Efecto de diferentes medios de cultivo sobre el desarrollo de Phytophthora nicotianae Breda de Haan
Effect of different culture media on Phytophthora nicotianae Breda de Haan develop
Instituto Nacional de Ciencias Agrícolas (INCA). Carretera a Tapaste km 3 ½, San José de las Lajas, Mayabeque, Cuba. Correo electrónico: dianevys@inca.edu.cu.
El objetivo del estudio fue determinar el efecto de diferentes medios de cultivo sobre el crecimiento, esporulación y secreción de enzimas hidrolíticas de Phytophthora nicotianae Breda de Haan. Para ello se emplearon los medios en estado sólido y líquido: Papa Dextrosa Agar (PDA), jugo de verduras (V8) y extracto de tabaco, combinados entre ellos y con suplemento de asparagina y carbonato de calcio. El mayor crecimiento del microorganismo se produjo en medio sólido con extracto de tabaco, pues alcanzó en menor tiempo el extremo de la placa; sin embargo, su crecimiento fue poco ramificado, a diferencia de los que contenían V8. Se comprobó que la biomasa producida fue mayor en los medios líquidos que contenían V8. El número de zoosporas estimadas en el momento en que el micelio alcanzó el borde de la placa fue significativamente mayor en el tratamiento que combinaba PDA con V8 y el menor valor se obtuvo al emplearse el extracto de tabaco. Al utilizarse la combinación del extracto de tabaco con V8, se obtuvieron las mayores actividades poligalacturonasa y xilanasa. Los resultados evidenciaron que los medios que favorecen la expresión de enzimas hidrolíticas, así como el crecimiento y esporulación de P. nicotianae, tienen V8 entre sus componentes, elemento de gran importancia, pues se disminuye el tiempo requerido para los ensayos y con ello el riesgo de contaminación de los experimentos.
Palabras clave: Phytophthora nicotianae, medio de cultivo, crecimiento, esporulación, actividad enzimática.
ABSTRACT
The effect of different culture media on Phytophthora nicotianae growth, sporulation, and hydrolytic enzymes secretion was studied. The liquid and solid culture media used were potato dextrose agar (PDA), vegetable juice (V8), and tobacco extract. They were combined one another and supplemented with asparagine and calcium carbonate. Phytophthora nicotianae growing on solid media containing only tobacco extract reached the plate edge in the shortest time; however, the mycelium was little branched, unlike the mycelium in the media containing V8. A greater biomass production was observed in liquid culture with V8. The zoospore number was significantly higher in the treatment combining PDA only with V8; the lowest number was obtained with tobacco extract. Polygalacturonase and xylanase activities were the highest when the liquid media contained the combination of tobacco extract and V8 juice. The culture medium V8 was shown to be useful to determine the enzymatic activity of the hydrolytic enzymes and the parameters of growth and sporulation of P. nicotianae; itreduced the time required for the trials and, consequently, the risk of contamination.
Key words: Phytophthora nicotianae, culture media, growth, sporulation, enzymatic activity.
INTRODUCCIÓN
Phytophthora nicotianae (Breda de Haan), es un fitopatógeno con una amplia gama de hospedantes que incluye un gran número de especies de Solanáceas (1). En Cuba, es elevada la incidencia de esta especie sobre cultivos de importancia económica como el tabaco, donde provoca la enfermedad conocida como Pata prieta (2). ]]>
Para establecer la infección en el vegetal, el microorganismo necesita secretar enzimas hidrolíticas que le permitan la penetración. Estas son conocidas como CWDEs (Cell Wall Degrading Enzymes, por sus siglas en inglés), puesto que su función es degradar las paredes celulares que constituyen la primera barrera de resistencia de la planta al patógeno (3). Se cree que estas enzimas y otras proteínas extracelulares son transportadas y secretadas en pequeñas vesículas ubicadas hacia la zona apical de la hifa de los oomicetos (4). Esas vesículas también están presentes en los quistes de zoosporas de Phytophthora y contienen además, compuestos adhesivos que le permiten fijarse a sus hospedantes, así como otras moléculas involucradas en la defensa, tales como inhibidores de glucanasas y proteasas (4,5,6,7). Estas y otras proteínas, son secretadas hacia el interior de la célula vegetal a través del haustorio (8).Entre las CWDEs también se incluyen celulasas, poligalacturonasas (PG) y xilanasas. La pectina es uno de los principales componentes de las paredes celulares de numerosas plantas y es escindida por pectinasas como poligalacturonasas, pectin metil esterasas (PME) y pectato liasas (PL) (3). Las PG, en general, hidrolizan los enlaces a1-4 entre los ácidos galacturónicos de manera aleatoria, produciendo mono, di y oligalacturonatos (9). El xilano también forma parte de las paredes celulares vegetales y son las xilanasas, las enzimas encargadas de su degradación. En ensayos in vitro se comprobó que pueden ser secretadas diferentes tipos de enzimas, en dependencia de las condiciones de crecimiento (9). Estas investigaciones tomaron auge en las últimas décadas y algunos trabajos se dirigieron a optimizar procesos fermentativos para la obtención de estas enzimas, en particular de xilanasas, debido a su uso en la industria (10, 11, 12, 13, 14, 15, 16).
