
Ureides LEVELS IN SOYBEAN PLANTS WITH DIFFERENT INOCULANTS AND SUBMITTED TO WATER DEFICIT.
J.A. Freixas1, Dr. C. W. Torres2, Dra. C. Inés M. ReynaldoI3, Dra. C. María C. Nápoles4
1 Reserva Científica. Email: freixas@inca.edu.cu
2 Investigador Titular ]]>
3 Investigadora Titular
4 Investigadora Titular. Instituto Nacional de Ciencias Agrícolas (INCA), Gaveta Postal 1, San José de las Lajas, Mayabeque, CP 32700.
RESUMEN
La soya [Glycine max (L.) Merr.] establece simbiosis con bacterias fijadoras del nitrógeno del grupo de los rizobios, específicamente del género Bradyrhizobium sp. El nitrógeno fijado en el nódulo es transferido hacia las hojas en forma de ureidos, los cuales incrementan en las hojas ante condiciones de sequía. De hecho, la fijación biológica del nitrógeno en la soya es muy sensible a la sequía y los ureidos parecen estar involucrados a través de un mecanismo de retroalimentación negativa desde las hojas hacia los nódulos. Adicionalmente, la inducción de los genes de la nodulación en Bradyrhizobium sp. tiene efectos positivos en el crecimiento de la soya cuando el déficit hídrico es moderado. El objetivo de este trabajo consiste en el estudio de los niveles de ureidos en plantas de soya con diferentes inoculantes en respuesta al déficit hídrico. Las plantas de soya (var. INCASoy27) crecieron en vermiculita con solución nutritiva por 15 días. Posteriormente, se estresaron por un período de 20 días añadiendo PEG 6000 a la solución. Bradyrhizobium elkanii ICA 8001 fue la cepa utilizada en este experimento para inocular las plantas de soya. Esta cepa creció separadamente en tres medios de cultivo, dos de ellos inducidos en la producción de factores de la nodulación. Se observó un incremento estadísticamente significativo en los niveles de ureidos en hojas y en nódulos de plantas con estrés y con inóculo sin inducir. Sin embargo, este incremento solo se observó en plantas con estrés y con inóculo inducido, lo cual pudiera sugerir un papel importante en la inducción de factores de la nodulación sobre la regulación de los niveles de ureidos en plantas de soya con déficit hídrico. Al final del tratamiento de estrés, se evaluó la actividad de la enzima glutamina sintetasa en nódulos de soya, pero no se detectaron diferencias estadísticamente significativas entre cada grupo de tratamiento, aunque si se observaron diferencias significativas entre las plantas con y sin estrés y con inóculo sin inducir.
Palabras clave: soya, Bradyrhizobium, déficit hídrico, ureidos, factores de nodulación.
ABSTRACT
Soybean [Glycine max (L.) Merr.] establishes symbiosis with N2 fixing bacteria from Rhizobia group, such as Bradyrhizobium sp. Nitrogen fixed in nodules is transferred to leaves as ureides, these molecules are able to increase its levels in soybean leaves under drought stress. Indeed, biological nitrogen fixation in soybean is especially sensitive to drought and ureides seem to be involved through a negative feedback from leaves to nodules. In addition, induction of nodulation genes in Bradyrhizobium sp. has positive effects in soybean growth under moderate drought stress. The goal of this work is to study ureides level in soybean with different inoculants in response to water deficit. Soybean plants (cv. INCAsoy27) were firstly grown in nutrient solution during 15 days. Afterwards, they were drought stressed for a 20 days period adding 10% PEG 6000. Bradyrhizobium elkanii ICA 8001 was the strain cultivated with the purpose of inoculating soybean plants. This strain was separately grown in three culture media, two of them induced in nodulation factor production and the other one without induction. A statistically significant increase of ureides level in leaves and nodules was observed in plants with water deficit and inoculants without nodulation factor induction (NFI). However, this increase was not observed in plants drought stressed and inoculated with the strain previously induced in nodulation factor production. It might suggest an important role of NFI in soybean ureides level regulation under drought conditions. At the end of the stress period, the activity of the enzyme glutamine synthetase in soybean nodules was measured, but there were no statistically significant differences among each treatment with NFI, although differences were reported between non-stressed and stressed soybean plants inoculated with B. elkanii ICA 8001 without NFI.
Key words: soybean, Bradyrhizobium, water deficit, ureides, nodulation factors.
La soya [Glycine max (L.) Merr.] es capaz de establecer simbiosis con bacterias del género Bradyrhizobium que reducen el nitrógeno atmosférico en amonio, el cual es asimilado por la planta evitando el uso de fertilizantes nitrogenados (1). Durante el diálogo molecular que se establece entre Bradyrhizobium y la soya, la bacteria es atraída hacia los pelos radicales de la planta debido a la exudación por ellos de diferentes compuestos, fundamentalmente flavonoides (2). Estos compuestos activan los genes de la nodulación en Bradyrhizobium, estimulando la producción de factores de la nodulación (factores Nod) y la consecuente inserción de la bacteria en la raíz (3). Los factores Nod inducen respuestas moleculares en la raíz de la planta que conllevan a la formación del nódulo radicular (1). En el nódulo, mediante la enzima nitrogenasa, la bacteria es capaz de reducir el nitrógeno atmosférico en amonio, proceso conocido como fijación biológica del nitrógeno (FBN) (4). La mayor cantidad del amonio resultante de la FBN se utiliza en el nódulo para la síntesis de novo de las purinas (5), las cuales son entonces degradadas en ureidos (alantoína y alantoato) que constituyen la forma de transporte del nitrógeno desde los nódulos hasta las hojas a través del xilema (6).
