
Patrón de la actividad de las β -1,3-glucanasas y quitinasas en la interacción HMA-sistemina en tomate. I. Fase presimbiótica
Pattern of β-1, 3-glucanase and chitinase activity in the AMF - systemin interaction in tomato. I. Presymbiotic phase
Dra.C. Blanca M. de la Noval-Pons,I Dra.C. Ondina León-Díaz,II M.Cs. Norma A. Martínez-Gallardo,III Dr.C. Eduardo Pérez-Ortega,I Dr.C. John P. Délano-FrierIII
IInstituto Nacional de Ciencias Agrícolas. Gaveta Postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700.
IICentro Nacional de Sanidad Agropecuaria (CENSA). Apartado 10. San José de las Lajas (La Habana). 32700 Cuba.
]]> IIIUnidad de Biotecnología e Ingeniería Genética de Plantas del Cinvestav-Campus Irapuato. Km 9,6. Libramiento Norte, Carr. Irapuato-León. A.P. 629, C.P. 36500, Irapuato, Gto. México.
RESUMEN
Los hongos micorrícicos arbusculares y los elicitores han sido ampliamente utilizados en la agricultura moderna con diversos objetivos. La sistemina constituye una de las hormonas polipeptídicas con amplias posibilidades como elicitor de respuestas de defensa en Solanáceas. Su aplicación de forma conjunta con los hongos micorrícicos arbusculares, permitiría incrementar los efectos en la protección contra patógenos diversos. Para determinar la participación de la sistemina en el proceso de establecimiento de los HMA en los estadios tempranos de la simbiosis micorrícica arbuscular y su influencia en la inducción de respuestas de defensa en tomate se realizó un experimento en fase presimbiótica. La sistemina aplicada en las raíces produce la inducción rápida de las β-1,3-glucanasas y quitinasas en etapas tempranas, de forma local y sistémica. Se observó un efecto sinérgico entre la sistemina y del HMA (F. mosseae), sobre la actividad de las β-1,3-glucanasas en raíz, desde la primera hora posterior a la aplicación de ambos. F. mosseae produjo la inducción de estas enzimas en raíz, a las 24 hora posterior a su inoculación.
Palabras clave: enzimas pectinolíticas, Mycorrhizae, respuesta de defensa.
ABSTRACT
The arbuscular mycorrhizal fungi and elicitor have been many used with diverse objectives in modern agriculture. Systemin is a polypeptide hormone with many possibilities like elicitor of answers of defense in Solanaceas. thus its application with the AMF, would allow increasing the effects in the protection against diverse pathogens. An experiment was made in which the possible participation of the systemin in the establishment of the mycorrhiza and the induction of defense response in pre-symbiotic stages was evaluated. The exogenous application of systemin in the roots produce the local and systemic induction of β-1,3-glucanase and chitinase in early stages. A synergic effect between systemin and AMF (F. mosseae) on the activity of β-1,3-glucanases in root from the first later hour the application was observed. F. mosseae produce the induction to these enzymes in root, at 24 hour the inoculation.
Key Words: pectinolytic enzymes, Mycorrhizae, defense mechanisms.
]]>
INTRODUCCIÓN
Los hongos micorrícicos arbusculares (HMA) pertenecen al phylum Glomeromycota, los que constituyen uno de los microorganismo del suelo de mayor importancia (1). Durante el proceso de establecimiento y desarrollo de la simbiosis entre los HMA y las plantas se produce la inducción de respuesta de resistencia con la inducción de proteínas diversas relacionadas con la patogenicidad (PRs), entre las que se reportan las β-1,3-glucanasas (GLN), quitinasas (QUI), y enzimas relacionadas con los procesos oxidativos (2,3). La inducción de estos mecanismos en etapas tempranas del establecimiento de la asociación micorrícica ha recibido especial atención debido a su posible implicación en la regulación de la simbiosis, así como en la protección de las plantas contra el ataque de patógenos (4).
La sistemina (S) fue la primera hormona peptídica identificada en plantas, la cual induce respuestas de defensa en Solanáceas ante la afectación por insectos y daño mecánico (5). Recientemente, se ha vinculado con la disminución de la incidencia de patógenos necrotróficos de origen fúngico (6), con la expresión significativa de genes que codifican para componentes claves del sistema de señalización de la defensa sistémica inducida (7). Posterior al reconocimiento de la sistemina en superficie celular por el receptor, se activa un cascada de señales que involucra la acumulación transciente de ácido jasmónico (AJ) y como consecuencia, la expresión de genes relacionados con las respuestas de defensa (6,8). Entre las proteínas que se inducen se encuentran las llamadas Proteínas de Respuesta Sistémicas al Daño (9), las cuales incluyen varias proteínas asociadas a las vías de señalización y otras similares a las inducidas por insectos y herbívoros (10). Estudios previos han demostrado la inducción por la sistemina de Proteínas Relacionadas con la Patogénesis (PRs) (11,12).