Hasta el momento, se han caracterizado algunos genes que codifican para estas enzimas entre los miembros del grupo oomycetes (17). Estos incluyen b- glucosidasa/ b xilosidasa y endopoligalacturonasas de diferentes especies de Phytophthora (17, 18).
Los estudios relacionados con medios de cultivo que favorecen la expresión de estas enzimas en Phytophthora son escasos (9, 19, 20); a pesar de que los valores de las actividades enzimáticas son considerados una medida de la patogenicidad de la cepa (9). Se puede considerar entonces que, un medio que favorezca la secreción de estas enzimas por parte del microorganismo, resultaría útil para ensayos que involucren la inoculación y el desarrollo de la infección en una planta hospedante.
Los experimentos in vitro con diferentes medios de cultivo, permiten conocer el comportamiento relacionado con el crecimiento y la esporulación de los microorganismos. A menudo se emplea el medio de cultivo Papa Dextrosa Agar (PDA, por sus siglas en inglés) para el cultivo de P. nicotianae, aunque se comprobó que la velocidad de crecimiento y la esporulación es superior en otros medios con extractos naturales, como los que poseen avena, maíz, zanahoria y tomate (1, 21, 22). Esto se debe, fundamentalmente, a la presencia de esteroles, los que constituyen un requerimiento importante para especies del género Phytophthora (1). La optimización de medios de cultivo y la búsqueda de alternativas más eficientes, resulta imprescindible para disminuir el tiempo de duración de los ensayos y los riesgos de contaminación de los experimentos. También es preciso considerar que, a medida que se realizan sucesivos cultivos de una cepa esta pierde virulencia, por lo que resulta imperioso contar con un medio natural, donde este rasgo se afecte lo menos posible.
El objetivo de este trabajo fue evaluar el efecto de diferentes medios de cultivo sobre el crecimiento, esporulación y secreción de enzimas hidrolíticas de P. nicotianae.
MATERIALES Y MÉTODOS
En los experimentos se empleó la cepa SS-11 de P. nicotianae, raza 0, grupo 1, aislada del cultivar de tabaco Habana 92, procedente del Instituto de Investigaciones del Tabaco.
Se partió de cultivos desarrollados previamente en medio PDA durante 7 días en condiciones de oscuridad y conservados a 27ºC de temperatura. ]]>
Crecimiento y esporulación de P. nicotianae en diferentes medios de cultivo sólidosPara determinar el efecto en el crecimiento y esporulación de P. nicotianae se emplearon tres medios de cultivo combinados y con suplemento de asparagina (Asp) y carbonato de calcio (CaCO3). Para los ensayos en medio sólidos se utilizaron placas Petri de 90mm de diámetro que contenían 25 ml de los medios:
a) V8 + Glucosa + Asp + CaCO3
b) Ext Tab + Glucosa + Asp + CaCO3
c) V8 + Ext Tab + Glucosa + Asp + CaCO3
d) PDA + Asp + CaCO3
e) PDA + V8 + Asp + CaCO3
f) PDA + Ext Tab + Asp + CaCO3
g) PDA + V8 + Ext Tab + Asp + CaCO3
h) PDA ]]>
Se empleó el jugo de verduras (V8), de la línea comercial Del Frutal (Guatemala), el cual contiene: tomate, apio, espinacas, remolacha, lechuga, perejil, berro y zanahoria. El jugo de verduras (V8) y el extracto de tabaco se emplearon al 20%. Este último se preparó con 100g de foliolo.L-1 de agua destilada, que se mezcló en licuadora. Se adicionó carbonato de calcio (2g.L-1), asparagina (2g.L-1), glucosa (20g.L-1) y se incluyó agar (20g.L-1) en los casos que no contenían PDA.Se establecieron 8 placas por tratamiento y en el centro se colocó un disco de 10mm de diámetro con micelio de P. nicotianae de 5-7 días de cultivado. Las placas se incubaron a 27°C en condiciones de oscuridad. El crecimiento radial se midió con el empleo de una regla graduada, a partir de las 72 horas y se determinó cada 24 horas hasta los 7 días.
