










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Cubana de Ciencias Forestales
versão On-line ISSN 2310-3469
Rev CFORES vol.11 no.2 Pinar del Río maio.-ago. 2023 Epub 03-Ago-2023
Original article
Effects of raising Sus scropha on the natural regeneration of Quercus cubana A. Rich
1Estación Experimental Agroforestal Viñales. Pinar del Río, Cuba.
2Universidad de Pinar del Río Hermanos Saíz Montes de Oca. Pinar del Río, Cuba.
The presence of Sus scropha in the areas of Quercus cubana could be affecting the propagation of this species, for which reason two natural areas of oak forest were selected on Alturas de Pizarra of the Viñales Agroforestry Experimental Station. The objective was to determine the effect of raising Sus scropha on the natural regeneration of Quercus cubana. Twenty 100 m 2 plots were established where the vegetation was characterized in terms of composition, diversity and structure. The abundance of pigs in the areas and the relationship between the natural regeneration of Quercus cubana with the intensity of their activity and the measured vegetation variables were determined. In the oak groves studied, there were no significant differences in the associated plant species according to the trend of the rarefaction curves and the non-parametric estimators. In both areas, the herbaceous layer was the best represented followed by the shrub layer in terms of species richness and number of feet, being the ones with the highest incidence in the sociological position. Pigs are common in the areas, and their intensity of activity was considered high and also affects the increase in the latter in the decrease in the natural regeneration of the oak. There are no significant differences between the top and the middle part of the area.
Key words: natural regeneration; oak; pig
INTRODUCTION
Within the growth dynamics of a forest, a number of aspects are present, from natural regeneration, through the stage of development and growth to its climax where it culminates with old age. From a general point of view, many factors come into play, both biotic and abiotic, which increase or decrease said natural regeneration. If we refer to biotic factors, animals take part in this regeneration, being means of transport in the dispersal of seeds that help in the recruitment of new species in new places, even genetics takes part, making the presence of a dominant tree with greater production clear of seeds, which tends to have a greater dissemination in a larger radius, unlike a codominant tree, where its dissemination radius will be lower (Abel Lanas, 2014).
Reyes et al. (2018) state that oak is basically distinguished by the fundamental role that its fruits (acorns) play in feeding pigs, especially the Criollo Pinareño pig in natural breeding. According to González et al (2016), the holm oak forests in Cuba present a critical situation, due to the mismanagement (silvicultural and overgrazing) to which they have been subjected during their development, which has resulted in the aging and deterioration of natural areas and poor growth in plantations, bringing with it a notable decrease in the production of fruits and their quality, affecting the sustained production of these for fattening pigs in rural communities. Due to the aforementioned, this research had as objective: to determine the effect of the breeding of Sus scropha on the natural regeneration of Quercus cubana.
MATERIALS AND METHODS
Location and characteristics of the study area
The present study was carried out in areas of natural oak forests belonging to the Viñales Agroforestry Experimental Station (EEAF), in the mountainous formation known as Alturas de Pizarras in the center of Pinar del Río.
Two areas of natural oak forests on Alturas de Pizarra were selected, having as a requirement the raising of pigs in them and that they were close to water sources, in this case rivers or streams. The first one selected had an average slope of 15 %, located on both sides of one of the tributaries of "La Majagua" river and the other with an average slope of 10 %, close to a stream.
Sampling
In each area, 10 square plots of 10 x 10 m were raised to characterize the vegetation in terms of composition, diversity and structure. Given the nature of the terrain (steep slopes and density of vegetation), they were established randomly, five on each side of the water source. The sampling of the vegetation was carried out between the years 2019 and 2020, since there were no anthropic or extreme environmental events that produced significant changes in the vegetation in these areas during the entire time the sampling lasted.
The vegetation variables that were determined in each selected area and in all plots were the following: plant species and their abundance, tree diameter at a height of 1.30 m (D 1.30), canopy height (H), basal area (G), canopy cover (Cd), soil cover (Cs), natural regeneration and disturbance or damage caused by pigs, taking into account the soil surface removed by them in each of the plots.
The nomenclature, endemism and threat category of the species present in the areas were determined according to the criteria of Berazaín et al. (2005) and Acevedo and Strong (2012).
Alpha diversity (α)
The diversity (alpha) of species in each area was determined using the smoothed accumulation curves of the observed richness (sample-based rarefaction curves) (Gotelli and Colwell, 2001). Taking into account the non-parametric estimators based on abundance (CHAO 1, ACE and Cole) since they assume homogeneity between the habitats according to (Magurran, 2013) and the curves of the number of species represented by an individual (singletons), using the software Estimate S version 9.0.0 Colwell, (2013).
Beta diversity (β)
A cluster analysis was performed to assess the similarity in floristic composition between plots and populations using the PC-ORD software, Version 4.17 (McCune and Mefford, 1999), using the quantitative Sorensen distance measure (Bray-Curtis) and the association method of the groups was that of Wards.