La sequía y su impacto negativo sobre la fijación biológica del nitrógeno pueden limitar el cultivo de la soya en regiones áridas y semiáridas del planeta. King y Purcell (2005) han demostrado que la soya es más sensible a la sequía que otras plantas leguminosas. Sin embargo, las causas que provocan la inhibición de la enzima nitrogenasa en la bacteria, por la sequía, no han sido completamente dilucidadas. Se ha demostrado que la acumulación foliar de ureidos en respuesta al déficit hídrico es menor en genotipos de soya tolerantes a este estrés que en genotipos sensibles (8).
Debido a esta acumulación de ureidos y la disminución de la fijación biológica del nitrógeno bajo condiciones de sequía, se ha propuesto la participación de estos compuestos en un mecanismo de inhibición sobre la FBN. Sin embargo, el suministro externo de ureidos, con el objetivo de mimetizar las respuestas fisiológicas de la soya a la sequía, no permite que estos compuestos lleguen al nódulo a través del xilema (7).
Adicionalmente, King y Purcell (2005) sugieren que, bajo condiciones de déficit hídrico, elevadas concentraciones de ureidos en hoja o de asparagina en nódulo no inhiben la FBN. Sin embargo, elevados niveles de ureidos y de asparagina en el nódulo están asociados con la disminución de la FBN para genotipos de soya sensibles y tolerantes al déficit hídrico. Más reciente, Ladrera y colaboradores (2007) demostraron que la acumulación foliar de ureidos en plantas de soya sensibles y tolerantes a la sequía, no están involucradas en estadios tempranos de la FBN. No obstante, sugieren un importante papel de estos compuestos nitrogenados en estadios más avanzados de la FBN con un estrés por sequía más prolongado (9). En este sentido, los resultados de Ladrera y colaboradores (2007) muestran una acumulación de ureidos mayor y más temprana en genotipos de soya sensibles a la sequía, la cual parece estar relacionada con la sensibilidad de la FBN a este estrés.
Se han realizado investigaciones dirigidas a lograr una mejor respuesta de la soya en condiciones adversas. Atti y colaboradores (2005) han demostrado que la aspersión foliar de factores Nod tiene efectos positivos en el crecimiento de la soya cuando el estrés por sequía es moderado.
Los resultados de Nápoles y colaboradores (2009) fueron similares, pero induciendo los genes nod en Bradyrhizobium, con la consecuente síntesis de factores Nod. Se ha señalado que los factores Nod incrementan el cierre estomático en hojas de soya bajo condiciones de déficit hídrico (10). Este efecto aumenta la retención de agua por la planta, como resultado de una disminución en la transpiración (12). Atendiendo al efecto de otros tipos de estrés abiótico, Duzan y colaboradores (2004) demostraron que el suministro de altos niveles de factores Nod en raíces de soya disminuye los efectos de la acidez y las altas temperatura, pero no el efecto de la salinidad. Sin embargo, los autores no estudiaron el efecto de los factores Nod en raíces de soya bajo condiciones de déficit hídrico.
MATERIALES Y MÉTODOS
Cultivo del microorganismo
La cepa utilizada en los experimentos fue Bradyrhizobium elkanii ICA 8001, la cual se utiliza ampliamente en el desarrollo de inoculantes para la siembra de soya en Cuba. Se multiplicó bajo condiciones de esterilidad en 40 mL, a pH 6,8, de tres medios de cultivo; Extracto de Levadura-Manitol (LM) (14) y un medio de cultivo (Bradyfact) formulado a partir de extracto de semillas de soya y melaza, que se ha demostrado que induce los genes nod en Bradyrhizobium spp. (15). Adicionalmente, el medio Extracto de Levadura-Manitol se suplementó con 10 µM de genisteína (SIGMA) (Genisteína), para inducir la síntesis de los factores Nod (13). Los tres medios de cultivo inoculados con la cepa B. elkanii ICA 8001 se mantuvieron en zaranda orbital a 150 rpm y 30oC, durante 72 horas y finalmente se utilizaron para inocular las plantas de soya. Para la determinación del número de unidades formadoras de colonias (UFC), la cepa se cultivó en el medio LM con 1.5% de agar bacteriológico (SIGMA).
Crecimiento de las plantas
Se emplearon semillas certificadas de soya, cultivar INCAsoy-27 (17), procedentes del banco de germoplasma de semillas del Instituto Nacional de Ciencias Agrícolas, Cuba. Las semillas se desinfectaron con etanol al 98% durante 2 min y posteriormente con hipoclorito de sodio al 2% durante 10 min, los cuales se eliminaron con abundante agua desionizada estéril. Consecutivamente, se colocaron 10 semillas por placa de Petri con 10 mL de medio agar-agua estéril (0.75% m/v) y se incubaron en la oscuridad a 27oC. A las 72 horas, las semillas pregerminadas se colocaron en macetas plásticas estériles de 500 mL, que contenían vermiculita mezclada con solución nutritiva, formulada por Norris y Date (1976). Posteriormente, se inocularon 700 µL de los cultivos bacterianos por semilla de soya pregerminada, a una concentración promedio del microorganismo de 7.92 x 108 UFC.mL-1. Se utilizaron 160 plantas en total y se distribuyeron 40 plantas por tratamiento, compuestos por el grupo de plantas no inoculadas e inoculadas con la cepa B. elkanii ICA 8001 crecida en los medios de cultivo: Bradyfact, Genisteína y LM, para constituir 4 tratamientos. Las plantas se desarrollaron en cámara de crecimiento entre 21-27oC de temperatura, 16/8 horas de luz/oscuridad, 60-70% de humedad relativa, 700 µmol.m-2.s-1 de PPFD (Densidad del Flujo de Fotones Fotosintéticos, por sus siglas en inglés) y circulación constante de solución nutritiva durante 35 días.
