ARTÍCULO ORIGINAL
Efecto de la competición larval sobre poblaciones del vector del dengue colectadas en Medellín, Colombia
Effect of larval competition on populations of the dengue vector collected in Medellín, Colombia
]]>
Dra. Beatriz Elena Giraldo Valencia, Dra. Marlen Martínez Gutiérrez
Universidad de Antioquia. Medellín, Colombia.
Introducción: Aedes aegypti es el vector responsable de transmitir el virus dengue, la densidad de la población en los estadios larvales puede afectar las características de vida del adulto, con implicaciones en la capacidad de transmisión viral.
Objetivo: determinar el efecto de la competición larval causada por la alta densidad en colonias de Aedes aegypti procedentes de zonas de alta y baja incidencia de dengue en la ciudad de Medellín, Colombia, así como en una colonia de referencia en el laboratorio.
Métodos: en las 3 colonias se evaluó el tiempo de desarrollo y supervivencia larval, así como el tamaño y el tiempo de supervivencia de los adultos.
Resultados: se encontró que la competición larval en condiciones de alta densidad redujo de manera significativa el tiempo de desarrollo y la supervivencia de los últimos estadios larvales. Por otro lado, la alta densidad experimentada durante la etapa larval, también generó cambios en los adultos de las 3 colonias, representados en una disminución significativa del tamaño y una disminución en el tiempo de supervivencia (esta última solo en las colonias de campo).
Conclusión: a pesar de que no se obtuvieron diferencias frente al efecto competitivo entre las colonias de campo provenientes de zonas con desigualdades en la incidencia de dengue en Medellín, la diferencia en respuesta obtenida en la colonia de laboratorio en comparación con las colonias de campo, hace pensar que es importante la evaluación de parámetros ecológicos sobre poblaciones de Aedes aegypti silvestres, porque estas evaluaciones acercarían más al comportamiento del vector. Estudios en curso en el laboratorio permitirán conocer si las diferencias encontradas entre las colonias afectan la competencia de Aedes aegypti frente a la infección por virus dengue. ]]>
Palabras clave: Aedes aegypti, virus dengue, competición larval, supervivencia vectorial, Colombia.
Introduction: Aedes aegypti is the vector transmitting the dengue virus. Larval density may affect adult life, with an impact on viral transmission capacity.
Objective: determine the effect of larval competition caused by high density in Aedes aegypti colonies from areas of high and low dengue incidence in the city of Medellín, Colombia, and in a reference colony in the laboratory.
Methods: the three colonies were evaluated for larval development and survival time, as well as size and survival time in adults.
Results: it was found that in high density conditions larval competition significantly shortened development and survival time in the last larval stages. High density during the larval stage also brought about changes in adults from the three colonies, represented by a significantly reduced size and a decrease in survival time (the latter only in the field colonies).
Conclusion: no differences in competitive effect were found between the field colonies from areas with unequal dengue incidence in Medellín. However, the difference in the response obtained from the laboratory colony in contrast to the field colony, points to the need to evaluate the ecological parameters of wild Aedes aegypti populations to obtain a more accurate view of the behavior of the vector. Laboratory studies in progress will make it possible to know whether the differences found between the colonies affect the competence of Aedes aegypti for dengue virus infection.
Key words: Aedes aegypti, dengue virus, larval competition, vector survival, Colombia.
]]>
INTRODUCCIÓN
El mosquito Aedes (Stegomyia) aegypti (subfamilia Culicinae) es el vector transmisor del virus dengue (DENV), virus responsable de la arbovirosis más importante a nivel mundial.1 El dengue puede presentarse con un cuadro clínico indiferenciado asintomático o como una enfermedad de variada intensidad, la cual puede estar acompañada de síntomas leves (dengue) o graves caracterizados por afectación de órganos, hemorragia severa e incremento de la permeabilidad vascular, que en última instancia pueden conducir a la muerte (dengue grave).2 En Colombia, Aedes aegypti se encuentra distribuido en cerca de 80 % del territorio nacional, en zonas ubicadas entre los 1 000 y 2 000 m sobre el nivel del mar. Esta amplia distribución, asociada a que el dengue es endémico en Colombia, son factores de alto riesgo para la transmisión del virus, agravando la problemática en salud pública en el país.
Ae. aegypti tiene 4 etapas en su ciclo de vida: huevo, larva, pupa y adulto. Las 3 primeras etapas se presentan en una fase acuática que inicia con el desarrollo embrionario, el cual se completa alrededor de las 48 h bajo condiciones óptimas de humedad y temperatura. Luego de este tiempo los huevos eclosionan, dando paso a la salida de la larva de primer estadio o instar. La duración de la etapa larval, la cual incluye 4 instar, depende de algunos factores ambientales como la temperatura, la disponibilidad de alimento y la densidad larval. Bajo condiciones óptimas, el tiempo de desarrollo larval es de aproximadamente 5 días, pero en condiciones desfavorables puede duplicarse.3 La etapa de pupa dura alrededor de 2 días y en ella el individuo realiza su metamorfosis para emerger luego como adulto. Poco después de emerger las hembras pueden aparearse y cerca de 24 a 36 h pueden alimentarse de sangre, de la cual obtienen la proteína necesaria para la maduración de sus huevos.4 Estudios realizados con colonias de Ae. aegypti han demostrado que la competencia larval por alimento disminuye la tasa de fecundidad de las hembras y produce variaciones en la longitud de sus alas.5 Adicionalmente la interacción competitiva en etapas tempranas de desarrollo puede generar efectos indirectos en la población adulta al interactuar con otros organismos o agentes patógenos.