Horizontal structure
The horizontal structure was evaluated by determining the values of abundance, dominance, and the relative frequency of each species (Acosta, Araujo and Iturre , 2006 and Lozada, 2010) ; as well as the value index of ecological importance of the species, IVI, (Lamprecht, 1990; Keels et al., 2002 and Bascopé and Jorgensen, 2005), according to the formula: IVI = Relative abundance + Relative dominance + Relative frequency.
Taking into account that in the middle and lower strata the diameter of the individuals was not measured, the IVI of the same was calculated according to Keels et al. (2002), by means of the sum of the relative abundance and the relative frequency.
The obtained diameters were grouped into diametric classes, from which a frequency histogram was constructed, allowing to know the distribution of the diameters in the studied areas.
Vertical structure
The vertical structure was described taking into consideration the three main strata by which these forests are formed (arboreal, shrubby and herbaceous) and the ferns, according to the methodology used by González et al. (2019).
Species richness in each stratum was determined, as well as their Sociological Position (PS) as an expression of their vertical expansion was determined.
For the analysis of both structures, the scientific name of each species was coded, for which the first three letters of the genus and the specific name were taken.
Abundance and effect of pig activity
To determine the abundance and the effect of the activity of the pigs in the areas, two samplings were carried out in each of the vegetation plots, one in November 2019, coinciding with the oak fruiting period and another in February 2020, after the expiration of said period. Since direct observation of animals is difficult, due to their crepuscular or nocturnal habits, added to their good sense of smell that allows them to perceive human presence from a distance and move away, presence records were made by indirect methods through their tracks. The traces considered were: footprints, excreta, hozaderos or trompeaderos (places in which pigs move or lift the earth with their snouts), wallows (places in which pigs roll on the ground) and scratching posts (structures used by pigs to rub their body and get rid of dried mud and ectoparasites). Direct observations were opportunistic and without any restrictions.
The relative abundance was considered as the percentage of presence - absence of traces in the visits to the plots, following the methodology carried out by Solís et al. (2009), being defined as follows:
Abundant: presence of traces between 76-100 % of the plots.
Common: presence of traces between 51 7 5 % of the plots.
Scarce: presence of traces between 2650 % of the plots.
Rare: presence of traces between 0 25 % of the plots.
To classify the intensity of the activity of the pigs, records were used using the system proposed by Pavlov et al. (1992) with modifications for the study areas, it is based on the count of the total number of traces per area, where a trace consists of the observation of a sign of the presence of the pig (footprints, excreta, holes, scratches and wallows) left by a single individual or a group of individuals, but grouped in the same place. Thus, the activity was recorded as follows:
High: when more than ten traces were recorded in the area.
Moderate: when 4 to 10 traces were recorded in the area.
Low: when there were 1 to 3 tracks in the area.
Null: when there were no traces in the area.
Another important factor for the analysis of the effect of pigs on the ecosystem is related to the slope of the sites where traces were observed, since on steep slopes the activity of pigs can accelerate erosive processes due to the removal of soil due to rooting. Therefore, the slope of the sites with traces was recorded and classified as follows:
Statistical analysis
To determine the relationship between the natural regeneration of Quercus cubana with the intensity of the activity of the pigs and the dasometric variables, a principal component analysis was carried out.
In addition, a one-way ANOVA comparison of means was performed with the HSD Tukey multiple comparison test, having the slope of the area as a factor and natural regeneration and the intensity of the activity of the pigs as dependent variables. As well as a simple linear regression between the natural regeneration and the intensity of the activity of the pigs.
RESULTS AND DISCUSSION
Alpha diversity (α)
The recorded flora included 26 families, 37 genera, 39 species and 4,068 individuals. The families with the highest species richness were Fabaceae (16 %) with six taxa, Poaceae and Rubiaceae with three taxa each (8 %) and Clusiaceae, Melastomataceae, Salicaceae and Sapindaceae with two taxa each (5 %).
In the area four out of ten families registered by Acevedo and Strong (2012) are represented as those with the greatest taxa diversity in the Antilles: Fabaceae, Melastomataceae, Poaceae and Rubiaceae, which highlights the ecological value of the analyzed vegetation.
These results correspond to those found by González et al. (2012) in studies carried out in pine-oak ecosystems in Alturas de Pizarras de Viñales, which also stated that the systematic nature of inadequate silvicultural treatments in this ecosystem threatened the conservation of natural floristic diversity and the occurrence of changes in the optimal ecological conditions for the development of native species of this formation.
Rarefaction curves
The species accumulation curves generated from the non-parametric estimators were analyzed, as shown in Figure 1.