Inducción del déficit hídrico
El día 15 de sembradas las semillas, se escogieron al azar 20 plantas por cada tratamiento para la aplicación del estrés hídrico por defecto, conformándose entonces 8 tratamientos. Para ello, se eliminó la circulación de solución nutritiva en estas plantas y se aplicaron 150 mL por maceta, cada 48 horas, de una solución de polietilenglicol (PEG 6000) al 10% m/v con -0.15 MPa de potencial osmótico, que es capaz de inducir un déficit hídrico en las plantas (18). Esta solución de PEG 6000 se aplicó sobre el soporte sólido de las plantas de soya (vermiculita), durante 20 días. La otra mitad del número de plantas por cada tratamiento se mantuvo sin estrés hasta el final del experimento.
Evaluación del Contenido Relativo de Agua
]]> Se tomó el primer nudo de hojas trifoliadas en el momento del alba, con 3 repeticiones por tratamiento a los 4, 8, 12, 16 y 20 días de la aplicación del PEG 6000. Para evaluar el CRA se determinó la masa fresca (MF), la masa seca (MS) y la masa turgente (MT) de las muestras tomadas según el método de Purcell y Sinclair (1995).Determinación de ureidos
Las extracciones de ureidos se realizaron en hojas y nódulos, a partir de 3 plantas por cada tratamiento, a los 4, 8, 12, 16 y 20 días de aplicado el estrés. Para ello, se tomaron nódulos al azar y ponches de 10 mm de diámetro correspondientes al segundo nudo de hojas trifoliadas. Las muestras se secaron en una estufa (BINDER) a 85oC durante 30 min y las extracciones se realizaron a partir de 10 mg de masa seca en 1 mL de NaOH a 0.2 Mol.L-1, que se colocaron en baño de María durante 30 min. Finalmente, los extractos se centrifugaron a 13000 rpm por 5 min y los ureidos se determinaron según el método de detección espectrofotométrica de Young y Conway (1942). La cuantificación de ureidos se realizó a partir de una curva de calibración de alantoína (SIGMA) con un rango de concentración de 0.005-0.3 µMol.mL-1. Los niveles de estos compuestos nitrogenados se expresaron en µMol de ureidos.g-1 de masa seca (gms).
Determinación de la actividad glutamina sintetasa
Para realizar la extracción de la enzima glutamina sintetasa (GS1), se tomaron 0.5 g de nódulos a partir de 3 plantas por cada tratamiento, a los 20 días de impuesto el estrés. Los nódulos se maceraron en un mortero de porcelana con nitrógeno líquido y se añadieron 5 mL del tampón de extracción (pH 7.8) que contenía 100 mMol.L-1 de Tris-HCl, 0.5 mMol.L-1 de EDTA, 1 mMol.L-1 de DTT y 10% de etilenglicol. Finalmente, los extractos se filtraron a 4oC y se centrifugaron a esa temperatura y 15000 rpm durante 20 min. Para determinar la actividad de la enzima GS1, se utilizó glutamato de sodio e hidroxilamina (SIGMA) como sustratos y se evaluó la formación de ?-glutamilmonohidroxamato (?-GMH). El ?-GMH se detectó a 540 nm después de la adición de cloruro férrico, según el método discontinuo de Rhodes y colaboradores (1975). El ?-GMH se cuantificó a partir de una curva de calibración de ?-GMH (SIGMA) a un rango de concentración de 0.1-1.5 µMol.mL-1. Simultáneamente, se determinó la concentración de proteínas totales en los extractos nodulares a través del método de Bradford (1976) y se utilizó albúmina de suero bovino (SIGMA) como estándar para la curva de calibración, con un rango de concentración de 0.5-5 mg.mL-1. La actividad específica de la enzima GS1 se expresó en µMol de ?-GMH.mg-1de proteínas totales. h-1.
Diseño experimental y procesamiento estadístico
Todos los ensayos se realizaron dos veces y los resultados que se muestran corresponden a la media de ambas repeticiones. Los tratamientos se aplicaron siguiendo un diseño completamente aleatorizado y las determinaciones realizadas muestran los intervalos de confianza para a=0.05. El programa StatGraphic 5.1 se utilizó para el procesamiento estadístico de los datos.
RESULTADOS
La figura 1 representa el comportamiento del CRA en las plantas de soya (cv. INCAsoy-27) con los diferentes tratamientos impuestos. La línea superior en negro muestra la media del CRA para todos los tratamientos sin estrés, dada la similitud de este indicador en las plantas sin déficit hídrico. Las plantas sometidas a estrés e inoculadas con la cepa B. elkanii ICA 8001 cultivada en los medios Bradyfact y Genisteína, mostraron un CRA similar ante la imposición del déficit hídrico. Sin embargo, el CRA de las plantas inoculadas con esta cepa multiplicada en el medio LM tuvo un comportamiento inferior, que fue similar en algunos momentos al CRA de las plantas sin inocular. Ambos tratamientos (LM y el control sin inocular) mostraron valores de CRA estadísticamente inferiores al de las plantas inoculadas con la cepa cultivada en los medios Bradyfact y Genisteína, a partir de los 12 días de la inducción del déficit hídrico (Fig. 1).