Estudios recientes dirigidos a analizar el posible papel de la sistemina en la modulación local y/o sistémica de la asociación micorrícica indican que la aplicación en plantas de tomate micorrizadas induce la acumulación de β-1,3-glucanasas y quitinasas (12).
Con el objetivo de poder dilucidar la posible interconexión de las vías de señalización inducida por la sistemina y los HMA, se evaluó el patrón de la actividad de proteínas PRs inducidas por esta interacción, en la fase presimbiótica.
MATERIALES Y MÉTODOS
Obtención del material vegetal y aplicación de los productos
Se emplearon plantas de tomate (Solanum lycopersicum L.) del cultivar ´Amalia´ (13). Las semillas fueron desinfectadas superficialmente con hipoclorito de sodio comercial al 10 % durante 10 min, seguido de tres lavados con agua destilada (H2O d) estéril, posteriores a los cuales se procedió a su siembra. El sustrato estuvo conformado por una mezcla de suelo ferralítico rojo lixiviado típico y humus de lombriz en relación 1:1 (v:v), estéril (Tabla I). Las plantas se desarrollaron en cámara de crecimiento, en condiciones controladas, con fotoperíodo de 8 h de oscuridad a 16 °C y 16 h de luz a 28 °C.
Se seleccionó la especie de HMA Funneliformis mosseae (T.H. Nicolson & Gerd.) (14) (Fm), procedente del cepario del Laboratorio de Micorrizas del Instituto Nacional de Ciencias Agrícolas (INCA).
Se emplearon bandejas de 120 pocillos de 10 mL de capacidad cada uno, donde se sembró una semilla desinfectada en cada uno. A los 21 días posteriores a la emergencia, se procedió a la inoculación del hongo micorrizógeno y a la aplicación de la sistemina en el sustrato, de forma independiente y combinada, según el diseño de los tratamientos. El HMA se preparó en una suspensión líquida con una concentración de 20 esporas mL-1, de la cual se añadió 1 mL por planta en el sustrato, en los tratamientos micorrizados y la sistemina sintética de tomate (Laboratorios BQ SOS, de México) fue adicionada al sustrato (1 mL por planta a 44,06 nmol) (12), en los tratamientos correspondientes. En el tratamiento control se adicionó agua destilada (2 mL por planta). Para los muestreos se siguió una dinámica durante las 48 horas posterior a la inoculación (hpi): 1, 2, 4, 8, 12, 18, 24 y 48 hpi.
En los momentos de muestreos las plantas fueron lavadas y separadas por órganos, dividiéndose las raíces en dos grupos. Una porción se empleó para la determinación de las actividades enzimáticas, la cual fue congelada en nitrógeno líquido y conservada a -70 C, mientras que la otra se secó en estufa a 70 C para la determinación del establecimiento de la simbiosis micorrícica (colonización).
Colonización micorrícica
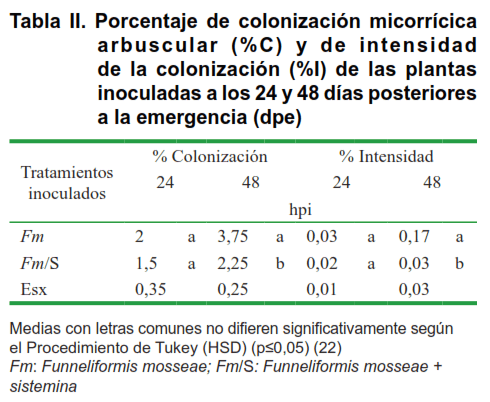
Para cuantificar la colonización micorrícica, las raíces secas a 70C se digirieron con KOH, para luego teñirse con azul de tripano. Se evaluó el porcentaje de colonización micorrícica (%C) y la intensidad de la colonización según el método de los interceptos (%I) (12).