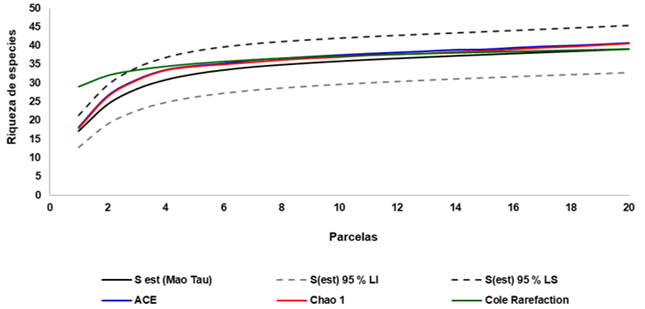
Fig. 1. - Smoothed accumulation curves of observed richness according to the non-parametric estimators ACE, CHAO 1 and Cole, for the holm oak groves studied
The area-species curve indicated that the sampling with 20 plots distributed in the area was sufficient to represent the floristic composition, since it is asymptotic, so it is considered that a good sampling has been achieved, coinciding with what was proposed by Villareal et al. to the. (2006) in which these when they are asymptotic or tend to decrease, indicate that a good sampling has been achieved, being the same representative.
In general, all the non-parametric estimators evaluated were always above the observed richness values, being within the 95 % confidence interval and ending at the same value or close to it.
As the values of the set of estimators behave in a similar way and present values close to those observed, it is considered that a good sampling has been obtained, coinciding with Villareal et al. (2006), so that increases in the sampling effort would not cause substantial increases in species richness.
Beta diversity (β)
Figure 2 shows the dendrogram resulting from the classification analysis of the plots according to species composition.

The cluster analysis shows that despite the fact that the oak groves studied are distant from each other, they present similar floristic characteristics that determine their location in the dendrogram. The plots according to the composition of the species that compose them, were fundamentally grouped into two groups with more than 60 % similarity and another four separated from them.
The grouping of the plots in terms of the composition of the species present in them, could be given by the altitude at which they are found and the proximity to parts of water, since according to Körner (2000) cited by Medina et al. (2010), the vertical dimensions of the mountains produce climatic gradients with abrupt or gradual variations in temperature, relative humidity, solar radiation and precipitation, which causes an effect on the distribution and abundance of flora; also coinciding with what was stated by Álvarez and Varona (2006), in which the distribution and composition of species varies in mountainous areas depending on altitude and proximity to parts of water where other acidophilic species can be found.
Structural analysis of vegetation
Distribution by diameter classes
One of the ways to determine how the intraspecific competition behavior of the stand has been, is through the analysis of the structure by diameter classes. The interval of 2 cm defined, allowed to establish 16 diameter classes. For a better representation of these results, Figure 3 shows the histogram with the distribution of all the trees for each diameter class.
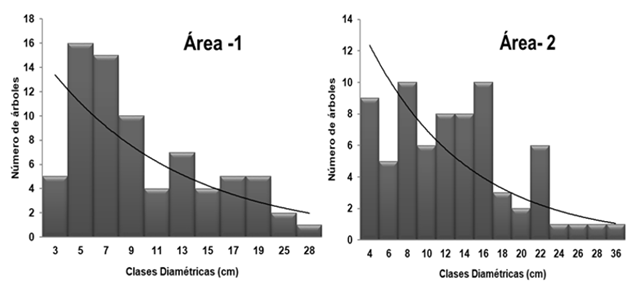
Diameter distribution of vegetation in the holm was decreasing, irregular, typical of a multi-age forest, in the shape of an inverted J where the number of individuals decreases with increasing diameter, which is a physical characteristic of native forests.
This structure allows the assurance of the viability of the populations of the species, since, over time, the regeneration begins to occupy the larger diameter classes, allowing the populations to be renewed in this way. In addition, the concentration of smaller diameter individuals suggests that the vegetation is self-sustaining since they belong to natural regeneration and are capable of establishing themselves during the first years. It can be seen that as the diameter classes increase, the number of individuals decreases, which may be the product of intra and interspecific competition and the light requirements that some of the species require to occupy a place within the vegetation, for so many fail to acclimatize to new conditions and die.
Quercus cubana participates in the diametric structure of the forest in an irregular way, sharing its trees with the most competitive vegetation, which shows a strong interspecific competition, while it dominates in the upper diametric classes.
Importance Value Index (IVI)
To have a broader vision of the importance of each species in the group, the proportion of the IVI is expressed by means of a histogram with respect to the first 20 species with the greatest ecological weight, with the rest of them located within the category of species rare (Figure 4), an approximation of the value of the diversity and heterogeneity of the forest is obtained. The vegetation was characterized as heterogeneous and rich in species since, according to Melo and Vargas (2003), this occurs whenever the greatest ecological weight favors rare species as a whole.