]]>
Por otra parte, las plantas de soya inoculadas con la cepa B. elkanii ICA 8001 cultivada en el medio LM, mostraron un CRA de aproximadamente un 75%, que fue estadísticamente superior al CRA de las plantas sin inóculo a los 20 días de impuesto el tratamiento de estrés. Sin embargo, ambos valores de CRA alcanzados por estas plantas, se corresponden con un déficit hídrico severo según Hsiao y colaboradores (1973). Cabe destacar que, a los 20 días de impuesto el déficit hídrico, las plantas inoculadas con esta cepa cultivada en los medios Bradyfact y Genisteína alcanzaron valores de CRA de aproximadamente un 85%, que corresponde a un déficit hídrico moderado (Hsiao y cols., 1973). Estos resultados concuerdan con los presentados por Nápoles y colaboradores (2009), donde el CRA en plantas de soya con déficit hídrico impuesto y tratadas con inoculante a base de Bradyrhizobium japonicum inducido con genisteína, fue estadísticamente superior al de plantas con inóculo sin inducir.
La figura 2 muestra los niveles de ureidos foliar en las plantas de soya sin estrés (A) y en las plantas sometidas a estrés hídrico por defecto (B) para los diferentes tratamientos. En las hojas de las plantas con inoculantes, los niveles de ureidos superan a los de plantas sin inocular, tanto en condiciones normales como de estrés. En las plantas de soya inoculadas con B. elkanii ICA 8001 cultivada en el medio Genisteína y sin ser sometidas al déficit hídrico, los niveles de ureidos en hojas fueron iguales a los de plantas inoculadas con esta cepa multiplicada en el medio Bradyfact a los 4, 16 y 20 días de impuesto el estrés (Fig. 2A).
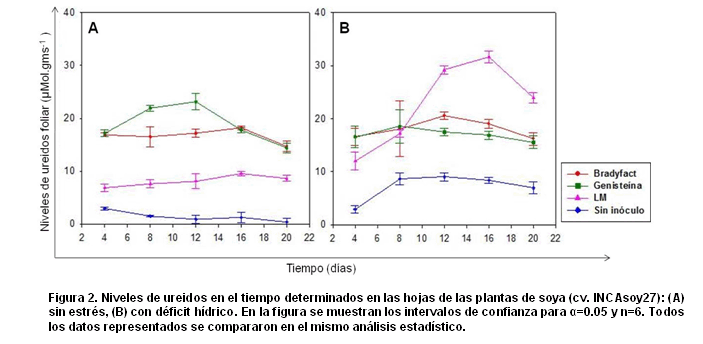
Por otra parte, las plantas de soya inoculadas con la cepa B. elkanii ICA 8001 multiplicada en el medio LM, experimentaron ligeras variaciones en los niveles de ureidos foliar cuando no fueron sometidas a estrés. Otros tratamientos sin estrés muestran niveles de ureidos en hojas con pocas variaciones en el tiempo, entre ellos se destacan las plantas sin inóculo y las plantas inoculadas con esta cepa cultivada en el medio Bradyfact.
Sin embargo, cuando se impuso el déficit hídrico, los niveles de ureidos en las plantas inoculadas con la cepa B. elkanii ICA 8001 sin inducir (LM), fueron significativamente superiores, incluso con respecto a los de plantas inoculadas con esta cepa cultivada en los medios de cultivo Bradyfact y Genisteína. Estos resultados muestran una acumulación de ureidos foliar en las plantas de soya con el inóculo que no presentaba inductores de los genes nod (LM), a partir de los 8 días de impuesto el estrés hasta el final del experimento.
Los niveles de ureidos en hojas de las plantas de soya, con y sin estrés e inoculadas con la cepa B. elkanii ICA 8001 multiplicada en el medio Bradyfact, sólo tuvieron diferencias significativas el día 12 de la evaluación, con incrementos en este indicador para la condición de estrés. Adicionalmente, se observa un incremento significativo en los niveles de ureidos en hojas de plantas sin inoculante y con déficit hídrico en relación con los niveles de ureidos en estas plantas sin estrés (Fig. 2).