Ensayo enzimático
La extracción de proteína y las determinaciones enzimáticas se realizaron según la metodología descrita por Noval y colaboradores (12). A los extractos se les determinó la concentración de proteína mediante el método de Bradford (16), empleando el kit comercial (Bio-Rad Laboratories, USA). Se realizó la determinación de las actividades enzimáticas in vitro en las raíces y en las hojas de las plantas de tomate. Para la cuantificación de la actividad β-1,3-glucanasa (GLN; EC- 3.2.1.39) se utilizó laminarina como sustrato y se leyó la absorbancia a 450 nm en un lector de microplacas SUMA, PR 521. Para la actividad quitinasa (QUI; EC- 3.2.1.14) se usó como sustrato la quitina coloidal preparada a partir de quitina grado reactivo (Fluka) y la absorbancia se leyó a 585 nm en el espectrofotómetro (12). Todas las actividades enzimáticas fueron transformadas a microkatales (µKat) y nanokatales (Kat) por miligramos de proteínas, según fue necesario (17). Las lecturas de la absorbancia en los diferentes métodos empleados se realizaron en un espectrofotómetro, Ultrospec Plus Spectrophotometer, Pharmacia LKB. Todos los sustratos fueron adquiridos de Sigma-Aldrich Chemical (St. Louis, MO, USA).
Diseño experimental y análisis estadístico
El experimento se desarrolló siguiendo un diseño completamente aleatorizado con tres repeticiones, conformadas cada una por 10 plantas, en cada momento de muestreo. Los datos fueron procesados estadísticamente según el análisis de varianza de clasificación simple, comparándose las medias mediante los intervalos de confianza con un grado de significación de 95 %. El experimento se repitió tres veces en el tiempo, de los cuales se seleccionó uno representativo. Todas las comparaciones se realizaron según el paquete estadístico STATGRAPHIC Plus Versión 5.1 (18). Los porcentajes de incrementos se calcularon comparando los tratamientos con el control correspondiente.
RESULTADOS Y DISCUSIÓN
]]> Dinámica del establecimiento de la simbiosis En la variante donde se realizó la aplicación combinada del HMA y del elicitor se alcanzaron niveles del establecimiento de la micorrización inferiores a los observados en las plantas donde solo se inoculó F. mosseae. Esta respuesta puede deberse a que en las raíces de las plantas donde se aplicaron ambos productos se produce la ocurrencia de dos eventos, la inducción de respuestas rápidas por la sistemina (9,11) y el estrés producido por los procesos de penetración y diseminación del micelio del hongo en las raíces (19–21), procesos que han sido documentados en la literatura, pero de forma independiente.
En ambos indicadores se observaron niveles bajos (1,5-3,75 %C y 0,023-0,16 %I), lo cual indica que se encontraba en las primeras fases del establecimiento del micelio de F. mosseae en las raíces. A pesar de ello, la evaluación de la colonización en etapas tempranas permitió detectar los momentos iniciales del establecimiento de la simbiosis, los que ocurrieron entre las 24 y 48 horas.
Patrón de actividad de enzimas relacionadas con defensa: glucanasas y quitinasas
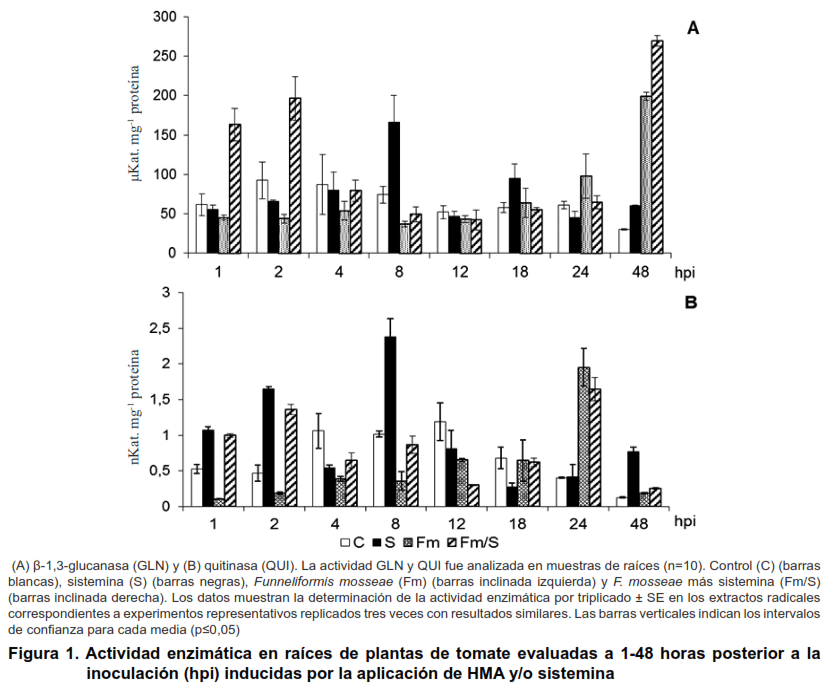
Los patrones de actividad de las enzimas GLN en las raíces de las plantas de tomate inoculadas con F. mosseae de forma independiente (Fm) mostraron niveles de actividad similares o inferiores a las obtenidas en el tratamiento control en las primeras 18 hpi (nivel basal), con diferencias significativas entre ellos (Figura 1A). A partir de las 24 horas se produjo un incremento gradual que alcanzó un pico significativo a las 48 hpi (98,05 y 199,08 µKat mg-1 proteína), con incrementos de 60,44 % y 558,85 %, respectivamente.