Se realizó el conteo de zoosporas en 4 de las 8 réplicas, cuando el micelio alcanzó el borde de la placa en cada uno de los tratamientos. Para ello, se adicionaron 10 ml de agua destilada por placa y suavemente se maceró la colonia con espátula de Drygalski sin destruir el agar. Las placas se incubaron a 4°C durante 2 horas seguido de un choque térmico durante 30 minutos a 37°C, para la liberación de las zoosporas. El conteo de la suspensión de esporas se realizó en cámara de Neubauer. A los 14 días se realizó el conteo a las 4 restantes placas para determinar la presencia y cuantía de zoosporas transcurrido ese tiempo.
Crecimiento y actividades enzimáticas en medios de cultivo líquidos
Para el crecimiento del microorganismo en medio líquido se emplearon los medios de cultivo:
a) V8 + Glucosa + Asp + CaCO3
b) Ext Tab + Glucosa + Asp + CaCO3
c) V8 + Ext Tab + Glucosa + Asp + CaCO3
d) PDA
e) PDA + Asp + CaCO3 ]]>
En los casos en que se empleó el medio PDA, este se filtró para eliminar el agar. El pH se ajustó a 5,6 con HCl y NaOH, en dependencia de los casos, y se midió nuevamente el pH después de esterilizar los medios de cultivo en autoclave durante 20 minutos.Se utilizaron erlenmeyers de 250ml de capacidad que contenían 70ml de los diferentes medios de cultivo. A cada uno se le adicionó un disco de micelio de P. nicotianae de 10mm de diámetro. Se colocaron 5 réplicas por cada tratamiento, que se mantuvieron en condiciones estáticas a temperatura de 25°C ± 2°C. A los 7 días el medio de cultivo con el microorganismo se filtró al vacío. La masa se colocó en la estufa a 60°C durante 7 días y se pesó en una balanza analítica.
A los filtrados se les determinó concentración de proteínas y la actividad de enzimas poligalacturonasas y xilanasas.
El contenido de proteínas (mg de proteína.g de tejido vegetal fresco-1) se determinó mediante la metodología micro- Lowry descrita por Sun (23).
La actividad poligalacturonasa se determinó con la adición de 100µl del filtrado a 400µl de polipectato de sodio al 0,1% en buffer acetato de sodio 50mmol.L-1, pH 5. La mezcla de reacción se incubó 1 hora a 37°C y se determinaron los azúcares reductores totales según Nelson (24), contra una curva patrón de galactosa de 1g.L-1 obtenida mediante la toma de alícuotas desde 20 y hasta 100 µl para un volumen final de 1ml. La actividad enzimática se expresó como microgramos (µg) de galactosa formados por mg de proteína.ml-1 del filtrado.
La actividad xilanasa se cuantificó con la adición de 100µl del extracto enzimático a 400µl de una solución de xilano al 0,1% en buffer acetato de sodio 50mmol.L-1, pH 5. La mezcla de reacción se incubó 1 hora a 37°C y la reacción se detuvo con la adición de mezcla cúprica alcalina de acuerdo al método de Nelson (24). Se determinaron los azúcares reductores totales contra una curva patrón de xilosa. La actividad enzimática se expresó como los µg de xilosa formados por mg de proteína.ml-1 del filtrado.
Los datos se procesaron estadísticamente mediante un ANOVA de clasificación simple, y previamente se evaluó la normalidad y homocedasticidad de estos. Las medias se compararon mediante la prueba de Tukey con un nivel de significación de 0.05, con el paquete estadístico SPSS 11.5 y los gráficos se confeccionaron con SigmaPlot 2001, versión 7.0.