Todos los tratamientos que fueron sometidos a estrés, mostraron un incremento en los niveles de ureidos foliar a los 20 días de impuesto el déficit hídrico, siendo más acentuado en las plantas inoculadas con la cepa B. elkanii ICA 8001 crecida en el medio LM. Los resultados presentados se relacionan con los propuestos por King y Purcell (2005), quienes observaron una acumulación de ureidos foliar en las plantas de soya sometidas a un déficit hídrico. ]]>
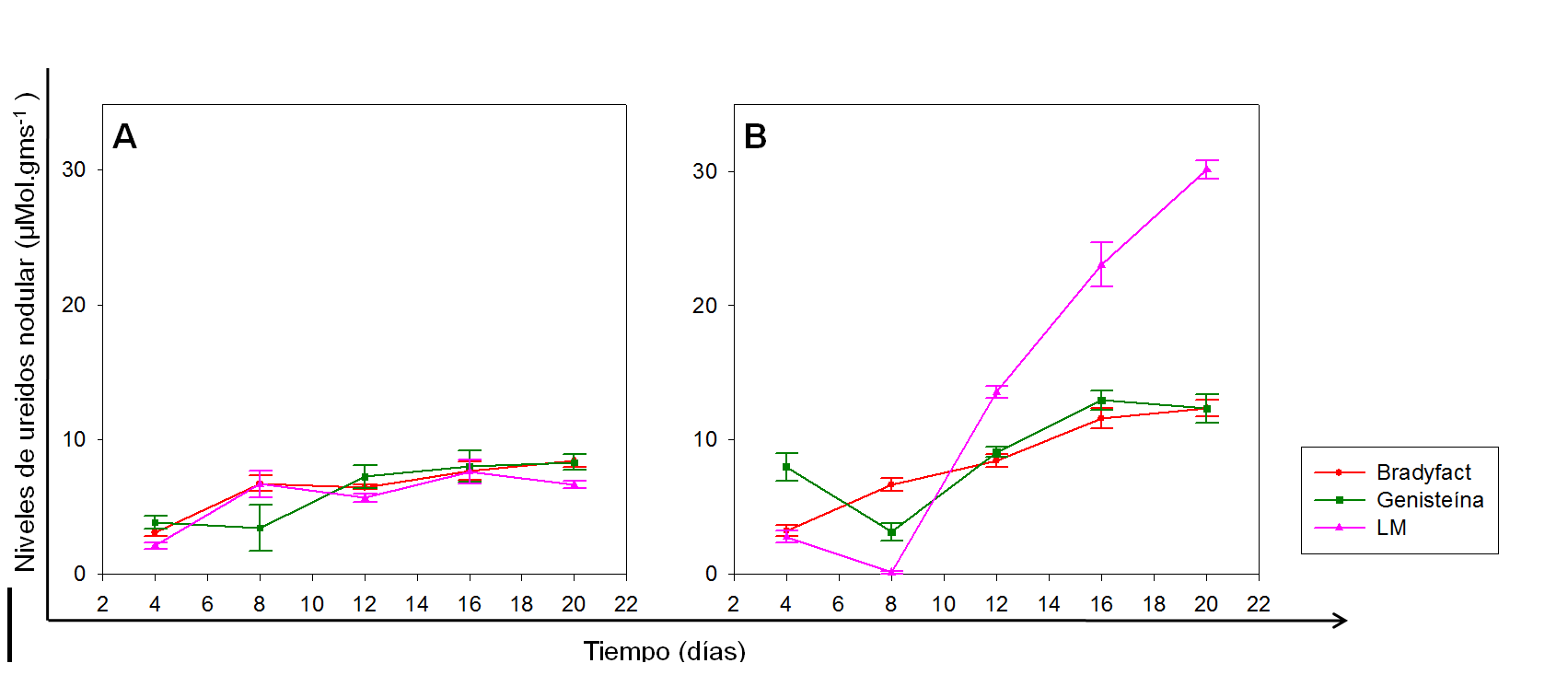
El comportamiento de los niveles de ureidos en los nódulos de las plantas de soya para los diferentes tratamientos de inoculación se muestra en la figura 3 En las plantas sin estrés, no se observaron diferencias significativas en el tiempo que evidenciaran un comportamiento distintivo para algún tratamiento de inoculación. (Fig. 3A).
En contraste, todos los tratamientos con estrés mostraron un incremento significativo en los niveles de ureidos nodular, con respecto a las plantas sin estrés a partir del día 8 de impuesto el déficit hídrico (Fig. 3B). Posterior a ese momento, las plantas inoculadas con la cepa B. elkanii ICA 8001 cultivada en el medio LM y sometidas a déficit hídrico, tuvieron un incremento en los niveles de ureidos significativamente superior a los del resto de las plantas estresadas.
Estos resultados concuerdan con los presentados por Ladrera y colaboradores (2007), quienes demostraron que bajo condiciones de un déficit hídrico, existió una acumulación de ureidos en nódulos de las plantas de soya sensibles a la sequía. De manera similar a la acumulación de ureidos en hojas, no se ha estudiado la posible relación entre la inducción de los genes nod en Bradyrhizobium spp. y la acumulación de estos compuestos en nódulos de plantas de soya bajo condiciones de sequía.
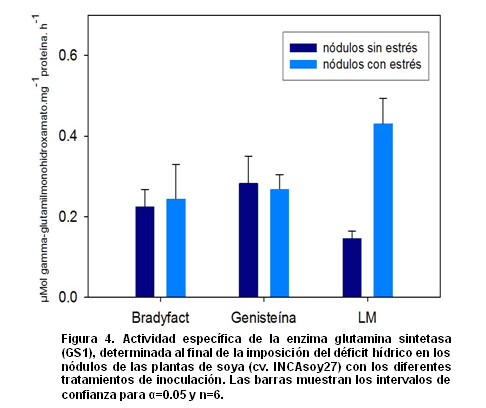
Al finalizar el experimento, se determinó la actividad específica de la enzima GS1 en la fracción de proteínas solubles correspondiente a los nódulos formados sobre las raíces principales y secundarias de las plantas de soya (cv. INCAsoy-27), sometidas o no a déficit hídrico y con los diferentes tratamientos de inoculación.
De manera general, la actividad específica de la enzima GS1 en los nódulos de estas plantas, con y sin déficit hídrico e inoculadas con la cepa B. elkanii ICA 8001 cultivada en los medios Bradyfact y Genisteína, no evidenció diferencias significativas entre estos tratamientos (Fig. 4). Sin embargo, la actividad de esta enzima en los nódulos de las plantas de soya con inóculo sin inductores de los genes nod (LM), sí mostró un comportamiento diferenciado al resto de las plantas inoculadas.