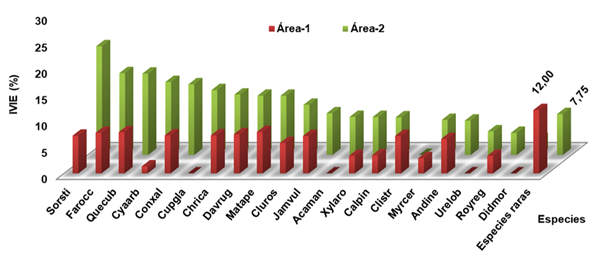 Legend: Sorghastrumstipoides (Sorsti), Farameaoccidentalis (Farocc), Quercus cubana (Quecub), Cyatheaarborea (Cyaarb), Conostegiaxalapensis (Conxal), Cupania Glabra (Cupgla), Chrysobalanusicaco (Chrica), Davilla rugosa (Davrug), Mataybaapetala (Matape), Clusia rosea (Cluros), Jambosa vulgaris (Jamvul), Acacia mangium (Acaman), Xylopiaaromatica (Xylaro), Calophyllumpinetorum (Calpin), Clidemiastrigillosa (Clistr), Myricacerifera (Myrcer), Andira inermis (Andine), Urenalobata (Urelob), Roystonea regia (Royreg), Didymopanaxmorototoni (Didmor
Legend: Sorghastrumstipoides (Sorsti), Farameaoccidentalis (Farocc), Quercus cubana (Quecub), Cyatheaarborea (Cyaarb), Conostegiaxalapensis (Conxal), Cupania Glabra (Cupgla), Chrysobalanusicaco (Chrica), Davilla rugosa (Davrug), Mataybaapetala (Matape), Clusia rosea (Cluros), Jambosa vulgaris (Jamvul), Acacia mangium (Acaman), Xylopiaaromatica (Xylaro), Calophyllumpinetorum (Calpin), Clidemiastrigillosa (Clistr), Myricacerifera (Myrcer), Andira inermis (Andine), Urenalobata (Urelob), Roystonea regia (Royreg), Didymopanaxmorototoni (DidmorFig. 4. - Behavior of the Ecological Importance Value Index (IVIE) in the oak groves studied for the first 20 species and rare species
Among the species with the greatest ecological weight S. stipoides, Q. cubana and C. arborea they occupied the first positions mainly due to their abundance and frequency. It is important to highlight that in the case of Q. cubana, the species that identifies the formation under study was present in all the inventoried plots.
F. occidentalis, a typical species of this type of formation, occupied the second position mainly due to its high abundance and frequency, since it was found in both holm oak groves and in all plots.
Like the previous species M. apetala, a common species in holm oaks, it was determined by its high frequency and abundance within these areas, especially in the lower areas, near water sources.
Vertical structure
To describe and analyze the distribution of the species in a vertical direction, the Sociological Position index was determined for each stratum in each pine forest. As can be seen in Table 1, the herbaceous stratum in both oak groves was where the greatest number of individuals existed, coinciding with the fact that it presented the greatest richness of species, followed by the shrub.
The presence of a greater number of feet in the herbaceous stratum could indicate that they are forests or areas with abundant regeneration, some species presented a distribution in the three strata, while others were present in only one or two. Of the three strata analyzed, the one that had the greatest incidence in the sociological position was the herbaceous one followed by the shrubby one, due to the fact that a greater number of individuals were detected in them, especially due to the natural regeneration of some of the species.
Among the species found in the 3 strata were: Q. cubana, X. aromatica, A. inermis, Bursera simaruba, C. pinetorum, C. rosea, C. xalapensis, C. arborea, F. occidentalis, J. vulgaris, Mastichodendron foetidissimum, M. apetala and M. cerifera; thus, guaranteeing its place in the structure and composition of the forest; of these F. occidentalis was the one that reached the maximum value in terms of sociological position, highlighting its presence in this area.
Of the total number of species registered, five were only found in the tree layer: Abarema obovalis, A. mangium, Mangifera indica, Pinus caribaea and Samanea saman; three were only found in the shrub layer C. sylvestris, Cecropia peltata and Bambusa vulgaris; while they were located only in the herbaceous stratum: Psychotria androsaemifolia, Arthrostylidium cubense, Peltophorum dubium, C. hirsuta, Hypericum hypericoides and S. stipoides, the latter being the most abundant.
Table 1. - Phytosociological value (VF) of each stratum in both oak groves
| holm oak | Stratum | N/ ha | VF (%) | simplified VF |
| AREA 1 | Herbaceous | 9480 | 51.35 | 5.13 |
| shrubby | 7070 | 38.30 | 3.8 | |
| Arboreal | 1910 | 10.35 | 1.03 | |
| AREA-2 | Herbaceous | 17600 | 69.70 | 6.97 |
| shrubby | 6110 | 23.94 | 2.39 | |
| Arboreal | 1540 | 6.10 | 0.61 |
In the two areas studied, the behavior of natural regeneration was dominated by the following species: C. arborea, F. occidentalis, C. xalapensis, M. apétala and C. strigillosa, with a lower number of Q. cubana, which it is also reflected in the shrub layer, which could affect the continuity of the species in the long term.
This coincides with what was reported by Encina et al. (2009) in the holm oak forests of the Sierra de Zapalinamé where the density of juvenile plants and saplings was lower than that of adult trees, which may be due to the competition of shrubs, the grazing effect and the low production of acorns, factors that prevent the establishment of these oaks. The high coverage and dominance of shrubby plants over young individuals and tree saplings suggest that the holm oak forest could be replaced in the medium term by scrubs. A similar sequence has been documented by Casas et al. (1995) for some oak forests in Durango, Mexico.