En ese contexto por ejemplo, se sabe que factores ambientales como la temperatura,6 la precipitación, la altitud, entre otros, pueden determinar de cierto modo el riesgo de transmisión de algunos virus;7 pero cambios en la ecología del vector relacionados con factores intrínsecos de la población, tales como la competición intraespecífica,8 pueden modificar componentes asociados con su capacidad vectorial,que son aquellos procesos que le permiten al virus completar su ciclo de replicación y diseminación dentro del mosquito para luego ser transmitido a un huésped susceptible.9 En el hábitat acuático donde se crían los mosquitos, es común la competición larval,10 la cual puede estar ligada en muchos casos a factores denso-dependientes que en última instancia pueden alterar algunas características de la historia de vida de estos.5 Dentro de esas características se encuentran el tiempo de desarrollo, el peso corporal en estado de pupa, la supervivencia, el tamaño de los adultos y la fecundidad, entre otros.11,12 Adicionalmente la competición larval puede ser capaz de generar variaciones en la respuesta inmune de los mosquitos,13 que podría alterar el comportamiento en la trasmisión viral. Por ejemplo, se ha demostrado que condiciones de alta densidad larval experimentada por mosquitos Ae. aegypti y Ae. albopictus en su medio acuático alteran la interacción de las hembras adulta con el virus Sindbis, resultando en un aumento en la susceptibilidad a la infección y diseminación de este arbovirus.14
La mayoría de los estudios enfocados en el conocimiento de la biología y ecología del vector, se realizan sobre colonias que han permanecido por largo tiempo dentro del laboratorio, las cuales no se comportan de manera igual que las colonias obtenidas de campo. Por ejemplo, recientemente hemos demostrado que colonias de Ae. aegypti aisladas en 2 regiones colombianas son más susceptibles a la infección por algunos serotipos del DENV que la colonia Rockefeller. Mientras que las colonias de campo muestran niveles de infección por DENV serotipo 1 de 2x106 copias genómicas virales/mL, la cepa Rockefeller solo muestra niveles de infección de 5 x105 copias genómicas virales/mL.15
Teniendo en cuenta que dentro del conocimiento de la ecología del vector, las interacciones competitivas que se presentan en etapas inmaduras del mosquito (etapa larval) pueden tener un impacto sobre la competencia vectorial y que, adicionalmente las colonias de campo tienen un comportamiento diferente frente a la infección comparado con la cepa de referencia Rockefeller, se planteó como objetivo de este estudio determinar el efecto de la competición larval causada por la alta densidad, sobre características de la historia de vida de mosquitos Ae. aegypti de colonias de alta y baja incidencia de dengue de la ciudad de Medellín (Colombia), así mismo sobre la colonia de laboratorio Rockefeller.
]]>
MÉTODOS
Colecta del material entomológico
El material entomológico se colectó en la ciudad de Medellín, Antioquia (localizada a 6°132´553" de latitud Norte y 75°342 053 de latitud Oeste, y su altura sobre el nivel del mar varía entre los 1 800 a 1 500 m). Previo a ello se definieron las zonas de muestreo teniendo en cuenta la incidencia de dengue (según el número de casos reportados de dengue y dengue grave con hospitalización y sin esta, en las primeras 24 semanas epidemiológicas de 2010, sobre el número de habitantes por barrio); las condiciones socioeconómicas (barrios que pertenecían al mismo estrato socio económico, favoreciendo la colección en el estrato socioeconómico bajo, porque las deficientes condiciones sanitarias influyen en un aumento de la disponibilidad de criaderos artificiales para el mosquito); la abundancia del vector (índice de Breteau alto), con el fin de aumentar la probabilidad de encontrar mosquitos en una mayor proporción. Basados en estos criterios se seleccionó el barrio Las violetas (88 casos de dengue en las primeras 24 semanas epidemiológicas de 2010) y los barrios El Progreso 2 y El Triunfo, en conjunto (8 casos de dengue en las primeras 24 semanas epidemiológicas de 2010). En los barrios seleccionados se realizó captura de estadios inmaduros, así como de mosquitos adultos para el establecimiento de las colonias en insectario.
Establecimiento de las colonias en el insectario
Las variables tiempo de desarrollo y supervivencia de las larvas, así como tamaño y tiempo de supervivencia en hembras adultas, fueron evaluadas en 3 colonias de Ae. aegypti: La colonia Rockefeller, utilizada como colonia control de laboratorio y 2 colonias establecidas a partir de material entomológico recolectado en 2 zonas de la ciudad de Medellín (Colombia) con alta y baja incidencia (9,41 y 0,89 casos/1 000 habitantes, respectivamente) como se explicó antes. Una vez recolectado todo el material entomológico se llevó al insectario del Programa de Estudio y Control de Enfermedades Tropicales-PECET de la Universidad de Antioquia en Medellín para su cría y mantenimiento. Cuando las colonias fueron establecidas, se procedió a hacer una verificación taxonómica hasta especie de individuos utilizando individuos hembras y las claves taxonómicas de Forattini.16 ]]>
Mantenimiento de las colonias en el laboratorio
Todas las colonias se mantuvieron a 28 °C ± 2° C, con 75 % ± 5 % de humedad relativa y expuestos a un fotoperíodo de 12 h de luz y 12 h de oscuridad. Las larvas se mantuvieron en bandejas plásticas con agua limpia y se alimentaron una vez al día con truchina estéril (alimento para peces pulverizado). Por otro lado, las pupas fueron transferidas a jaulas y una vez que los adultos emergieron, se alimentaron con solución azucarada 10 % ad libitum. Posterior a la oviposición, las hembras adultas recolectadas en el campo fueron procesadas para detección por reacción en cadena de la polimerasa (PCR-polymerase chain reaction) de genoma viral, para determinar que no se encontraran infectadas de manera natural con ninguno de los serotipos del DENV.17
Para evaluar el efecto de la competición larval causada por la alta densidad, se establecieron condiciones de alta y baja densidad larval desde el primer estadio larval (aproximadamente 12 h de edad). Las larvas se contabilizaron y colocaron en bandejas plásticas con 600 mL de agua limpia. Se tomó un número de 70 larvas para la condición de baja densidad larval (0,12 larvas/mL) y 140 larvas para la condición de alta densidad larval (0,23 larvas/mL). Se realizaron 2 experimentos, cada uno con 3 réplicas. Estas condiciones se establecieron según reportes previos sobre densidades larvales en ambientes naturales.13 La alimentación en la etapa larval para ambos tratamientos fue de 0,1 g de Truchina estéril cada día.