En condiciones de déficit hídrico, la actividad específica de la enzima GS1 en los nódulos de las plantas de soya inoculadas con la cepa B. elkanii ICA 8001 cultivada en el medio LM, mostró un incremento significativo con respecto al resto de los tratamientos. En contraste, la actividad de esta enzima en los nódulos de estas plantas, pero sin ser sometidas a estrés, fue menor estadísticamente en relación a su actividad en los demás tratamientos.
Los resultados correspondientes a la actividad enzimática GS1, para el grupo de plantas con déficit hídrico y con inóculo sin inductores de los genes nod (LM), se relacionan con los presentados por Clement y colaboradores (2008). Estos autores han demostrado una elevada acumulación del ARNm que codifica para la isoforma GS1 en nódulos de plantas de soya bajo condiciones de estrés osmótico, confirmando la presencia de una regulación positiva sobre el gen que codifica esta enzima bajo estas condiciones de estrés.
El hecho de que la actividad específica de la enzima GS1 no varíe con la imposición de un déficit hídrico en las plantas de soya previamente inoculadas con la cepa B. elkanii ICA 8001, cultivada en los medios Bradyfact y Genisteína, parece indicar que dicha enzima no se afecta con el estrés impuesto (Fig. 4). Sin embargo, en las plantas de soya inoculadas con esta cepa, cultivada en el medio LM, se detectó un mayor nivel de actividad de esta enzima a los 20 días de impuesto el déficit hídrico.
]]>
DISCUSIÓN
La disminución del CRA en las plantas de soya con déficit hídrico, puede deberse a una pérdida de vapor de agua a través de los estomas, influenciada por una disminución en la absorción radical de agua (25, 26). Esta afectación en la reposición del agua se debe al tratamiento con PEG 6000, que provocó un déficit hídrico en las plantas (Fig. 1).
Los resultados presentados sobre el comportamiento del CRA en las plantas de soya sometidas a estrés por déficit hídrico, muestran un efecto anti estrés en las plantas inoculadas con la cepa B. elkanii ICA 8001 previamente inducida en la producción de factores Nod (Bradyfact y Genisteína) (Fig. 1). Estos resultados evidenciaron un efecto positivo de la inducción de los genes nod en B. elkanii ICA 8001 sobre los mecanismos de tolerancia de la planta, que permitieron contrarrestar el déficit hídrico.
La cepa B. elkanii ICA 8001, cultivada en los medios Bradyfact y Genisteína, puede incrementar los niveles de secreción de los factores Nod (15, 27), lo que ha sido demostrado en otras cepas multiplicadas en medios de cultivo con inductores de los genes nod (Subramanian y cols., 2006). En este sentido, los factores Nod pudieran actuar como moléculas señales que participen en la síntesis de ácido abscísico (ABA) en las raíces de estas plantas (12, 28), de esta forma pudieran participar indirectamente en el cierre estomático (10), lo cual garantizaría una mayor retención de agua bajo condiciones de estrés por sequía. Por tanto, estos aspectos pudieran justificar que, bajo condiciones de déficit hídrico, las plantas de soya con inoculantes inducidos en la producción de factores Nod muestran valores del CRA superiores a los de plantas con inoculantes sin inducir y a los de plantas sin inocular (Fig. 1).
Los productos finales de la FBN en plantas de soya constituyen compuestos nitrogenados denominados ureidos, que se transportan desde el nódulo hasta las hojas para su posterior degradación (29). Mientras la FBN disminuye en respuesta al déficit hídrico, ocurre una acumulación de ureidos en hojas debido a una reducción de su catabolismo (30). King y Purcell (2005) han sugerido que la inhibición de la enzima nitrogenasa bajo condiciones de deficiencia hídrica está relacionada con un mecanismo de retroalimentación negativa, donde elevados niveles de ureidos en hojas o en nódulos constituyen los efectores fundamentales.
En las hojas de las plantas de soya con estrés e inoculadas con la cepa B. elkanii ICA 8001 sin inducir (LM), la acumulación de ureidos (Fig. 2) pudiera estar desencadenando un mecanismo de retroalimentación negativa sobre la FBN a medida que se incrementa el déficit hídrico en estas plantas (Fig. 1). Este posible mecanismo pudiera explicarse por el incremento de los niveles de ureidos nodular en este grupo de plantas (Fig. 3), donde estos compuestos nitrogenados pueden estar movilizándose desde las hojas hasta el nódulo para ejecutar su función inhibitoria sobre la FBN.
Ladrera y colaboradores (2007), han demostrado una disminución en los niveles de ureidos en hojas de plantas de soya tolerantes a la sequía cuando son sometidas a este estrés. Sin embargo, las plantas de soya sensibles al déficit hídrico presentan un incremento en los niveles de estos compuestos nitrogenados en hojas (9). Aunque la posible relación entre los niveles de ureidos foliar y nodular con los factores de la nodulación no se ha esclarecido completamente, es posible que a través del efecto anti estrés que estos oligosacáridos presentan (10), se pueda reducir la acumulación de ureidos en plantas de soya sometidas a deficiencia hídrica.