Relationship and intensity of the activity of pigs, dasometric variables and natural regeneration of Quercus cubana
As a result of the study to determine the relative abundance of pigs in the area considering the percentage of presence-absence of traces in the visits to the plots, it was found that they are common since there was a presence of traces between 5175 % of the monitored plots. This result is similar to that obtained by Solís et al. (2009) who state that this species was common, when evaluating the relative abundance and distribution of pigs in oak forests of the Sierra La Laguna Biosphere Reserve, Baja California, Mexico.
On the other hand, the intensity of the activity of the pigs was considered to be high in the areas taking into account the count of the total number of traces left by them, which could be affecting the natural regeneration of the species and especially in that of Q. cubana, since it feeds on its fruits.
According to Izquierdo et al. (2013) in interviews with one of the farmers in charge of grazing this cattle, he states that in the season of fruiting of the oaks the peasants of the area use the area for illegal breeding, considerably and disproportionately increasing the load of pigs on the site, resulting in the depletion of the fruits before time, with the consequent degradation of the soils and effects on natural regeneration, affecting the natural succession.
Table 2 and Figure 5 are obtained from the principal component analysis, where the components that explain 89.94% of the total explained variance can be seen, as well as a sum of squared load extraction equal to the sum of rotation squared of the rotated components, therefore, the matrix of rotated components does not explain a better solution than the unrotated one.
Table 2. - Total explained variance
| Component | initial eigenvalues | Extraction Sums of Squared Charges | Rotation sums of squared charges | ||||||
| Total | % variance | % accumulated | Total | % variance | % accumulated | Total | % variance | % accumulated | |
| 1 | 3,806 | 47,572 | 47,572 | 3,806 | 47,572 | 47,572 | 3,281 | 41,011 | 41,011 |
| 2 | 2,093 | 26,166 | 73,738 | 2,093 | 26,166 | 73,738 | 2,338 | 29,230 | 70,241 |
| 3 | 1,296 | 16,200 | 89,938 | 1,296 | 16,200 | 89,938 | 1,576 | 19,697 | 89,938 |
| 4 | .478 | 5,971 | 95,909 | ||||||
| 5 | .178 | 2,227 | 98,136 | ||||||
| 6 | .093 | 1,165 | 99,301 | ||||||
| 7 | .049 | .617 | 99,918 | ||||||
| 8 | .007 | .082 | 100,000 | ||||||
Extraction method: principal component analysis.
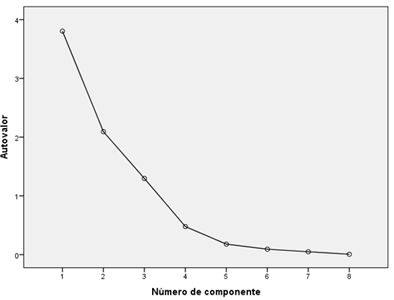
In Figure 5, from the fourth value there is no d-mark dependent on the other values, so it should be considered that only the first three factors should be extracted.
In the component matrix of Table 3, natural regeneration is grouped into the first factor, with soil cover, intensity of pig activity (IACerdos), basal area and plant cover, which indicates a strong relationship between these components within the first function. All these variables saturate in a single factor because they constitute a differentiated group of variables in the correlation matrix. This factor seems to reflect the extent of the spread within the area.
Table 3. - Matrix of components
| variables | Component | ||
| 1 | 2 | 3 | |
| natural regeneration | ,898 | -.308 | -.160 |
| ground cover | .877 | -.384 | -.134 |
| IA Pigs | -.825 | .263 | .384 |
| basal area | ,698 | ,550 | .426 |
| vegetal cover | ,693 | -.356 | .182 |
| Average height | .285 | ,891 | -.237 |
| mean diameter | .591 | .746 | -.207 |
| N/ha | .388 | -.006 | ,889 |
The comparison of means ANOVA of a factor in Table 4, demonstrated having the area as a factor and natural regeneration and IACerdos as dependent variables, the existence of significant differences between the areas with these variables (Tabla 4).

Taking into account that one of the important factors for the analysis of the effect of pigs on the ecosystem is related to the slope of the sites where traces were observed, the HSD Tukey multiple comparison test was selected (Table 5) and as a result of it, significant differences were found between the two areas in terms of natural regeneration, but this was not the case, with natural regeneration and the intensity of the activity of the pigs where no significant differences were reported between the top and the middle part of the area.
This behavior of the top and the middle part can be due to the fact that when there is a slope and the fruits fall, they tend to move naturally towards the lowest parts, in addition, in the rainy season due to water erosion the soil can remain exposed to the slate devoid of vegetation, all this conditions that there are not abundant fruits on the top and the middle part that can favor the natural regeneration of Q. cubana, and that are attractive for the process of rooting in search of them by the of the pigs.