Evaluación del tiempo de desarrollo larval
El tiempo de desarrollo larval se definió como el número de días transcurridos entre la eclosión de los huevos y la pupación, el cual se tomó como una medida del tiempo en el cual las larvas demoran en pasar al estado de pupa. Para la determinación del tiempo de desarrollo de las larvas, cada una de las bandejas se examinó diariamente (desde el día 0 hasta el día 12) realizándose un registro del número de larvas presentes por bandeja y calculando un porcentaje diario de estas. Ese registro se llevó a cabo hasta que todas las larvas pasaron al estadio de pupa o hasta que las larvas presentes en las bandejas murieron, que correspondió al día 12.
]]>
Evaluación de la supervivencia en la etapa larval
En ambas condiciones de crecimiento se realizó un registro diario mediante un conteo de larvas vivas y de individuos que fueron pasando al estado de pupa; este experimento se llevó a cabo hasta que todos los individuos hubiesen pasado al estado de pupa o murieron. El porcentaje de la supervivencia se determinó como el número total de individuos que alcanzaron el estado de pupa (obtenido al final del experimento), de un total de individuos de primer estadio larval que estuvieron presentes en cada una de las bandejas al inicio del experimento.
Evaluación del tamaño de los adultos
Para determinar el tamaño de los mosquitos adultos se seleccionaron 40 hembras de aproximadamente 5 a 10 días de edad (por cada condición de densidad poblacional y tipo de colonia), las cuales fueron sacrificadas por medio de congelación durante 20 min, para después removerles el ala derecha, la cual fue medida como indicador del tamaño. La medida se hizo desde el álula hasta la margen apical (zona más alejada del surco axilar) del ala, sin considerar las escamas del borde, conforme a la metodología previamente descrita.18 La longitud de las alas fue tomada en mm y se realizó sobre una plantilla de hoja milimetrada con ayuda de un microscopio estéreo.
Evaluación del tiempo de supervivencia de hembras adultas
]]>
La evaluación del tiempo de supervivencia se realizó con las hembras adultas recuperadas del ensayo de competición larval. Las pupas de cada uno de los experimentos se retiraron a diario y posteriormente se colocaron en recipientes plásticos con alrededor de 40 mL de agua. Esos recipientes se introdujeron dentro de jaulas plásticas para cría de insectos hasta el momento de la emergencia de los adultos. Para la evaluación de supervivencia se seleccionaron, con ayuda de un aspirador manual, un total de 33 hembras (entre ±1 y 1 días de emergidas) de cada colonia, las cuales fueron alimentadas a diario con solución azucarada 10 %. Para determinar el tiempo de supervivencia se realizó un registro diario (desde el día 0 hasta el día 60) de las muertes en cada una de las jaulas. Este tiempo se determinó teniendo en cuenta que bajo condiciones de laboratorio Ae. aegypti vive un promedio de 55,6 días.19
Análisis estadístico
En este estudio se realizó comparación entre tratamientos y entre variables de estudio (colonias), los tratamientos correspondieron a alta y baja densidad larval; las variables de estudio fueron colonia baja incidencia, colonia alta incidencia y colonia de laboratorio (control); las variables respuesta resultaron tiempo de desarrollo larval, supervivencia larval, tamaño corporal y supervivencia de hembras adultas. Para evaluar las diferencias en el tiempo de desarrollo larval se evaluó primero la normalidad de los datos mediante la prueba Shapiro-Wilk. Se determinó la normalidad, entonces se utilizó la prueba t de Student para comparar las medias entre las condiciones de alta y baja densidad larval, cada uno de los días del tiempo de desarrollo. Para el caso donde la normalidad no fue asumida se utilizó en el análisis de las medias la prueba U de Mann-Whitney. Para evaluar las diferencias en el tiempo de desarrollo entre las 3 colonias dentro de una misma condición (alta o baja densidad larval) se realizó un ANOVA seguido de un análisis post-hoc de Tukey, si los datos se asumían dentro de una distribución normal. Para el caso donde no se asumió la normalidad de los datos se realizó la prueba H de Kruskal-Wallis. En todos los casos se consideraron diferencias significativas con un valor p< 0,05.
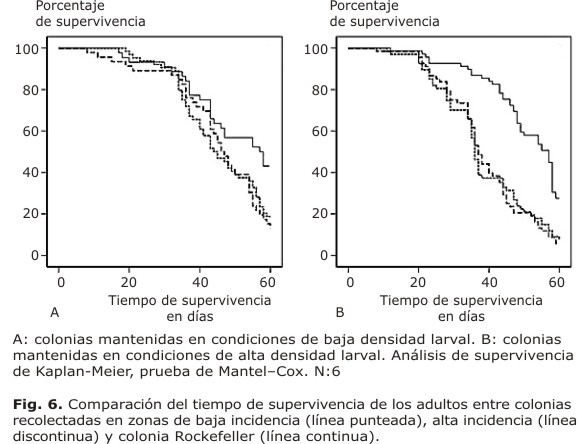
Para la supervivencia larval, se evaluó primero la normalidad de los datos mediante la prueba Shapiro-Wilk, comprobándose esta, se utilizó la prueba t de Student para analizar las diferencias entre los promedios de la supervivencia larval. Para determinar cambios en el tamaño corporal que pudieran ser influenciados por la alta densidad larval, se compararon los tamaños de las alas (entre las 3 colonias) con la prueba no paramétrica U de Mann-Whitney. Para el análisis de los promedios de la longitud de las alas de cada una de las colonias de campo, en comparación con la colonia control dentro de una misma condición (alta densidad o baja densidad larval), se utilizó la prueba no paramétrica U de Mann-Whitney, con p< 0,05. Finalmente, para determinar variaciones en la supervivencia de las hembras de Ae. aegypti se realizaron curvas de supervivencia mediante el estimador no paramétrico de Kaplan-Meier y, después se utilizó la prueba de Mantel-Cox,para evaluar el efecto de la densidad en la supervivencia, dentro de cada colonia y comparar si hubo diferencias significativas entre las colonias dentro de una misma condición.