]]> En este trabajo, tanto en las hojas como en los nódulos de las plantas de soya con déficit hídrico e inoculadas con la cepa cultivada en los medios Bradyfact y Genisteína, los niveles de ureidos fueron inferiores a los de plantas inoculadas con la cepa multiplicada en el medio LM a los 12, 16 y 20 días de impuesto el estrés (Figs. 2B y 3B). En esta comparación se puede relacionar la influencia de la inoculación con la cepa B. elkanii ICA 8001 inducida en la síntesis de factores Nod, sobre los niveles de ureidos en las plantas de soya sometidas a estrés por sequía. Estos resultados permitieron inferir que la inducción de la síntesis de los factores Nod puede estar contrarrestando la acumulación de ureidos tanto en hojas como en nódulos. De alguna forma, la inducción de los genes de la nodulación en la cepa B. elkanii ICA 8001, pudiera estar actuando indirectamente sobre el catabolismo de los ureidos en las hojas.Las rutas de degradación de ureidos en las hojas involucran la enzima alantoato amidohidrolasa que requiere manganeso como cofactor (31). Purcell y colaboradores (2004) demostraron que las plantas de soya tolerantes a la sequía son capaces de absorber mayores niveles de manganeso que plantas sensibles a este estrés. Adicionalmente, los autores demostraron que, el incremento de manganeso en las plantas de soya sensibles y tolerantes a la sequía puede disminuir los niveles de ureidos en hojas bajo condiciones de estrés por déficit hídrico (32). Otras líneas de evidencias demuestran que el suplemento de manganeso bajo condiciones moderadas de déficit hídrico permite mejorar la FBN en plantas de soya sensibles al estrés hídrico por defecto (30).
Dado que la bacteria permanece en el nódulo radicular, es poco probable que exista una relación directa entre la inducción de la síntesis de factores Nod con el catabolismo de los ureidos en las hojas. Sin embargo, es posible pensar que la inducción de los genes nod en B. elkanii ICA 8001 así como la producción de los factores Nod por esta cepa, pudieran estar actuando positivamente sobre la absorción de manganeso en las plantas de soya sometidas a deficiencia hídrica. De hecho, existen reportes sobre el efecto de estos lipoquitooligosacáridos en incrementar el flujo de elementos como el calcio en las raíces de las plantas de soya durante las primeras etapas de la simbiosis (33).
De esta forma, las rutas de degradación de los ureidos pueden mantener una actividad metabólica que impida la acumulación de estos compuestos nitrogenados y la consiguiente inhibición de la FBN. Por tanto, bajo condiciones de déficit hídrico, la inhibición de la FBN en plantas de soya puede estar relacionada con el incremento de los niveles de ureidos en hojas y/o en nódulos, mientras que la disminución de estos compuestos nitrogenados se pudiera garantizar con la capacidad de la planta de incorporar mayores niveles de manganeso, a través de la absorción de nutrientes por la raíz.
Los controles de plantas sin inocular, sometidas o no al déficit hídrico, también mostraron niveles de ureidos en hojas, pero inferiores a los del resto de las plantas inoculadas. Este resultado permitió corroborar que los ureidos en las hojas no provienen únicamente de las rutas de asimilación del amonio simbiótico, sino también pueden resultar del catabolismo de las bases purínicas que normalmente ocurre en tejidos vegetales (34). No obstante, sí se observa un incremento de los niveles de ureidos foliar en las plantas de soya sin inocular y sometidas al déficit hídrico (Fig. 2), lo cual pudiera sugerir que las rutas de degradación de los ureidos también pueden bloquearse bajo condiciones de sequía en plantas de soya que no hayan desarrollado la FBN. Estas consecuencias validan la importancia de utilizar inoculantes inducidos en la producción de factores Nod para el cultivo de la soya.
Bajo condiciones de sequía, pueden acumularse ureidos en las plantas de soya afectando la FBN, por tanto, al lograr disminuir el efecto inhibitorio de estos compuestos nitrogenados sobre la FBN, es posible establecer un indicativo de tolerancia a la sequía en plantas de soya sometidas a este estrés.
La asimilación del amonio resultante de la FBN depende de una eficiente actividad de la enzima glutamina sintetasa (GS1) (24). En plantas de soya, la isoforma GS1 se expresa en la región infectada del nódulo y está directamente involucrada en la asimilación del amonio simbiótico (35). En este trabajo, la mayor actividad GS1 se obtuvo en plantas de soya inoculadas con la cepa B. elkanii ICA 8001 cultivada en el medio LM y sometidas a estrés hídrico por defecto (Fig. 4).
Por tanto, si la enzima GS1 presenta una mayor actividad en las plantas de soya inoculadas con la cepa sin inducir y sometidas a estrés por sequía, entonces la producción de glutamina pudiera estar incrementada en los nódulos de este grupo de plantas. El posible incremento en los niveles de glutamina bajo condiciones de déficit hídrico, puede estar en consonancia con el incremento en los niveles de ureidos foliar y nodular. Más aún, este incremento en los niveles de ureidos puede acentuarse si las rutas de degradación en las hojas se encuentran inhibidas como consecuencia del estrés hídrico impuesto.