Table 5. - Multiple comparisons HSD Tukey
| Dependent variable | (I) Strata | (J) Strata | Mean difference (IJ) | Standard error | Next. | 95% confidence interval | |
|---|---|---|---|---|---|---|---|
| Lower limit | Upper limit | ||||||
| natural regeneration | top | half | 4.43056 * | .90598 | ,000 | 2,1064 | 6.7547 |
| base | 7.87500 * | 1.26226 | ,000 | 4.6368 | 11.1132 | ||
| half | top | -4.43056 * | .90598 | ,000 | -6.7547 | -2.1064 | |
| base | 3.44444 * | 1.24299 | .033 | .2557 | 6.6332 | ||
| base | top | -7.87500 * | 1.26226 | ,000 | -11.1132 | -4.6368 | |
| half | -3.44444 * | 1.24299 | .033 | -6.6332 | -.2557 | ||
| IA Pigs | top | half | -10.34722 | 4.28028 | .067 | -21.3277 | .6332 |
| base | -36.45833 * | 5.96355 | ,000 | -51.7570 | -21.1597 | ||
| half | top | 10.34722 | 4.28028 | .067 | -.6332 | 21.3277 | |
| base | -26.11111 * | 5.87250 | .001 | -41.1762 | -11.0461 | ||
| base | top | 36.45833 * | 5.96355 | ,000 | 21.1597 | 51.7570 | |
| half | 26.11111 * | 5.87250 | .001 | 11.0461 | 41.1762 | ||
*. The difference in means is significant at the 0.05 level.
From the simple linear regression, a linear relationship between natural regeneration and AIPigs in these ecosystems was obtained.
Table 6 shows the goodness of fit where the model obtained has correlation coefficients (R), coefficient of determination (R2) and estimation error (Sx) acceptable for prediction.
Table 6 Summary of the model
| R. | r squared | adjusted r squared | Standard error of the estimate |
| .875 | .765 | .752 | 1,696 |
The independent variable is damage.
In Table 7, summary of the ANOVA, it can be seen that there is a significant relationship between the variables α<0.05, the F statistic contrasts the null hypothesis that the population value of R is zero.
Table 7. -Significant relationship between the variables
| Sum of squares | gl | Mean square | F | Next. | |
| Regression | 168,400 | 1 | 168,400 | 58,518 | ,000 |
| Residue | 51,800 | 18 | 2,878 | ||
| Total | 220,200 | 19 |
The independent variable is Damage.
For its part, in Table 8 the t value and its critical levels (sig) contrast the null hypothesis that the coefficients are zero in the population α<0.05, the population slope of the regression line is significantly different from zero, which which allows us to conclude that there is a significant linear relationship between natural regeneration and damage.
Table 8. - Non-standardized coefficients
| Unstandardized coefficients | standardized coefficients | t | Next. | ||
|---|---|---|---|---|---|
| B. | Standard error | Beta | |||
| Damage | -,200 | .026 | -.875 | -7,650 | ,000 |
| (Constant) | 12,096 | .584 | 20,715 | ,000 | |
Figure 6 shows the adjustment of the mean equation of natural regeneration for the species under study based on damage.

In this it can be seen, as the intensity of the activity of the pigs increases in relation to the process of feeding on the acorn itself, rooting and the creation of places to rest, the natural regeneration of the seeds that fall to the ground decreases. In the soil, many do not germinate, because the greatest percentage is consumed by rodents or pigs, or they lose their germinative capacity. Free-living pigs can constitute a threat to diversity, because they compete with other species for food and also graze the soil so greatly that they affect plant regeneration.
CONCLUSIONS
The richness of species shows a high floristic diversity associated with the natural holm oak in the study area, where a total of 38 associated species were inventoried, belonging to 25 families, being the Melastomataceae family with four species, Fabaceae, Rubiaceae and Sapindaceae with three each, the best represented.
The raising of pigs in the wild influences the structure and composition of the natural oak forests in the studied area, having a negative effect on the germination of this species, which could affect its future presence in the upper strata.