]]>
Fm) se produjo la disminución de la actividad de esta enzima hasta las 12 horas, con relación a los niveles alcanzados por el control, con los cuales mostraron diferencias significativas (Figura 1B). A las 24 horas se produjo un incremento transciente (1,95 nKat mg-1 proteína), que difirió estadísticamente con las demás variantes estudiadas.
Se ha encontrado que en las plantas micorrizadas se produce la regulación de la expresión de las quitinasas, y se ha propuesto un modelo especulativo de los mecanismos de supresión de la expresión constitutiva de esta enzima (tratamiento control), inducidos por las MA en estadios tempranos (23). En el mismo se observa que en estadios tardíos se produce la acumulación de quitinasas específicas inducidas exclusivamente por la micorrización. Este mecanismo podría explicar el proceso de regulación de la diseminación del HMA en la raíz.
La aplicación exógena de la sistemina, de forma independiente indujo la actividad GLN a las ocho, 18 y 48 horas, siendo significativo los niveles alcanzados a los ocho horas (166,33 µKat mg-1 proteína), correspondientes a 123,85 % de incrementos. Con relación a la actividad QUI se produjo una inducción temprana desde los primeros momentos de evaluación, 1 y 2 hpi (1,07 y 1,64 38 nKat mg-1 proteína), correspondientes a 103,97 y 251,75 % de incrementos, respectivamente. Se produjo un segundo incremento a las ocho horas (2,38 nKat mg-1 proteína), que mostró el mayor nivel alcanzado en el experimento, con 133,96 % de incrementos y diferencias significativas con todas las variantes estudiadas. La inducción de las GLN y QUI producida por la sistemina per se, en las primeras horas posteriores a su aplicación en las raíces, sugiere, que en las raíces de las plantas de tomate deben existir receptores de la sistemina, donde participe la proteína SR-160 u otra similar, que actúe como señal primaria, de forma similar a como ha sido demostrado en hoja (24).
Se observó un efecto sinérgico con la aplicación conjunta de Fm y de la sistemina, con niveles importantes de la actividad de las GLN a las 1 y 2 hpi (163,16 y 196,53 µKat mg-1 proteína con 223,31 y 258,37 % de incremento con relación a los tratamientos donde se aplicaron de forma independiente) y a las 48 hpi con 269,22 µKat mg-1 proteína y un incremento significativo de 790,99 % con relación al control. Con relación a la actividad QUI se observó un efecto inhibitorio en las plantas inoculadas con Fm, el cual fue superado durante las etapas tempranas (1-8 hpi) al ser combinado con la aplicación de la sistemina (Fm/S). No se observó un efecto sinérgico entre Fm y la sistemina sobre la actividad de esta enzima.
