages, respectively (Table 1). According to Polanco et al., (2011), the fruits and flowers of medicinal plants represent, after the leaves, the organs most used by the inhabitants of this community to combat the various conditions and diseases.
The best represented botanical families are: Lamiaceae with seven species, followed by Asteraceae and Fabaceae with five, while the genera with the largest number of infra-generic taxa were Citrus (Rutaceae) with three species, followed by the genera Annona (Annonaceae), Bursera (Burseraceae), Jatropha (Euphorbiaceae), Plectranthus (Lamiaceae), and Solanum (Solanaceae), with two in each case. In ethnobotanical studies conducted in the community of Gran Piedra, Hernández (2001) reported the Fabaceae and Asteraceae families as the most represented for their medicinal uses.
From the total of species, only four are endemic: Acrocomia crispa C.F. Baker ex Becc, Vernonanthura havanensis (DC.) H. Rob., Protium cubense Urb. and Phyllanthus tenuicaulis Müll. Arg. subsp. tenuicaulis. 24 % of the identified species are naturalized (many of them are fruit trees), while 19 % correspond to non-indigenous but possibly naturalized species, according to Greuter and Rankin (2017). A total of eight species are cultivated.
There is a report on the floristic diversity of Verraco (Figueredo et al., 2013), where 57 % of the total plant species grow in the surroundings of the community and only 44 % are cultivated by the inhabitants.
Pelicié and Hernández (1985) obtained 91 % of endemism at the ethnobotanical level in that coastal sector (Verraco-Cazonal); later, in 1988, they updated the list of medicinal plants for that locality, identifying 59 species with 27 % of endemism.
According to Albert-Puentes et al., (1993), 40 species flowers during the rainy season from May to October, including Bromelia pinguin L. and Cyperus rotundus L., while 36 species bear fruit during the same period, such as Solanun torvum Sw. and Solanum americanum Mill., both of the family Solanaceae.
In the dry or low rainfall stage, a total of 57 species were recorded that bloom during this period, although many of them overlap this phenophase during the months of April and May, which mark the end of the dry stage and the beginning of the rainy stage (Bermúdez and Durán, 1991). The possible dependence of the phenological patterns on environmental factors, as well as their relationship with other plants and animals was pointed out by Ramírez and Brito (1987).
Figure 1 shows the categories of species according to the period in which they bloom, highlighting in this aspect the Late Blooming (FTA) with 31 species, followed by the opportunistic species (CP) with 24, those that bloom at any stage of the year.
For Opler et al., (1976), the phenological phases are closely related to rainfall, given that the seasonality in the tropics is fundamentally given by this climatic factor. Bawa et al., (1990) reported spatial and temporal variations in phenological patterns revealed in individual and community studies over two decades; however, the factors determining these patterns remain unknown (Bawa and NG., (1990).
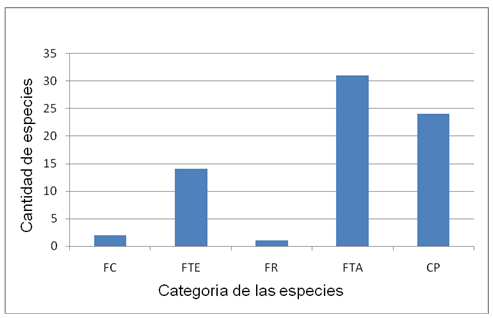
Fig. 1. - Species categories according to flowering period, FC: continuous flowering, FTE: early flowering, FR: delayed flowering, FTA: late flowering, CP: opportunistic species
A plant may not develop all its phenological stages if it grows in different climatic conditions than its region of origin. Abu-Asab et al., (2001) report extensions of the flowering period in 89 species in the Washington DC area, analyzing 30-year records; correlating these observations directly with local increases in minimum temperatures.
According to Castillo and Carabias (1982), and as shown in Figure 2, a total of 45 species (55 % of the total recorded) were identified that bloom in a period of less than four months, as is the case of Argemone mexicana L.(Figure 3); that is, they could be defined as short-lived species. In the same category there are 26 species (32 % of the total recorded), whose fruiting stage does not exceed the same number of months.