RESULTADOS
]]>
Tiempo de desarrollo larval
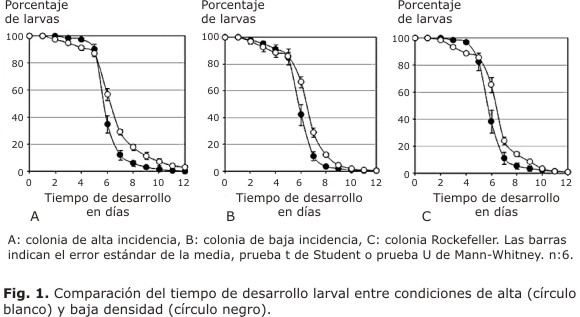
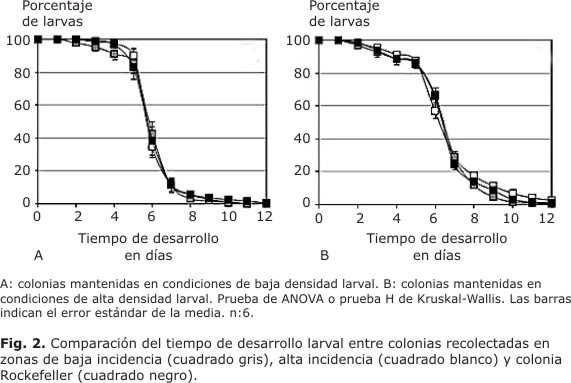
Cuando se analizó el tiempo de desarrollo larval en la colonia originada a partir de material recolectado en la zona de alta incidencia de dengue, se encontró un mayor porcentaje de larvas mantenidas en condiciones de alta densidad que en condiciones de baja densidad larval. Estas diferencias fueron significativas (p< 0,05) a los días 3, 4, 6, 7, 8 y 9 (Fig. 1A). Cuando se comparó el tiempo de desarrollo larval en la colonia, que fue originada a partir de material recolectado en zonas de baja incidencia para dengue se encontró, de igual manera, que el porcentaje de aquellas larvas que experimentaron condiciones de alta densidad resultó mayor (p< 0,05) en comparación con las larvas se criaron en condiciones de baja densidad larval a los 6, 7, 8 y 9 días (Fig. 1B). Cuando se evaluó el tiempo de desarrollo larval en la colonia de laboratorio Rockefeller, también se encontró que el porcentaje de larvas en las colonias conservadas en condiciones de alta densidad larval fue mayor con respecto a las larvas mantenidas en condiciones de baja densidad larval. Estas diferencias resultaron significativas (p< 0,05) a los días 3, 4, 6, 7 y 8 (Fig. 1C). Al comparar el tiempo de desarrollo larval entre la colonia Rockefeller y las colonias de alta y baja incidencia de dengue, en condiciones de alta o baja densidad larval, no se encontraron diferencias significativas (p> 0,05) en comparación con la cepa control Rockefeller (Fig. 2A y 2B).
]]>
Supervivencia larval
Al comparar el porcentaje promedio de supervivencia de las larvas en las 3 colonias (Control Rockefeller, alta incidencia y baja incidencia), se encontró que fue más bajo (p< 0,05) en las colonias mantenidas en alta densidad larval en comparación con las colonias conservadas en condiciones de baja densidad larval. Los porcentajes se muestran en la tabla.
Tamaño de los adultos
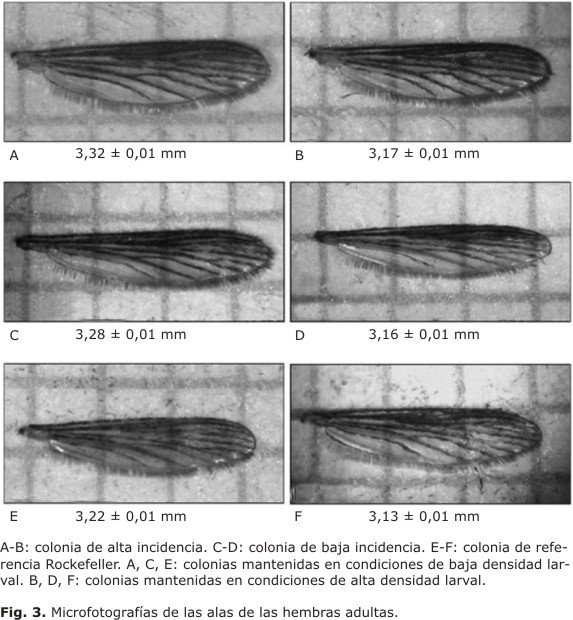
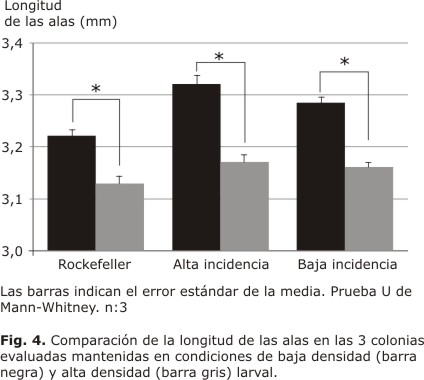
Cuando se comparó el promedio del tamaño de las alas de los mosquitos de la colonia de alta incidencia (Fig. 3A y 3B) entre las 2 densidades larvales, se encontraron diferencias significativas (p< 0,05) en su longitud, siendo mayor el tamaño de las alas en los mosquitos que fueron criados en condiciones de baja densidad larval (3,32 ± 0,01 mm de longitud) en comparación con los mosquitos criados en condiciones de alta densidad larval (3,17 ± 0,01 mm de longitud) (Fig. 4). Para la colonia de baja incidencia (Fig. 3C y 3D), el promedio del tamaño de las alas de los mosquitos que fueron criados en condiciones de baja y alta densidad larval fue de 3,28 ± 0,01 y 3,16 ± 0,01 mm, respectivamente. Cuando se comparó el promedio de la longitud de las alas para esta colonia, cuyos individuos en su etapa inmadura se mantuvieron en baja densidad y alta densidad larval, el resultado indicó diferencias significativas (p< 0,05) (Fig. 4). Se encontró que los mosquitos criados a baja densidad larval tenían un tamaño mayor de alas con respecto a los mosquitos criados a alta densidad larval. El promedio del tamaño de las alas para los mosquitos de la colonia Rockefeller, que fueron criados a baja densidad larval resultó de 3,22 ± 0,01 mm y para los criados a alta densidad 3,13 ± 0,01 mm (Fig. 3E y 3F). Cuando se compararon estos promedios el resultado mostró diferencias significativas (p< 0,05), observándose que los mosquitos con baja densidad larval en su etapa inmadura obtuvieron un tamaño significativamente mayor en comparación con los que experimentaron crecimiento en alta densidad larval (Fig.4).