REFERENCIAS BIBLIOGRÁFICAS
ABEL LANAS, B., 2014. "Bases ecológicas de la regeneración natural" Cuad. Soc. Esp. Cienc. For. 40: 1-18 Disponible en: https://dialnet.unirioja.es/servlet/articulo?codigo=6939789 [ Links ]
ACEVEDO RODRÍGUEZ, P. y STRONG, M.T., 2012. Catalogue of Seed Plants of the West Indies. En: Accepted: 2012-01-01T00:00:00Z, Smithsonian Contributions to Botany [en línea], no. 98. [Consulta: 25/01/2023]. ISSN 0081-024X. Disponible en: Disponible en: http://repository.si.edu/xmlui/handle/10088/17551 . [ Links ]
ACOSTA, V., ARAUJO, P., y ITURRE, M., 2006. Caracteres estructurales de las masas [en línea]. Serie Didáctica no. 22. Agentina: Univerisdad Nacional de Santiago del Estero. Facultad de Ciencias Forestales. ISBN 978-987-1676-34-7. Disponible en: https://fcf.unse.edu.ar/archivos/series-didacticas/SD-22-Caracteres-estructurales-ACOSTA.pdf. [ Links ]
ÁLVAREZ, P.A. y VARONA, J.C., 2006. Silvicultura. La Habana: Ciencia y Técnica. [ Links ]
BASCOPÉ, F., y MOLLER JORGENSEN, P., 2005. Caracterización de un bosque montano húmedo: Yungas, La Paz. Ecología en Bolivia: revista del Instituto de Ecología [en línea], vol. 40, no. 3, pp. 365-379. [Consulta: 27 /01/2023]. ISSN 1605-2528. Disponible en: Disponible en: https://dialnet.unirioja.es/servlet/articulo?codigo=2299508 . [ Links ]
BERAZAÍN ITURRALDE, R., ARECES BERAZAÍN, F., LAZCANO LARA, J.C. y GONZÁLEZ TORRES, L.R., 2005. Lista roja de la flora vascular cubana 2005 [en línea]. España: Jardín Botánico Atlántico de Gijón. [Consulta: 25/01/2023]. Disponible en: Disponible en: http://repositorio.geotech.cu/jspui/handle/1234/1320 . [ Links ]
CASAS, S. R., GONZÁLEZ, E. S., y TENA, F.J.A. 1995. Estructura y tendencias sucesionales en vegetación de clima templado semi-seco en Durango, México. Madroño [en línea], vol. 42, no. 4, pp. 501-515. [Consulta: 27/01/2023]. ISSN 0024-9637. Disponible en: Disponible en: https://www.jstor.org/stable/41426250 . [ Links ]
COLWELL, R.K., 2013. Estimates: Statistical Estimation of Species Richness and Shared Species from Samples [en línea]. 2013. S.l.: EstimateS. Disponible en: http://purl.oclc.org/estimates. [ Links ]
ENCINA DOMÍNGUEZ, J.A., ZÁRATE LUPERCIO, A., ESTRADA CASTILLÓN, E., VALDÉS REYNA, J. y VILLARREAL QUINTANILLA, J.Á., 2009. Composición y aspectos estructurales de los bosques de encino de la Sierra de Zapalinamé, Coahuila, México. Acta botánica mexicana [en línea], no. 86, pp. 71-108. [Consulta: 25/01/2023]. ISSN 0187-7151. Disponible en: Disponible en: http://www.scielo.org.mx/scielo.php?script=sci_abstract&pid=S0187-71512009000100004&lng=es&nrm=iso&tlng=es . [ Links ]
GONZÁLEZ, M., SÁNCHEZ, O., y FIGUEROA, C., 2012. Restauración de la biodiversidad florística en ecosistemas degradados de Pinus caribaea Morelet. Revista Forestal Baracoa, vol. 31, no. 1, pp. 17-23. [ Links ]
GONZÁLEZ-MENÉNDEZ, M., ALONSO-TORRENS, Y., URRUTIA-HERNÁNDEZ, I., MIÑOSO, Y., SANTANA, I., LUIS-SUÁREZ, J., y GONZÁLEZ-TORRES, C.M. 2019. Estructura y composición de encinares naturales en las Alturas de Pizarras del municipio de Viñales, Pinar del Río, Cuba. Madera bosques [en línea], vol.25 no.2 Xalapa ago. 2019 Epub 28-Ene-2020. Disponible en: https://doi.org/10.21829/myb.2019.2521667 [ Links ]
GONZÁLEZ, M., FIDELMANT, M., LEBBET, M., MIÑOSO, Y., SÁNCHEZ, O., FRIOL, P., 2016. Inicio de un programa de manejo silvícola de Quercus cubana A. Rich. en la provincia Pinar del Río, Cuba. Fase I. Memorias del Simposio Internacional sobre Manejo Sostenible de los Recursos Forestales. Cuba: Universidad de Pinar del Río “Hermanos Saíz Montede Oca. [ Links ]
GOTELLI, N., y COLWELL, R., 2001. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters [en línea], vol. 4, no. 4, pp. 379-391. DOI 10.1046/j.1461-0248.2001.00230.x. Disponible en: https://www.researchgate.net/publication/227925773_Quantifying_biodiversity_Procedures_and_pitfalls_in_the_measurement_and_comparison_of_species_richness. [ Links ]
IZQUIERDO MEDERO, K., PIMENTEL CHIRINO, A., y MÁRQUEZ LAM, C.A., 2013. Adaptación de plántulas reintroducidas de Quercus cubana (Fagaceae), endemismo de Cuba Occidental, en su medio natural del bosque de encinos de Manuel Lazo, Sandino, Pinar del Río. Revista Ecovida [en línea], vol. 4, no. 1, pp. 101-110. [Consulta: 27/01/2023]. ISSN 2076-281X. Disponible en: Disponible en: https://revistaecovida.upr.edu.cu/index.php/ecovida/article/view/50 . [ Links ]