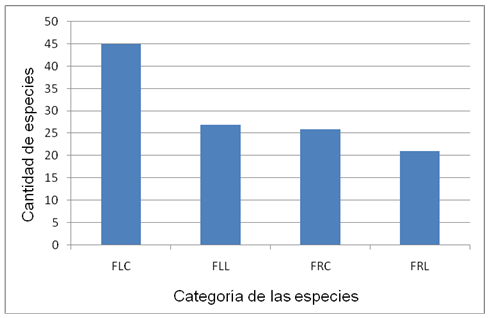
Fig. 2 - Species categories according to flowering and fruiting time. FLC: short period flowering, FLL: long period flowering, FRC: short period fruiting, FRL: long period fruiting
A total of 27 species have a long flowering period and 21 species bear fruit over a period of more than 4 months, as they are: Nasturtium officinale W. T Aiton, Cassia grandis L., Mangifera indica L., among others; many of them are edible and cultivated. Only one species has been recorded that flowers and bears fruit all year round: Heliotropium angiospermum Murria.
Some species are recorded that overlap both phenological periods: Dicliptera sexangularis (L.) Juss, from January to September and C. rotundus, from May to October; both are herbaceous species. Other species, such as Cocos nucifera Lin., and Trichilia hirta L.(Figure 4), present their flowering and fruiting phases in the same period, although the latter presents several fruiting peaks (Table 1).


Other authors, such as Albert-Puentes et al. (1993), carried out phenological studies where they obtained similar results of overlapping with T. hirta(Figure 4) and other tree species such as: Prunus occidentalis Sw. and Matayba apetala f. oppositifolia (A. Rich.) Radlk. Similar fruiting patterns were found by Sánchez et al. (2009) for species analogous to those reported in these localities.
A total of 21 species are synchronized, that is, they flower and bear fruit at the same time, at the same time, as they are: Bursera graveolens Triana & Planch , and Plantago major L.; Augspurger (1983) defined synchronism as an escape of plants from flower predators and then from seeds, so it is an adaptive strategy for successful pollination and seed dispersal (Steven et al., 1987).
For Castillo and Carabias (1982), one aspect to take into account is the variability of phenological patterns (not only between species and years, but between individuals of the same species, so that not all individuals belonging to the same species flower and bear fruit simultaneously, and sometimes not even in the same year).
Bursera simaruba (L.) Sarg. (seedling), extends its flowering for nine months, while fructification is recorded for six months. These results show a very different behavior to the ones obtained by Hechavarría et al. (2000), in Itabo, Matanzas, where fruits are harvested only in July.
It is known that many species, such as Carica papaya L., are able to revert their reproductive habits in extreme conditions, which probably is a mechanism for the perpetuation of the species; for that reason it is important to complement the phenological observations in the field with the herbarium records, because according to Croat (1969) individuals can bloom in a year out of season, producing a lengthening of the period of the observed phenophase.
Conclusions
In the community of Verraco, Baconao Biosphere Reserve, 87 % and 61 % of the components of the flora for medicinal and traditional use are recorded with flowers and fruits, which enables the inhabitants, based on their knowledge of these plant phenophases, to make appropriate use of them for the treatment and cure of various conditions, and to be able to trace strategies for the planting and cultivation of these medicinal species
Acknowledgements
Thanks are due to the national project "Components of biological diversity used by Cuban families in natural and traditional medicine", which is part of the Natural and Traditional Medicine Program of the Ministry of Public Health (MINSAP), for its logistical support for the development of this research.