RESULTADOS Y DISCUSIÓN
Crecimiento y esporulación de P. nicotianae en diferentes medios de cultivo sólidos ]]>
La dinámica de crecimiento de P. nicotianae en los diferentes medios de cultivo sólidos mostró que aquellos que contenían los extractos naturales o sus combinaciones con PDA, beneficiaron el crecimiento micelial del microorganismo (Figura 1). A los 6 días, el micelio en el medio con extracto de tabaco- asparagina y CaCO3 alcanzó el borde de la placa (90 mm), seguido por aquellos que contenían V8. A diferencia de estos, las colonias crecidas en los medios PDA y PDA- asparagina- CaCO3 a los 7 días apenas alcanzaron 2,5 y 4 cm, respectivamente, con diferencias estadísticamente significativas entre ellas, las que parecen estar relacionadas con la adición de asparagina y CaCO3 (1). Se comprobó que la velocidad de crecimiento en el medio PDA es lenta, comparada con otros con extractos naturales que le permiten un mejor desarrollo al microorganismo, aspecto señalado por diversos autores, a pesar de que dicho medio es el más común para el cultivo de hongos y oomycetes (1, 21, 22).Se observó que la velocidad de crecimiento del microorganismo fue superior en aquellos medios que contenían V8, en relación con los otros, excepto en el caso del extracto de tabaco, lo que permite un menor tiempo de duración de los experimentos y menor riesgo de contaminación de estos. Este medio combina 8 verduras que constituyen una variada fuente de nutrientes, elemento significativo si se tiene en cuenta que P. nicotianae es una especie con un amplia gama de hospedantes, que incluye cultivares tan diversos como: frutabomba, pimiento, areca, begonia, piña, limonero, aguacatero, plátano, tabaco y tomate, entre muchas otras. Por esta razón, algunos trabajos reflejan la importancia de medios de cultivo a base de tomate para el crecimiento y esporulación de especies de Phytophthora, particularmente frente a extractos de lo que serían sus hospedantes naturales. En tal sentido, los resultados de Cotilla et al. (22), muestran un mayor crecimiento de Phytophthora palmivora Butler en jugo de tomate con CaCO3, al igual que López et al. (21), en los medios V3, V8 y V10 comparados con PDA. Asimismo, Akinrefon (19), también comprobó que las cepas de P. palmivora crecían a mayor velocidad en medios como berenjena, cacao y tomate, mientras que en PDA tardaban mucho más tiempo en alcanzar el borde de la placa, resultados con los que coinciden los obtenidos en el presente estudio.
En los diferentes medios de cultivos, se observaron colonias blancas, las que, en la mayoría de los casos, presentaron bordes regulares, excepto cuando se cultivó el microorganismo en PDA, donde además de un crecimiento pobre, se observó el micelio adherido al sustrato. Las colonias de los medios sólidos que contenían jugo V8 se caracterizaron por ser algodonosas, pero las que se cultivaron en el extracto de tabaco tuvieron un crecimiento superficial y menos compacto. Esto pudo haberse debido a una menor ramificación de las hifas, y un crecimiento de estas hacia el frente, razón por la cual las colonias del medio de tabaco pudieron alcanzar el extremo de la placa en menor tiempo que aquellas que, al encontrarse en un medio rico, divergieron su micelio para obtener más nutrientes y se tornaron en colonias más densas.