KEEL, S., GENTRY, A. y SPINZI, L., 2002. Using Vegetation Analysis to Facilitate the Selection of Conservation Sites in Eastern Paraguay. Conservation Biology [en línea], vol. 7, no. 1, pp. 66-75. DOI 10.1046/j.1523-1739.1993.07010066.x. Disponible en: https://www.researchgate.net/publication/227863765_Using_Vegetation_Analysis_to_Facilitate_the_Selection_of_Conservation_Sites_in_Eastern_Paraguay. [ Links ]
KÖRNER, C., 2000. Why are there global gradients in species richness? Mountains might hold theanswer. Trends in Ecology and Evolution 15: 513-514. https://www.cell.com/trends/ecology-evolution/fulltext/S0169-5347(00)02004-8. DOI: https://doi.org/10.1016/S0169-5347(00)02004-8 [ Links ]
LAMPRECHT, H., 1990. Silvicultura en los trópicos: los ecosistemas forestales en los bosques tropicales y sus especies arbóreas; posibilidades y métodos para un aprovechamiento sostenido [en línea]. Alemania: TZ-Verlag-Ges. ISBN 978-3-88085-440-6. Disponible en: https://books.google.com.cu/books/about/Silvicultura_en_los_tr%C3%B3picos.html?id=1H0JywAACAAJ&redir_esc=y. [ Links ]
LOZADA, J., 2010. Consideraciones metodológicas sobre los estudios de comunidades forestales. Revista Forestal Venezolana [en línea], vol. 54, no. 1, pp. 77-78. Disponible en: https://www.researchgate.net/publication/47512174_Consideraciones_metodologicas_sobre_los_estudios_de_comunidades_forestales. [ Links ]
MAGURRAN, A.E., 2013. Ecological Diversity and Its Measurement [en línea]. Estados Unidos: Springer Science & Business Media. ISBN 978-94-015-7358-0. Disponible en: https://books.google.com.cu/books/about/Ecological_Diversity_and_Its_Measurement.html?id=X7b7CAAAQBAJ&redir_esc=y. [ Links ]
MCCUNE, B., y MEFFORD, M.J., 1999. PC-ORD: Multivariate Analysis of Ecological Data; Version 4 for Windows; [user' S Guide] [en línea]. Estados Unidos: MjM Software Design. Disponible en: https://books.google.com.cu/books/about/PC_ORD.html?id=pZPQOwAACAAJ&redir_esc=y. [ Links ]
MEDINA MACÍAS, M.N., GONZÁLEZ BERNAL, M.A., y NAVARRO SIGÜENZA, A.G., 2010. Distribución altitudinal de las aves en una zona prioritaria en Sinaloa y Durango, México. Revista mexicana de biodiversidad [en línea], vol. 81, no. 2, pp. 487-503. [Consulta: 30/01/2023]. ISSN 1870-3453. Disponible en: Disponible en: http://www.scielo.org.mx/scielo.php?script=sci_abstract&pid=S1870-34532010000200021&lng=es&nrm=iso&tlng=es . [ Links ]
MELO CRUZ, O.A., y VARGAS RÍOS, R., 2003. Evaluación ecológica y silvicultural de ecosistemas boscosos [en línea]. Colombia: Universidad del Tolima. [Consulta: 27/01/2023]. ISBN 956-9243-03-07. Disponible en: Disponible en: https://docplayer.es/7014511-Evaluacion-ecologica-y-silvicultural-de-ecosistemas-boscosos.html . [ Links ]
PAVLOV, P., CROME, F., y MOORE, L., 1992. Feral Pigs, Rainforest Conservation and Exotic Disease in North Queensland. Wildlife Research [en línea], vol. 19, no. 2. DOI 10.1071/WR9920179. Disponible en: https://www.researchgate.net/publication/240505662_Feral_Pigs_Rainforest_Conservation_and_Exotic_Disease_in_North_Queensland. [ Links ]
REYES, A., SANTANA, I., y SÁNCHEZ, N., 2018. El encino cubano. Un recurso fitogenético pinareño desatendido. Boletín Técnico Porcino, no. 37, pp. 3-7. ISSN 2077-4745. [ Links ]
SOLÍS CÁMARA, A.B., ARNAUD FRANCO, G., ÁLVAREZ CÁRDENAS, S., GALINA TESSARO, P., y MONTES SÁNCHEZ, J.J., 2009. Evaluación de la población de cerdos asilvestrados (Sus scrofa) y su impacto en la Reserva de la Biosfera Sierra La Laguna, Baja California Sur, México. Tropical Conservation Science [en línea], vol. 2, no. 2, pp. 173-188. [Consulta: 27/01/2023]. ISSN 1940-0829. DOI 10.1177/194008290900200205. Disponible en: Disponible en: https://bioone.org/journals/tropical-conservation-science/volume-2/issue-2/194008290900200205/Evaluaci%c3%b3n-de-la-poblaci%c3%b3n-de -cerdos-asilvestrados-Sus-scrofa-y/10.1177/194008290900200205.full . [ Links ]
Received: January 20, 2023; Accepted: July 19, 2023