Referencias bibliográficas
ABU ASAB, M.S., PETERSON, P.M., SHETLER, S.G. y ORLI, S.S., 2001. Earlier plant flowering in spring as a response to global warming in the Washington, DC, area. Biodiversity and Conservation [en línea], vol. 10, pp. 597-612. [Consulta:22/05/2020]. ISSN 0960-3115. Disponible en: Disponible en: http://repository.si.edu/xmlui/handle/10088/3371 . [ Links ]
ALBERT PUENTES, D., LÓPEZ, A., RODRÍGUEZ, M. y DUARTE, M., 1995. Recursos fitogenéticos forestales I. Familia Meliaceae. Fontqueria, vol. 42, pp. 329-351. [ Links ]
ALBERT PUENTES, D., LÓPEZ, A. y ROUDNÁ, M., 1993. Observaciones fenológicas en árboles tropicales. Consideraciones metodológicas. Fontqueria, vol. 36, pp. 257-263. [ Links ]
AUGSPURGER, C.K., 1983. Phenology, Flowering Synchrony, and Fruit Set of Six Neotropical Shrubs. Biotropica [en línea], vol. 15, no. 4, pp. 257-267. [Consulta: 22/05/2020]. ISSN 0006-3606. DOI 10.2307/2387650. Disponible en: Disponible en: https://www.jstor.org/stable/2387650 . JSTOR [ Links ]
BAWA, K.S. y HADLEY, M., 1990. Reproductive Ecology of Tropical Forest Plants [en línea]. S.l.: Unesco. ISBN 978-0-929858-22-7. Disponible en: https://books.google.com.cu/books/about/Reproductive_Ecology_of_Tropical_Forest.html?id=AuHwAAAAMAAJ&redir_esc=y. [ Links ]
BERMÚDEZ, G. y DURÁN, T., 1991. Mapas de precipitación. Atlas de Santiago de Cuba. Cuba: Academia de Ciencias de Cuba. [ Links ]
CASTILLO, S. y CARABIAS, J., 1982. Ecología de la vegetación de las dunas costeras: fenología. Biótica [en línea], vol. 7, no. 4, pp. 551-568. [Consulta: 22/05/2020]. Disponible en: Disponible en: https://biblat.unam.mx/es/revista/biotica-mexico-d-f/6 . [ Links ]
DE STEVEN, D., WINDSOR, D.M., PUTZ, F.E. y DE LEON, B., 1987. Vegetative and reproductive phenologies of a palm assemblage in Panama. Biotropica [en línea], vol. 19, no. 4, pp. 342-356. [Consulta: 28/05/2020]. ISSN 0006-3606. DOI 10.2307/2388632. Disponible en: Disponible en: https://www.jstor.org/stable/2388632 . JSTOR [ Links ]
FIGUEREDO CARDONA, M., POLANCO DURÁN, G. y REVILLA GÓNGORA, Y., 2010. Estudio para la conservación de las plantas útiles de las Terrazas Costeras de la Reserva de la Biosfera Baconao, Santiago de Cuba, Cuba. Sistemas Biocognitivos Tradicionales: Paradigmas en la Conservación Biológica y el Fortalecimiento Cultural [en línea]. México: Asociación Etnobiológica Méxicana, A. C., pp. 36-40. ISBN 978-607-482-095-9. Disponible en: https://www.researchgate.net/publication/280309669_Sistemas_Biocognitivos_Tradicionales_Paradigmas_en_la_Conservacion_Biologica_y_el_Fortalecimiento_Cultural. [ Links ]
FIGUEREDO, l.M., ACOSTA, F., CASTELL M.A., y POLANCO G. 2013. Objetos de conservación de la flora y la vegetación de los cerros calizos costeros de la Reserva de la Biosfera Baconao, Santiago de Cuba. Foresta Veracruzana vol 15 no.1 pp. 9-24. Disponible en: https://www.redalyc.org/jatsRepo/1813/181359681003/html/index.html [ Links ]
GREUTER, W. y RANKIN RODRÍGUEZ, R., 2017. Vascular plants of Cuba. A preliminary checklist. Second, updated Edition of The Spermatophyta of Cuba, with Pteridophyta added [en línea]. Berlin: Botanischer Garten und Botanisches Museum Berlin. ISBN 978-3-946292-18-0. Disponible en: https://www.researchgate.net/publication/321886706_Vascular_plants_of_Cuba_A_preliminary_checklist_Second_updated_Edition_of_The_Spermatophyta_of_Cuba_with_Pteridophyta_added /link/5a381f76458515919e71ed8a/download. [ Links ]
HECHAVARRÍA, O., RODRÍGUEZ, E., MORALES, N., VERA, N., ESPÍN, G., CORRALES, B., FUENTES, V., y PÉREZ, A. 2000. Calendario fenológico de 51 especies forestales de Cuba. Mejoramiento Genético y Semillas Forestales. Revista Forestal Centroamericana 30, Turrialba, Costa Rica.p.5-8. [ Links ]