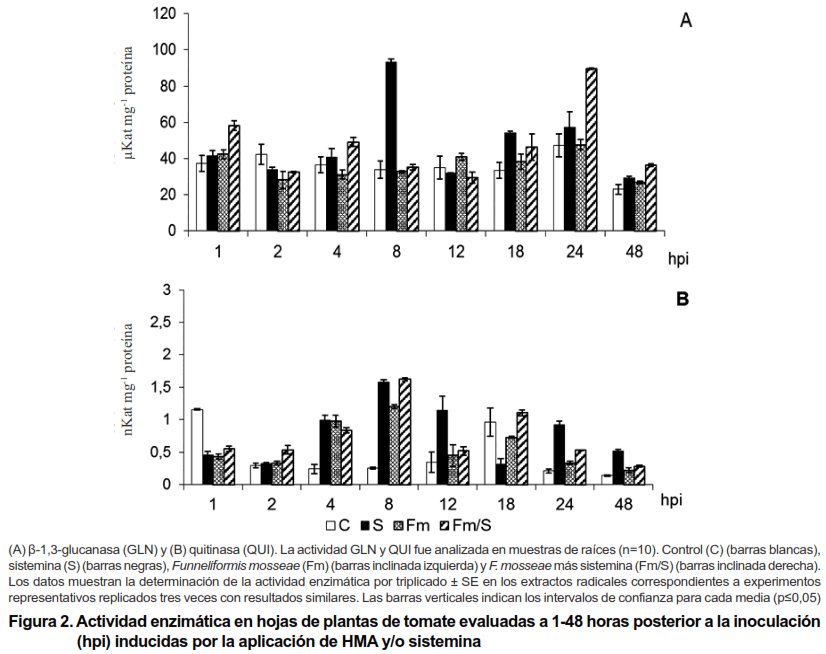
Al analizar la cuantificación de la actividad de las GLN y QUI en hojas se observó que se alcanzaron niveles más bajos que los obtenidos en las raíces (Figura 2). En este órgano también se encontró la presencia de un nivel basal de las GLN dado por el tratamiento control, con valores promedio de 38,26 µKat mg-1 proteína (Figura 2A), el cual no fue modificado por la inoculación del HMA cuando se aplicó de forma independiente (Fm). De igual forma, se encontró la presencia de niveles basales de la enzima QUI (control), los que fueron superados por los tratamientos, en la mayoría de los tiempos en que se realizó la evaluación (Figura 2B). La inoculación con Fm, de forma independiente indujo incrementos de la actividad de esta enzima a las cuatro y ocho hpi (0,98 y 1,2 nKat mg-1 proteína) con incrementos de 304,47 y 374,79 %, respectivamente; sin embargo, en el resto de los momentos evaluados, esta variante no superó al control.
]]>
-1 proteína), los cuales mostraron incrementos de 175,87 y 62,30 %, respectivamente. Con relación a las QUI la inducción se observó a las cuatro hpi, la que fue mantenida en el tiempo (8 y 12 hpi), con valores de 0,99, 1,58 y 1,14 nKat mg-1 proteína e incrementos de 310,09; 522,77 y 234,71 %, respectivamente. Posteriormente, durante las 24 y 48 hpi se produjo una segunda inducción de menor magnitud a la observada con anterioridad (0,91 y 0,51 nKat mg-1 proteína), momento en los cuales se alcanzaron los niveles basales más bajos.
De forma similar a lo obtenido en la raíz en hoja se observó también un efecto sinérgico entre el HMA y el elicitor (Fm/S) sobre la actividad GLN en estadios tempranos (1 hpi) que alcanzó niveles de 58,22 µKat mg-1 proteína, el cual fue más significativo a las 24 hpi, con 89,63 µKat mg-1 proteína y 89,46 % de incremento; sin embargo, con relación a las QUI no se observó un efecto sinérgico en ninguno de los momentos evaluados.
En las plantas micorrizadas se produjo la inducción de las GLU solo a partir de las 24 hpi con un máximo de actividad a las 48 hpi, mientras que las QUI solo mostraron niveles superiores al control a las 24 hpi. En esta fase ocurre el reconocimiento entre los simbiontes, con la activación de una serie de cascadas de señales, resultantes en la producción de hormonas relacionadas con defensa (25–27). Se ha planteado que los estadíos iniciales de la colonización radical por los HMA son acompañados por la inducción transciente de las defensas de la planta seguida por la supresión en estadíos tardíos de la interacción (4,28,29).
A pesar de que las 24 y 48 hpi fueron los momentos de evaluación finales del experimento, estos corresponden a fases tempranas del proceso de establecimiento de la simbiosis MA como lo demostraron los niveles de colonización e intensidad alcanzados en los mismos (Tabla II).
Estudios previos han demostrado que la aplicación de forma exógena de la sistemina en las raíces induce respuestas de defensa a nivel local y sistémico (12) y la acumulación de Proteínas de Respuesta Sistémicas al Daño (SWRPs) (9). En el presente trabajo se corrobora el carácter de la sistemina de inducción de respuestas rápidas, el que fue más marcado sobre la actividad QUI con incrementos desde las primeras horas de evaluación (1 y 2 hpi), mostrando a las 8 hpi el nivel más elevado de todo los tratamientos en estudio. Para las GLU la respuesta fue más tardía mostrando incrementos solo a las 8 hpi, con un segundo momento de inducción a las 18 y 48 hpi pero de intensidad menor.