Finalmente, el tamaño de las alas entre cada una de las 2 colonias de campo y la colonia Rockefeller (que funcionó como control), también fue comparado en cada una de las condiciones de densidad larval. En condiciones de baja densidad larval tanto la colonia de baja incidencia como la colonia de alta incidencia mostraron un mayor tamaño de alas (3,32 ± 0,01 mm de longitud y 3,28 ± 0,01 mm de longitud, respectivamente) con respecto al tamaño de la colonia control Rockefeller (3,22 ± 0,01 mm de longitud). Al comparar las medias entre las colonias de campo con la colonia control que fueron mantenidas en condiciones de baja densidad larval, se obtuvieron diferencias significativas entre la colonia de campo de alta incidencia o la de baja incidencia en comparación con el control (p<0.05) (Fig. 4).
Al comparar las medias entre las colonias de alta y baja incidencia con el Control, las cuales fueron mantenidas en condiciones de alta densidad larval, solo se encontraron diferencias significativas (p< 0,05) entre la colonia de alta incidencia y el control (3,17 ± 0,01 mm de longitud y 3,13 ± 0,01 mm de longitud; respectivamente), siendo mayor el tamaño de las alas en la colonia de alta incidencia. No se encontraron diferencias significativas entre el tamaño de las alas de la colonias de baja incidencia vs. Control (p> 0,05) (Fig. 4).
Supervivencia de las hembras adultas
Al comparar el tiempo de supervivencia entre las 2 condiciones de densidad larval para la colonia de alta incidencia, se encontró que el tiempo de supervivencia en la vida adulta se disminuyó de manera significativa (p< 0,05) cuando los mosquitos en su etapa inmadura experimentaron alta densidad larval (Fig. 5A). Cuando se evaluó la supervivencia de las hembras adultas de la colonia de campo baja incidencia, que estuvieron expuestas a condiciones de baja densidad y alta densidad larval, se encontraron diferencias significativas (p< 0,05) entre las 2 densidades. Aquellas hembras que experimentaron alta densidad de larvas en su etapa inmadura presentaron menor supervivencia que aquellas que se criaron a bajas densidades larvales (Fig. 5B). Al comparar la supervivencia de las hembras de la colonia Rockefeller que fueron criadas en condiciones de baja densidad larval con respecto aquellas que se criaron a alta densidad larval, el resultado indicó que no existen diferencias significativas entre estas 2 condiciones (p> 0,05) y que la supervivencia de los mosquitos de la colonia Control no se vio afectada por el efecto de la densidad larval (Fig. 5C).
]]>
Finalmente, al comparar el tiempo de supervivencia de las colonias de alta y baja incidencia de dengue en relación con la cepa Rockefeller, se encontraron diferencias significativas (p< 0,05), tanto en las colonias mantenidas a baja densidad larval (Fig. 6A) como en las colonias mantenidas en condiciones de alta densidad larval (Fig. 6B).
DISCUSIÓN
A pesar de que Colombia es un país endémico para el dengue y que ciudades como Medellín presentan un gran número de casos por año, las interacciones competitivas que se presentan en la etapa larvaria de Ae. aegypti no han sido estudiadas en el país. Adicionalmente, a nivel mundial, son pocos los estudios que se han realizado para evaluar la ecología del vector en poblaciones nativas. Por esa razón, este estudio se enfocó en evaluar el efecto de la competición larval causada por la alta densidad, sobre 4 características de vida del vector: 2 en etapa larvaria (tiempo de desarrollo y supervivencia) y 2 en etapa adulta (tamaño y tiempo de supervivencia) en 2 colonias de campo recolectadas en la ciudad de Medellín y en la cepa de referencia Rockefeller.