HERNÁNDEZ, C.J. 2001. Uso de las plantas. En: Diversidad Biológica de los Macizos Montañosos Orientales. Tomo II. Programa Científico-Técnico Nacional. Desarrollo Sostenible de la Montaña. 549 p. [ Links ]
OPLER, P.A., FRANKIE, G.W. y BAKER, H.G., 1976. Rainfall as a Factor in the Release, Timing, and Synchronization of Anthesis by Tropical Trees and Shrubs. Journal of Biogeography [en línea], vol. 3, no. 3, pp. 231-236. [Consulta: 26/05/2020]. ISSN 0305-0270. DOI 10.2307/3038013. Disponible en: Disponible en: https://www.jstor.org/stable/3038013 . JSTOR [ Links ]
ORTIZ, R., 1990. Fenología de árboles en un bosque semideciduo. Acta Botánica Venezuélica [en línea], vol. 16, no. 1, pp. 93-116. [Consulta: 26/05/2020]. ISSN 0084-5906. Disponible en: Disponible en: https://www.jstor.org/stable/41740491 . JSTOR [ Links ]
PELICIÉ, O. y HERNÁNDEZ, J., 1985. Estudio etnobotánico del matorral xeromorfo costero comprendido entre las playas Verraco y Cazonal de Santiago de Cuba. En: Meeting Name: Simposio de Botánica, Memorias: Primer Simposio de Botánica, Palacio de las Convenciones, Editora de la Academia de Ciencias de Cuba, Ciudad de La Habana, Cuba [en línea] Disponible en: https://koha.inpa.gov.br/cgi-bin/koha/opac-detail.pl?biblionumber=13571.580.63 S612 [ Links ]
POLANCO, D. G., SIMÓN, R. F. R., MORELl B. A., y ECHEZARRETA C. C. M. 2011. Uso, consumo y demanda de las plantas medicinales en la ciudad de Santiago de Cuba. Ediciones UO. Santiago de Cuba, Cuba. ISBN: 978-959-207-416-3. [ Links ]
RAMÍREZ, N. y BRITO, Y., 1987. Patrones de floración y fructificación en una comunidad pantanosa tipo morichal (Calabozo-Guàrico, Venezuela). Acta Científica Venezolana [en línea], vol. 38, pp. 276-281. Disponible en: https://www.researchgate.net/publication/281627160_Ramirez_N_Y_Brito_1987_Patrones_de_floracion_y_fructificacion_en_una_comunidad_pantanosa_tipo_morichal_Calabozo -Guarico_Venezuela_Acta_Cientifica_Venezolana_38_276-281. [ Links ]
ROSETE, S., RICARDO, N. y NÁPOLES, R., 2019. Productos forestales no maderables en la Reserva de la Biosfera Península de Guanahacabibes. Biodiversidad, usos tradicionales y conservación de los productos forestales no maderables en Cuba [en línea]. S.l.: s.n., pp. 34-67. ISBN 978-84-9717-376-6. Disponible en: https://www.researchgate.net/publication/335971002_Biodiversidad_usos_tradicionales_y_conservacion_de_los_productos_forestales_no_maderables_en_Cuba. [ Links ]
SÁNCHEZ, J.A., MUÑOZ, B. y MONTEJO, L., 2009. Rasgos de semillas de árboles en un bosque siempreverde tropical de la Sierra del Rosario, Cuba. Pastos y Forrajes [en línea], vol. 32, no. 2, pp. 1-1. [Consulta: 28/05/2020]. ISSN 0864-0394. Disponible en: Disponible en: http://scielo.sld.cu/scielo.php?script=sci_abstract&pid=S0864-03942009000200007&lng=es&nrm=iso&tlng=es . [ Links ]
SARMIENTO, G. y MONASTERIO, M., 1983. Life Form and Phenology. Tropical Savannas. Amsterdam: Elsevier, pp. 79-108. [ Links ]
VILAMAJÓ, D., 1984. Comportamiento fenológico de especies del estrato arbóreo de un bosque siempreverde. Ciencias, vol. 11, pp. 79-92. [ Links ]
VILAMAJÓ, D., 1985. Comportamiento fenológico de especies de vegetación secundaria. Ciencias, vol. 13, pp. 51-56. [ Links ]
VILAMAJÓ, D. y MENÉNDEZ, L., 1988. Fenología de algunas especies importantes en Sierra del Rosario. En: Google-Books-ID: np1JGwAACAAJ, Ecología de los bosques siempreverdes de la Sierra del Rosario, Cuba: Proyecto MAB no. 1, 1974-1987 [en línea]. La Habana: Oficina Regional de Ciencia y Tecnología de la Unesco para América Latina y el Caribe, pp. 243-260. Disponible en: https://books.google.com.cu/books/about/Ecolog%C3%ADa_de_los_bosques_siempreverdes_d.html?id=np1JGwAACAAJ&redir_esc=y. [ Links ]
Received: May 18, 2020; Accepted: July 03, 2020