Crecimiento y actividades enzimáticas en medios de cultivo líquidos
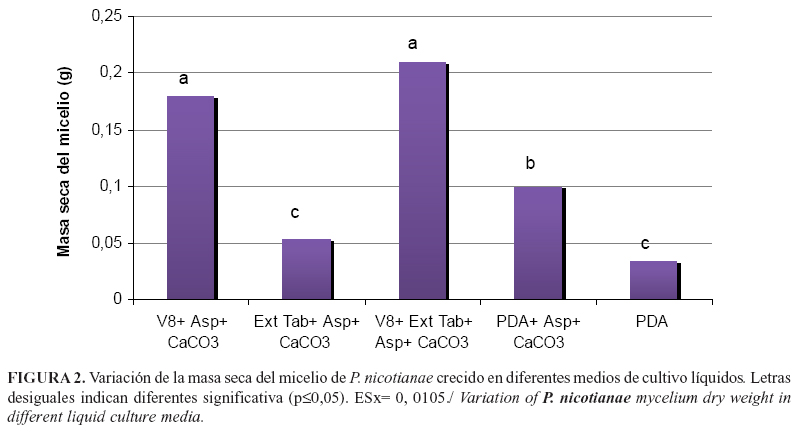
El crecimiento de P. nicotianae en medio líquido fue disperso. El mayor valor (0,21g) correspondió al medio con jugo V8 y no presentó diferencias significativas con el medio que contenía además el extracto de tabaco (Figura 2). Los medios menos favorables para el crecimiento fueron los que contenían el extracto de tabaco (0,054g) y PDA (0,034g). Particularmente, en este último se corroboró lo señalado por otros autores (1), acerca de la importancia de la asparagina y el CaCO3 como suplementos en el medio que benefician el desarrollo del microorganismo, al observarse diferencias estadísticamente significativas entre los medios que contenían o no estos nutrientes. Estos resultados, en general, muestran que aunque en medio sólido los tratamientos con extracto de tabaco alcanzaron rápidamente el borde de la placa, ello no se tradujo en un incremento de la biomasa. En medio sólido el crecimiento fue poco ramificado, con menor número de puntas hifales que en medio V8, y sí con una gran extensión de las apicales en busca de nutrientes, por lo que se alcanzó el extremo de la placa en menor tiempo que otros tratamientos. Es por ello, que es necesario realizar ensayos en cultivo líquido para comprobar un real incremento en la producción de micelio.
Los resultados del crecimiento en ambos experimentos confirman al medio de cultivo suplementado con jugo V8 como el más adecuado para el crecimiento de la cepa SS-11 de P. nicotianae, ya que en poco tiempo se produce abundante micelio y este alcanza el extremo de la placa, lo que resulta de importancia para futuros ensayos in vitro, como las relacionados con la evaluación de la capacidad antimicrobiana de algunos compuestos.
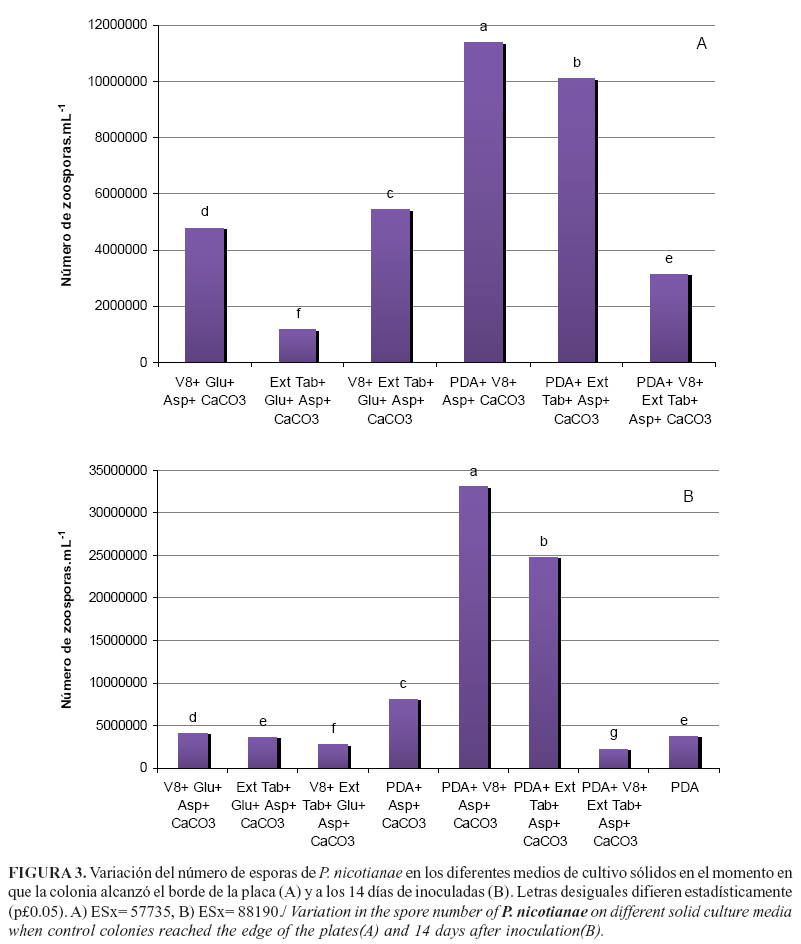
La determinación del número de esporas cuando cada colonia alcanzó el borde de la placa (Figura 3A) y a los 14 días de inoculadas muestra diferencias entre los tratamientos (Figura 3B). A los 8 días ya se había alcanzado el extremo de la placa en casi todos los medios de cultivo, excepto en PDA y su combinación con asparagina - CaCO3, por lo que en estos tratamientos se contó el número de esporas solo a los 14 días de inoculadas. Para ambos momentos de conteo se obtuvieron diferencias significativas entre los tratamientos.