Los resultados del presente estudio demostraron que el tiempo de desarrollo larval es altamente sensible a la condición de alta densidad, experimentada por los individuos en su hábitat durante los estadios larvales, porque tanto las colonias de campo (alta y baja incidencia), como la colonia Control (Rockefeller) exhibieron una prolongación en el tiempo de desarrollo, que fue mucho más evidente en los últimos estadios larvales que correspondieron a los días 6, 7, 8 y 9. Estos resultados concuerdan con lo reportado previamente en donde se ha observado un aumento en la prolongación del tiempo de desarrollo larval en condiciones de crecimiento de alta densidad.20-22 Este retraso en el tiempo de desarrollo de las larvas mantenidas en alta densidad puede ser debido a la competencia por alimento, como ha sido postulado antes.23 Adicionalmente, un retraso en el tiempo de desarrollo dependiente de las reservas nutricionales tiene un efecto negativo sobre los últimos estadios larvales, los cuales son claves para la larva, porque esta debe obtener una masa corporal crítica que le permita pasar al estado de pupa. Es por ello que el alimento debe estar presente después de que las larvas alcanzan este punto en su masa crítica, porque si este se ve disminuido o no está disponible, las larvas llegan a la etapa de metamorfosis con un tamaño más reducido.24 Aparte de la competencia por alimento, otros factores dependientes de la densidad han sido considerados por otros autores como responsables de tener un impacto negativo en el desarrollo larvario. Por ejemplo, una mayor cantidad de individuos en el medio acuático se traduce en un aumento de materiales de desecho, como son las larvas muertas en descomposición, exuvias y productos de excreción de las larvas como nitrógenos en forma de amoniaco, que pueden ser responsables del retraso en el crecimiento larval.3 El tiempo de desarrollo larval entre las 3 colonias alta incidencia, baja incidencia y Control bajo una misma densidad también fue comparado, encontrándose que aquellas colonias crecidas a bajas densidades larvales presentaron un tiempo de desarrollo muy similar entre sí y eso estuvo acorde con lo reportado en la literatura para larvas criadas bajo condiciones óptimas; donde las larvas se desarrollan a pupa en 8 días o menos.4 Para las larvas que experimentaron alta densidad larval, el tiempo de desarrollo desde primer estadio hasta su estado de pupa presentó un retraso donde los últimos individuos (larvas) puparon hasta el día 11 y 12 del experimento; este retraso en el tiempo tuvo similar comportamiento en las 3 colonias evaluadas, no mostrándose diferencias significativas al hacer la comparación entre estas.
Además del retraso en el tiempo de desarrollo exhibido por las larvas en condiciones de alta densidad, otra característica que se vio afectada por este factor fue la supervivencia de las larvas. Se pudo observar una significativa disminución en la supervivencia de las larvas que fueron mantenidas bajo condiciones de alta densidad, tanto para las colonias de campo como para la colonia control. Sin embargo, la colonia de alta incidencia de dengue fue la que presentó mayor tasa de mortalidad bajo esta condición, donde solo 78 % de las larvas alcanzaron el estado de pupa. La disminución en la tasa de supervivencia de los individuos en el medio ambiente larval bajo condiciones de alta densidad, podría estar asociada con un comportamiento de tipo caníbal entre las larvas de Ae. aegypti. Aunque el canibalismo no es de esperarse en larvas que son filtradoras de detritus como ocurre en especies de Aedes, algunos estudios sobre larvas de Ae. triseriatus han mostrado el canibalismo entre esta especie y su vulnerabilidad, la cual se hace presente en los 4 estadios larvales.25 De igual manera, otros estudios han demostrado que cuando se presenta baja disponibilidad de alimento en el ambiente larval, existe una considerable pérdida de individuos, caso que lo han atribuido al canibalismo presentado entre las larvas.26 ]]>
En la etapa adulta, el tamaño corporal de las hembras se vio afectado por el aumento de la densidad poblacional. En este estudio, ese tamaño fue determinado sobre la base de la longitud del ala de las hembras, debido a que el tamaño del mosquito no puede tomarse sobre una medida directa de su cuerpo por la forma acorazonada que presenta su abdomen, de igual manera el peso seco del mosquito tampoco es considerado como buen estimador de su tamaño corporal porque se podrían presentar diferencias en dependencia de si la hembra está o no alimentada, o presenta o no gravidez.27 Por lo tanto, se optó por emplear la medida del ala, como un estimador de tamaño porque permite hacer una medición lineal en los insectos.28 Se sabe que el tamaño corporal de los mosquitos tiene una base genética, pero modificaciones en su hábitat larval determinadas por factores ecológicos, como por ejemplo la alta densidad poblacional, pueden generar variaciones en su tamaño.11 Estudios realizados en especies de mosquitos vectores han registrado cómo el incremento en la densidad larval tiene un efecto en el tamaño corporal del mosquito en su etapa adulta, mostrando que a altas densidades, el mosquito disminuye significativamente su tamaño29 y que este en parte es influenciado por la disponibilidad de alimento que tienen los individuos en su etapa inmadura. Es así como los mosquitos Culex pipiens quinquefasciatus criados a bajas densidades tienen mayores reservas calóricas, siendo su tamaño mayor en la fase adulta que aquellos criados a altas densidades y con reservas deficientes.30.El tamaño de los mosquitos adultos también ha sido relacionado con otros componentes de la capacidad vectorial, como la supervivencia,11 la cual es considerada la variable más crítica en la estimación de la capacidad vectorial, debido a que la probabilidad de supervivencia diaria del mosquito esencialmente determina la duración de vida infectiva de un vector.31 A pesar de que la supervivencia del mosquito es dependiente en gran parte de factores como la temperatura y humedad, otros factores como las condiciones de cría en estadio larval pueden generar variabilidad en esta.30 Aquí se obtuvo una significativa disminución en la supervivencia de las hembras adultas cuyas larvas fueron criadas en condiciones de altas densidades larvales, en comparación con aquellas que experimentaron bajas densidades, donde cerca de 50 % de las hembras no sobrevivieron más de los 35 días, en contraste con la colonia Control donde la supervivencia bajo las mismas condiciones fue extremadamente amplia; cercana a los 55 días. Tal respuesta en la colonia de laboratorio podría pensarse que está relacionada con los procesos de cría establecidos por un largo período, como lo son la cantidad de larvas criadas por bandeja y los ciclos de alimentación; que en muchos casos varían en dependencia de los requerimientos de material (mosquitos) para ensayos, asímismo la calidad del alimento, entre otros, los cuales pudieron haber creado en estos individuos algún mecanismo que les permitiera contrarrestar los efectos negativos de la competición sobre su supervivencia. Tales resultados, en los cuales se refleja una diferencia significativa en la supervivencia de las colonias de campo con respecto a la colonia control en ambos tratamientos, podría ser un buen indicador de que estas colonias aún conservan muchas características de la población silvestre. Esto podría indicar que esos estudios hechos en laboratorio sobre colonias de campo pueden ser un buen referente, el cual puede acercarnos a respuestas y comportamientos que tendrían estas poblaciones en su entorno natural. De ahí la importancia de que factores ecológicos de este vector sean evaluados en colonias de campo.