]]>
El sinergismo observado en fases muy tempranas con la aplicación de Fm y de la sistemina sobre la actividad GLU, manifiesta la posible acción conjunta de componentes de las vía de traducción de señales del elicitor y de las respuestas activadas durante la simbiosis micorrícica en la fase presimbiótica, correspondiente a procesos de señalización y comienzo de la formación de los apresorios.
Se conoce que luego de la recepción de la sistemina se activa una serie de reacciones donde se produce la entrada de calcio, la activación de la actividad de las MAPK, la producción de especies activas de oxígeno, el incremento de la actividad de la enzima ATPasaH+, con la consecuente hiperpolarización de la membrana, la acumulación de hormonas como el ácido jasmónico y la acumulación de señales secundarias como las oxilipinas (32).
Diferentes autores han informado sobre elementos que constituyen señales secundarias o inductores de respuestas de defensa durante el proceso de micorrización. De esta forma se ha observado que en plantas micorrizadas se produce de forma temprana la expresión de quinasas, unas de ellas dependiente de cadmodulina y Ca (CCaMKs) (26,33), y de elementos del proceso de señalización como los receptores de la simbiosis (SYMRK), los cuales son requeridos como señal primaria del proceso de micorrización (34).
Varios de estos componentes son comunes a las vías de transducción de señales inducidas tanto por los HMA, como por la sistemina, los que constituyen elementos importantes de la ruta de señalización que conducen a la acumulación del ácido jasmónico (6,26,27,35) y de esta forma a la inducción de respuestas de defensa, entre las que se encuentran las proteínas PRs.
]]>
Fm y la sistemina no es producto de un efecto sinérgico. En los momentos evaluados entre 1 y 8 hpi se observó el mismo patrón de comportamiento al encontrado con la aplicación del elicitor de forma independiente, mientras que la respuesta observada a las 24 hpi, al parecer, es debido al efecto del HMA ya muestra niveles similares al obtenido por este, cuando se aplicó de independiente (Fm), momento en el cual la sistemina alcanzó niveles bajos.
CONCLUSIÓN
Los resultados sugieren un importante papel de la sistemina en su interacción con la simbiosis micorrícica arbuscular, al modular de forma temprana y transciente las respuestas de defensa durante el proceso de establecimiento de la micorrización, potenciando la acción de los HMA, el cual se refleja en la inducción de las proteínas PR. La sistemina per se produce la inducción rápida de las β-1,3-glucanasas y quitinasas en las primeras horas posteriores a su aplicación en las raíces. Se obtuvo un efecto sinérgico de esta hormona polipeptídica con el HMA (F. mosseae) sobre la inducción de β-1,3-glucanasa y quitinasa, desde etapas tempranas en raíz (1 hpi), mientras que en hoja fue más tardío. F. mosseae solo produjo la inducción de estas enzimas en raíz, en estadíos tardíos.
BIBLIOGRAFÍA
1. Berruti A, Borriello R, Lumini E, Scariot V, Bianciotto V, Balestrini R. Application of laser microdissection to identify the mycorrhizal fungi that establish arbuscules inside root cells. Frontiers in Plant Science. 2013;4. doi:10.3389/fpls.2013.00135
2. Balasubramanian V, Vashisht D, Cletus J, Sakthivel N. Plant β-1,3-glucanases: their biological functions and transgenic expression against phytopathogenic fungi. Biotechnology Letters. 2012;34(11):1983–90. doi:10.1007/s10529-012-1012-6
3. Evelin H, Kapoor R. Arbuscular mycorrhizal symbiosis modulates antioxidant response in salt-stressed Trigonella foenum-graecum plants. Mycorrhiza. 2014;24(3):197–208. doi:10.1007/s00572-013-0529-4
4. Jung SC, Martinez-Medina A, Lopez-Raez JA, Pozo MJ. Mycorrhiza-Induced Resistance and Priming of Plant Defenses. Journal of Chemical Ecology. 2012;38(6):651–64. doi:10.1007/s10886-012-0134-6
5. Pearce G, Strydom D, Johnson S, Ryan CA. A Polypeptide from Tomato Leaves Induces Wound-Inducible Proteinase Inhibitor Proteins. Science. 1991;253(5022):895–7. doi:10.1126/science.253.5022.895