]]> En contraste, las plantas inoculadas con la cepa multiplicada en los medios Bradyfact y Genisteína, sometidas o no al déficit hídrico, pudieran no tener variaciones en los niveles de producción de glutamina. Esta deducción se evidencia porque la actividad de la enzima GS1 no mostró diferencias significativas entre estos grupos de tratamientos (Fig. 4). Existen resultados contradictorios relacionados con la regulación de la isoforma GS1 en respuesta a la sequía. Estudios previos en nódulos de soya mostraron que el estrés oxidativo no afecta la enzima GS1 a niveles transcripcionales (36). Sin embargo, otras investigaciones han demostrado una regulación positiva sobre el gen que codifica la isoforma GS1 en nódulos de plantas de soya sometidas a estrés osmótico, así como un incremento de su ARNm sin provocar variaciones en la actividad catalítica de esta enzima (24, 37).El incremento en la actividad de la enzima GS1 en nódulos de las plantas de soya inoculadas con la cepa sin inducir (LM) y sometidas a un déficit hídrico, puede estar relacionado con una regulación positiva sobre el gen que codifica esta enzima (Fig. 4). En contraste, esta regulación génica puede estar disminuida en plantas con inoculantes inducidos en la producción de factores Nod (Bradyfact y Genisteína). Con la síntesis de estos lipoquitooligosacáridos, se pudiera estar regulando la expresión de la enzima GS1, de manera que sus niveles y/o su actividad catalítica no tengan incrementos bajo condiciones de deficiencia hídrica y, por tanto, constituya otro posible mecanismo que permita regular los niveles de ureidos bajo estas condiciones de estrés. Esta posible relación puede justificar las similitudes de la actividad enzimática GS1 en nódulos de plantas de soya con y sin déficit hídrico y con inoculantes inducidos en la producción de factores Nod (Bradyfact y Genisteína).
Este resultado es contradictorio con los presentados por Gordon y colaboradores (1997), quienes demostraron que los niveles de transcripción del gen que codifica la isoforma GS1 en plantas de soya bajo condiciones de déficit hídrico no fueron marcadamente afectados. Las divergencias presentadas pueden ser explicadas por los diferentes grados de susceptibilidad al estrés por sequía que pueden presentar las plantas utilizadas en estas investigaciones (24).
REFERENCIAS
1. Gage, D.J. Infection and invasion of roots by symbiotic nitrogen-fixing rhizobia during nodulation of temperate legumes. Microbiol. Mol. Biol. Rev., 2004, vol. 68, p. 280-300.
2. Morgan, J.; Bending, G. y White, P. Biological costs and benefits to plant–microbe interactions in the rhizosphere. J. Exp. Bot., 2005, vol. 56, p. 1729-1739.
]]>
3. Subramanian, S.; Stacey, G. y Yu, O. Endogenous isoflavones are essential for the establishment of symbiosis between soybean and Bradyrhizobium japonicum. The Plant Journal, 2006, vol. 48, p. 261-273.
4. Supanjani, S.; Habib, A.; Mabood, F.; Lee, K.D.; Donnelly, D. y Smith, D.L. Nod factor enhances calcium uptake by soybean. Plant Physiology and Biochemistry, 2006, vol. 44, p. 866-872.
5. Schubert, K.R. Products of biological nitrogen-fixation in higher plants: synthesis, transport, and metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol., 1986, vol. 37, p. 539-574.
6. Werner, A.K.; Sparkes, I.A.; Romeis, T. y Witte, C-P. Identification, biochemical characterization, and subcellular localization of allantoate amidohydrolases from Arabidopsis and soybean. Plant Physiol., 2008, vol. 146, p. 418-430.
7. King, C.A. y Purcell, L.C. Inhibition of N2 fixation in soybean is associated with elevated ureides and amino acids. Plant Physiol., 2005, vol. 137, p. 1389-1396.
]]>
8. Marino, D.; Frendo, P.; Ladrera, R.; Zabalza, A.; Puppo, A.; Arrese-Igor, C. y González, E.M. Nitrogen fixation control under drought stress: localized or systemic?. Plant Physiol., 2007, vol. 143, p. 1968-1974.
9. Ladrera, R.; Marino, D.; Estíbaliz, L.; González, E.M. y Arrese-Igor, C. Reduced carbon availability to bacteroids and elevated ureides in nodules, but not in shoots, are involved in the nitrogen fixation response to early drought in soybean. Plant Physiol., 2007, vol. 145, p. 539-546.
10. Atti, S.; Bonnell, R.; Prasher, S. y Smith, D.L. Response of soybean {Glycine max (L.) Merr.} under chronic water deficit to LCO application during flowering and pod filling. Irrig. Drain, 2005, vol. 54, p. 15-30.
11. Nápoles, M.C.; Guevara, E.; Montero, F.; Rossi, A. y Ferreira, A. Role of Bradyrhizobium japonicum induced by genistein on soybean stressed by water deficit. Spanish Journal of Agricultural Research, 2009, vol. 7, no. 3, p. 665-671.
12. Vendruscolo, E.C.G.; Schuster, J.; Pileggi, M.; Scapim, C.A.; Molinari, H.B.C.; Marur, C.J.; y Vieira, L.G.E. Stress-induced synthesis of praline confers tolerance to water deficit in transgenic wheat. J. Plant Physiol., 2007, vol. 164, no. 10, p. 1367-1376.
]]>
13. Duzan, H.M.; Zhou, A.; Souleimanov, A.; y Smith, D.L. Perception of Bradyrhizobium japonicum Nod factor by soybean [Glycine max (L.) Merr.] root hairs under abiotic stress conditions. Journal of Experimental Botany, 2004, vol. 55, no. 408, p. 2641-2646.
14. Vincent J.M. A manual for the practical study of root nodule bacteria. Oxford, Blackwell, Oxford, 1970.
15. Ponce, M.; Fé, C de la.; Ortiz, R. y Moya, C. INCAsoy-24 e INCAsoy-27: Nuevas variedades de soya para las condiciones climáticas de Cuba. Cultivos Tropicales, 2006, vol. 24, no. 3, p. 49.
Recibido 10/09/2010, aceptado 17/06/2011.
]]>{kind=link}
{kind=link}