Asímismo los hallazgos del presente estudio se podrían correlacionar con las posibles repercusiones sobre la capacidad vectorial de Ae. aegypti frente al DENV. Por ejemplo, una marcada disminución en el tamaño corporal de las hembras adultas (generado por el efecto de la competición bajo condiciones de alta densidad larval) podría estar modulando componentes tan importantes dentro de la capacidad vectorial como la tasa de picadura y dispersión. Varios estudios han demostrado que hembras más pequeñas tienden a alimentarse más frecuentemente de sangre, aparte de tener también una mejor dispersión que hembras de tamaño más grande,28,32 lo que promueve una mayor oportunidad de contacto del mosquito con un hospedero, hecho que podría potenciar la transmisión del virus. Por otro lado, otros estudios han demostrado que hembras de Ae. albopictus que experimentaron en su etapa larval efectos competitivos, disminuyen su tamaño y resultan más susceptibles de ser infectadas y de diseminar el DENV que hembras de mayor tamaño,33 hecho que podría contribuir a una mayor transmisión del virus.
En cuanto a la tasa de supervivencia de las hembras adultas la cual se vio disminuida para las colonias de campo que experimentaron altas densidades, asociar esta con el tamaño de los mosquitos, el cual disminuyó de una manera considerable bajo esta misma condición, puede ser de gran ayuda para entender cómo puede existir un sistema compensatorio entre el tamaño y la supervivencia. A pesar de que los mosquitos sobreviven menos tiempo, la capacidad de estos para alimentarse más a menudo de un hospedero34 y la habilidad que pueden tener para recorrer mayores distancias, lo cual puede estar determinado por su pequeño tamaño,28 lograría promover en esta clase de vectores un eficiente contacto con un hospedero. Este panorama, en última instancia, podría estar generando un aumento en los patrones de transmisión de DENV, creando así un mayor riesgo de infección para la población humana.
En conclusión, la competición presentada en el hábitat larval de Ae. aegypti tuvo un efecto negativo en cada una de las características de historia de vida evaluadas, el tiempo de desarrollo y la supervivencia de las larvas fue altamente alterado por este efecto, de igual manera las etapas posteriores a la etapa larval también presentaron alteraciones, donde el tamaño corporal de las hembras y su supervivencia se vieron disminuidas por la competición. Estos hechos indican que factores ecológicos intrínsecos de la población, como la alta densidad poblacional en la etapa inmadura, tienen un importante efecto en la alteración de características de historia de vida del mosquito Ae. aegypti y a su vez, estas alteraciones en las características de historia de vida de los mosquitos, que son importantes dentro de los componentes de su capacidad vectorial, podrían modular la epidemiología de enfermedades transmitidas por mosquitos. Estudios en curso permitirán evaluar la competencia intraespecífica en poblaciones de campo de Ae. aegyptiy las repercusiones que esta competición pueda tener sobre componentes intrínsecos del vector (la competencia vectorial) que puedan llevar a cambios en la dinámica de transmisión del DENV en Colombia.
Finalmente, a pesar de que no se obtuvieron diferencias frente al efecto competitivo entre las colonias de campo provenientes de zonas con diferencias en la incidencia de dengue en Medellín, el contraste en la respuesta obtenida en la colonia de laboratorio en comparación con las colonias de campo, hace pensar que es importante la evaluación de parámetros ecológicos sobre poblaciones de Ae. aegypti silvestres, porque estas acercan más a su comportamiento.
AGRADECIMIENTOS
]]>
A Alexander Darío Uribe Yepes por el establecimiento de la recolección del material entomológico y el de las colonias en el laboratorio. Este trabajo fue financiado por el Departamento Administrativo de Ciencia, Tecnología e Investigación- COLCIENCIAS. Proyecto 111549326083/2009.
REFERENCIAS BIBLIOGRÁFICAS
1. Jansen CC, Beebe NW. The dengue vector Aedes aegypti: what comes next. Microbes Infect. 2010;12(4):272-9.
2. Lin CY, Huang CH, Chen YH. Classification of dengue: the clinical use of World Health Organization 2009 guideline. J Formos Med Assoc. 2013 Feb;112(2):61-3.
3. Bargielowski I, Nimmo D, Alphey L, Koella JC. Comparison of life history characteristics of the genetically modified OX513A line and a wild type strain of Aedes aegypti. PLoS One. 2011;6(6):e20699.
4. Marquardt WC, Black W, Freier J, Hagedorn H, Moore C, Hemingway J, et al. Biology of Disease Vectors. Cap 8. 2da. ed. California (U.S.A.): Elsevier Academic Press; 2005.
5. Beserra EB, Fernandes CR, Ribeiro PS. Relação entre densidade larval e ciclo de vida, tamanho e fecundidade de Aedes (Stegomyia) aegypti (L.)(Diptera: Culicidae) em laboratório. Neotrop Entomol. 2009;38(6):847-52.
6. Westbrook CJ, Reiskind MH, Pesko KN, Greene KE, Lounibos LP. Larval environmental temperature and the susceptibility of Aedes albopictus Skuse (Diptera: Culicidae) to Chikungunya virus. Vector Borne Zoonotic Dis. 2010;10(3):241-7.
7. Halstead SB. Dengue virus-mosquito interactions. Annu Rev Entomol. 2008;53:273-91.
8. Alto BW, Reiskind MH, Lounibos LP. Size alters susceptibility of vectors to dengue virus infection and dissemination. Am J Trop Med Hyg. 2008;79(5):688.
9. Quintero Gil DC, Osorio Benítez JE, Martínez-Gutiérrez M. Vector competence: Entomological considerations and its implications on the epidemiology of dengue. Iatreia. 2010;23(2):137-45.
10. Reiskind M, Lounibos L. Effects of intraspecific larval competition on adult longevity in the mosquitoes Aedes aegypti and Aedes albopictus. Med Vet Entomol. 2009;23(1):62-8.