6. El Oirdi M, El Rahman TA, Rigano L, El Hadrami A, Rodriguez MC, Daayf F, et al. Botrytis cinerea Manipulates the Antagonistic Effects between Immune Pathways to Promote Disease Development in Tomato. The Plant Cell. 2011;23(6):2405–21. doi:10.1105/tpc.111.083394
7. Coppola M, Corrado G, Coppola V, Cascone P, Martinelli R, Digilio MC, et al. Prosystemin Overexpression in Tomato Enhances Resistance to Different Biotic Stresses by Activating Genes of Multiple Signaling Pathways. Plant Molecular Biology Reporter. 2015;33(5):1270–85. doi:10.1007/s11105-014-0834-x
8. Wasternack C, Hause B. Jasmonates: biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Annals of Botany. 2013;111(6):1021–58. doi:10.1093/aob/mct067
9. Savatin DV, Gramegna G, Modesti V, Cervone F. Wounding in the plant tissue: the defense of a dangerous passage. Frontiers in Plant Science. 2014;5. doi:10.3389/fpls.2014.00470
10. Rehrig EM, Appel HM, Jones AD, Schultz JC. Roles for jasmonate- and ethylene-induced transcription factors in the ability of Arabidopsis to respond differentially to damage caused by two insect herbivores. Frontiers in Plant Science. 2014;5:407–21. doi:10.3389/fpls.2014.00407
11. Chen Y-L, Lee C-Y, Cheng K-T, Chang W-H, Huang R-N, Nam HG, et al. Quantitative Peptidomics Study Reveals That a Wound-Induced Peptide from PR-1 Regulates Immune Signaling in Tomato. The Plant Cell. 2014;26(10):4135–48. doi:10.1105/tpc.114.131185
12. de la Noval B, Pérez E, Martínez B, León O, Martínez-Gallardo N, Délano-Frier J. Exogenous systemin has a contrasting effect on disease resistance in mycorrhizal tomato (Solanum lycopersicum) plants infected with necrotrophic or hemibiotrophic pathogens. Mycorrhiza. 2007;17(5):449–60. doi:10.1007/s00572-007-0122-9
13. Alvarez M, Armas G de, Martínez B. Amalia y Mariela, dos nuevas variedades de tomate para consumo fresco. Cultivos Tropicales. 1997;18(1):83.
14. Paneque PVM, Calaña NJM, Calderón VM, Borges BY, Hernández GTC, Caruncho CM. Manual de técnicas analíticas para análisis de suelo, foliar, abonos orgánicos y fertilizantes químicos [Internet]. 1st ed. La Habana, Cuba: Ediciones INCA; 2010 [cited 2016 Jan 27]. 157 p. Available from: http://mst.ama.cu/578/
15. Schüßler A, Walker C. The Glomeromycota: A species list with new families and new genera [Internet]. U.K.: CreateSpace Independent Publishing Platform; 2010 [cited 2017 Jul 7]. 58 p. Available from: https://www.amazon.com/Glomeromycota-species-list-families-genera/dp/1466388048
16. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 1976;72(1–2):248–54. doi:10.1016/0003-2697(76)90527-3
17. Tipton K. Principles of enzyme assays and kinetic studies. In: Eisenthal R, Danson MJ, editors. Enzyme Assays: A Practical Approach [Internet]. U.K.: Oxford University Press; 2002 [cited 2017 Jul 7]. p. 1–58. Available from: https://books.google.com.cu/books?id=W-KAirjgqdQC
18. Statistical Graphics Crop. STATGRAPHICS® Plus [Internet]. Version 5.1. 2000. (Profesional). Available from: http://www.statgraphics.com/statgraphics/statgraphics.nsf/pd/pdpricing
19. Rich MK, Schorderet M, Reinhardt D. The role of the cell wall compartment in mutualistic symbioses of plants. Frontiers in Plant Science. 2014;5. doi:10.3389/fpls.2014.00238
20. Lanfranco L, Young JPW. Genetic and genomic glimpses of the elusive arbuscular mycorrhizal fungi. Current Opinion in Plant Biology. 2012;15(4):454–61. doi:10.1016/j.pbi.2012.04.003
21. Campos-Soriano L, García-Martínez J, Segundo BS. The arbuscular mycorrhizal symbiosis promotes the systemic induction of regulatory defence-related genes in rice leaves and confers resistance to pathogen infection: Resistance to M. oryzae in mycorrhizal rice. Molecular Plant Pathology. 2012;13(6):579–92. doi:10.1111/j.1364-3703.2011.00773.x
22. Tukey JW. Bias and confidence in not quite large samples. The Annals of Mathematical Statistics. 1958;29(2):614–23. doi:10.1214/aoms/1177706647
23. Salzer P, Boller T. Elicitor induced reactions in mycorrhizae and their suppression. In: Podila GK, Douds DD, editors. Current advances in mycorrhizae research. St. Paul, USA: APS Press; 2000. p. 1–10.