11. Gama RA, Alves KdC, Martins RF, Eiras ÁE, Resende MCd. Effect of larval density on adult size of Aedes aegypti reared under laboratory conditions. Rev Soc Bras Med Trop. 2005;38(1):64-6.
12. Walsh R, Facchinelli L, Ramsey J, Bond J, Gould F. Assessing the impact of density dependence in field populations of Aedes aegypti. J Vector Ecol. 2011;36(2):300-7.
13. Muturi EJ, Blackshear M, Montgomery A. Temperature and density dependent effects of larval environment on Aedes aegypti competence for an alphavirus. J Vector Ecol. 2012;37(1):154-61.
14. Alto BW, Lounibos LP, Higgs S, Juliano SA. Larval competition differentially affects arbovirus infection in Aedes mosquitoes. Ecology. 2005;86(12):3279-88.
15. Uribe A, Ospina M, Díaz F, Osorio J, Gutierrez. MM. Diferencias en la capacidad de replicación de las cepas del virus dengue, serotipos 1 y 3, aisladas en Medellín en poblaciones urbanas de Aedes aegypti recolectadas en la misma zona geográfica. Infectio. 2012;16(S1):106.
16. Forattini OP. Culicidologia médica: identificação, biologia, epidemiologia; Medical culicidology: ID, biology, epidemiology; São Paulo:; Editora da Universidade de São Paulo; 2002. p. 860.
17. Uribe A, Marín E, Quintero C, Arbelaez L, Martínez-Gutiérrez M. Evaluación de infección natural por virus dengue en mosquitos Aedes aegypti colectados en zonas de alto y bajo riesgo de la enfermedad en la ciudad de Medellín. Hechos Microb. 2011;6(S1):53.
18. Harbach RE, Knight KL. Taxonomists' glossary of mosquito anatomy. Marlton, New Jersey: Plexus Publishing Inc.; 1980. p. 415. ]]>
19. Briegel H, Kaiser C. Life-span of mosquitoes (Culicidae, Diptera) under laboratory conditions. Gerontology. 1973;19(4):240-9.
20. Reiskind MH, Walton ET, Wilson ML. Nutrient-dependent reduced growth and survival of larval Culex restuans (Diptera: Culicidae): laboratory and field experiments in Michigan. J Med Entomol. 2004;41(4):650-6.
21. Maciá A. Effects of larval crowding on development time, survival and weight at metamorphosis in Aedes aegypti (Diptera: Culicidae). Rev Soc Entomol Argent. 2009;68(1-2):107-14.
22. Mitchell Foster K, Ma B, Warsame Ali S, Logan C, Rau M, Lowenberger C. The influence of larval density, food stress, and parasitism on the bionomics of the dengue vector Aedes aegypti (Diptera: Culicidae): implications for integrated vector management. J Vector Ecol. 2012;37(1):221-9.
23. Arrivillaga J, Barrera R. Food as a limiting factor for Aedes aegypti in water-storage containers. J Vector Ecol. 2004;29:11-20. ]]>
24. Telang A, Frame L, Brown MR. Larval feeding duration affects ecdysteroid levels and nutritional reserves regulating pupal commitment in the yellow fever mosquito Aedes aegypti (Diptera: Culicidae). J Exp Biol. 2007;210(5):854-64.
25. Koenekoop RK, Livdahl TP. Cannibalism among Aedes triseriatus larvae. Ecol Entomol. 1986;11(1):111-4.
26. Agnew P, Koella JC. Life history interactions with environmental conditions in a hostparasite relationship and the parasite's mode of transmission. Evol Ecol. 1999;13(1):67-91.
27. Jirakanjanakit N, Leemingsawat S, Thongrungkiat S, Apiwathnasorn C, Singhaniyom S, Bellec C, et al. Influence of larval density or food variation on the geometry of the wing of Aedes (Stegomyia) aegypti. Trop Med Int Health. 2007;12(11):1354-60.
28. Maciel de Freitas R, Codeco C, Lourenço de Oliveira R. Body size associated survival and dispersal rates of Aedes aegypti in Rio de Janeiro. Med Vet Entomol. 2007;21(3):284-92. ]]>
29. Agnew P, Haussy C, Michalakis Y. Effects of density and larval competition on selected life history traits of Culex pipiens quinquefasciatus (Diptera: Culicidae). J Med Entomol. 2000;37(5):732-5.
30. Rebollar-Téllez EA, Magallón E, Solís-Franco RR. Efecto de hacinamiento larval sobre algunas características del adulto de Culex pipiens quin-quefasciatus (Diptera:Culicidae). Rev Biomédica. 1995;6:5-10.
31. Milby M, Reisen W. Estimation of vectorial capacity: vector survivorship. Bull Soc Vector Ecol. 1989;14(1):47-54.
32. Briegel H. Metabolic relationship between female body size, reserves, and fecundity of Aedes aegypti. J Insect Physiol. 1990;36(3):165-72.
33. Alto BW, Lounibos LP, Mores CN, Reiskind MH. Larval competition alters susceptibility of adult Aedes mosquitoes to dengue infection. Proc Biol Sci. 2008;275(1633):463-71. ]]>
34. Scott TW, Amerasinghe PH, Morrison AC, Lorenz LH, Clark GG, Strickman D, et al. Longitudinal studies of Aedes aegypti (Diptera:Culicidae) in Thailand and Puerto Rico: blood feeding frequency. J Med Entomol. 2000;37(1):89-101.
Recibido: 12 de julio de 2013.
Aprobado: 28 de octubre de 2013.
Marlen Martinez Gutiérrez. Programa de Estudio y Control de Enfermedades Tropicales-PECET. Sede de Investigación Universitaria, Universidad de Antioquia, Calle 62 #52-59 Lab. 632. Medellín, Colombia. Correo electrónico: mmartinezg@udea.edu.co ]]>

{kind=link}