24. Scheer JM, Ryan CA. A 160-kD Systemin Receptor on the Surface of Lycopersicon peruvianum Suspension-Cultured Cells. The Plant Cell. 1999;11(8):1525–36. doi:10.1105/tpc.11.8.1525
25. Liu Y, Gianinazzi-Pearson V, Arnould C, Wipf D, Zhao B, van Tuinen D. Fungal genes related to calcium homeostasis and signalling are upregulated in symbiotic arbuscular mycorrhiza interactions. Fungal Biology. 2013;117(1):22–31. doi:10.1016/j.funbio.2012.11.002
26. Pozo MJ, López-Ráez JA, Azcón-Aguilar C, García-Garrido JM. Phytohormones as integrators of environmental signals in the regulation of mycorrhizal symbioses. New Phytologist. 2015;205(4):1431–6. doi:10.1111/nph.13252
27. Fernández I, Merlos M, López-Ráez JA, Martínez-Medina A, Ferrol N, Azcón C, et al. Defense Related Phytohormones Regulation in Arbuscular Mycorrhizal Symbioses Depends on the Partner Genotypes. Journal of Chemical Ecology. 2014;40(7):791–803. doi:10.1007/s10886-014-0473-6
28. Cameron DD, Neal AL, van Wees SCM, Ton J. Mycorrhiza-induced resistance: more than the sum of its parts? Trends in Plant Science. 2013;18(10):539–45. doi:10.1016/j.tplants.2013.06.004
29. Pérez E, Rodríguez Y, Fernández K, de La Noval BM, Hernández A. Percepción de señales de los hongos micorrízicos arbusculares por plantas de tomate (Solanum lycopersicum L.) en las fases iniciales del establecimiento de la simbiosis. Cultivos Tropicales. 2015;36(3):40–4.
30. Yan L, Zhai Q, Wei J, Li S, Wang B, Huang T, et al. Role of Tomato Lipoxygenase D in Wound-Induced Jasmonate Biosynthesis and Plant Immunity to Insect Herbivores. PLOS Genetics. 2013;9(12):e1003964. doi:10.1371/journal.pgen.1003964
31. Lee GI, Howe GA. The tomato mutant spr1 is defective in systemin perception and the production of a systemic wound signal for defense gene expression. The Plant Journal. 2003;33(3):567–76. doi:10.1046/j.1365-313X.2003.01646.x
32. Huffaker A, Pearce G, Veyrat N, Erb M, Turlings TCJ, Sartor R, et al. Plant elicitor peptides are conserved signals regulating direct and indirect antiherbivore defense. Proceedings of the National Academy of Sciences. 2013;110(14):5707–12. doi:10.1073/pnas.1214668110
33. Shimoda Y, Han L, Yamazaki T, Suzuki R, Hayashi M, Imaizumi-Anraku H. Rhizobial and Fungal Symbioses Show Different Requirements for Calmodulin Binding to Calcium Calmodulin-Dependent Protein Kinase in Lotus japonicus. The Plant Cell. 2012;24(1):304–21. doi:10.1105/tpc.111.092197
34. Nair A, Bhargava S. Reduced mycorrhizal colonization (rmc) tomato mutant lacks expression of SymRK signaling pathway genes. Plant Signaling & Behavior. 2012;7(12):1578–83. doi:10.4161/psb.20156
35. León-Morcillo RJ, Martín-Rodríguez JÁ, Vierheilig H, Ocampo JA, García-Garrido JM. Late activation of the 9-oxylipin pathway during arbuscular mycorrhiza formation in tomato and its regulation by jasmonate signalling. Journal of Experimental Botany. 2012;63(10):3545–58. doi:10.1093/jxb/ers010
Recibido: 28/09/2016
Aceptado: 28/03/2017
Dra.C. Blanca M. de la Noval-Pons, Instituto Nacional de Ciencias Agrícolas. Gaveta Postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700.Email: bdelanov@inca.edu.cu
]]>
{kind=link}
{kind=link}