El número de zoosporas producidas al momento en que las colonias alcanzaron el borde de la placa en los medios PDA-jugo V8 y PDA-extracto de tabaco fue casi dos veces superior, en relación a los valores del resto de los tratamientos. A su vez, la combinación PDA- jugo V8 fue mejor estadísticamente que la combinación PDA-extracto de tabaco. En los medios donde no se empleó PDA, la combinación jugo V8 con extracto de tabaco produjo mayor número de esporas.
El conteo de esporas a los 14 días corroboró que las combinaciones PDA-jugo V8 y PDA-extracto de tabaco inducen las mayores formaciones de zoosporas en P. nicotianae, respecto al resto de los tratamientos evaluados. Ambos conteos demostraron que la combinación de los extractos naturales con el medio semi-sintético PDA beneficia la esporulación y que en el medio PDA P. nicotianae puede esporular, pero en menor cantidad que en los demás. Estos resultados coinciden con los de otros autores relativos a la formación de esporangios en Phytophthora infestans (Mont.) de Bary cuando evaluaron medios de cultivo que incluían jugo de tomate; sin embargo, no observaron esporangios en PDA (21).
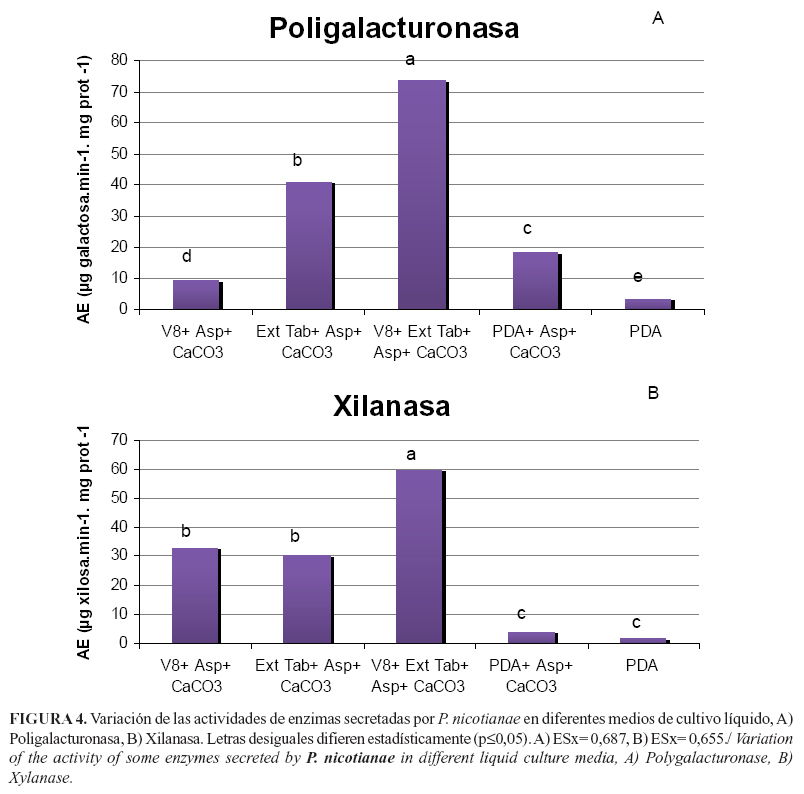
Las actividades enzimáticas determinadas en los filtrados mostraron que la combinación del jugo V8- extracto de tabaco fue la más eficaz para la expresión de las enzimas evaluadas, con diferencias significativas respecto a los otros tratamientos (Figura 4), lo que demuestra que la combinación de ambos sustratos naturales es más propicia para el despliegue del potencial enzimático del patógeno, que el medio PDA. ]]>
Los valores más elevados de actividad poligalacturonasa se encontraron en el medio donde se combinaron los compuestos naturales, seguidos del medio con extracto de tabaco, que difirió estadísticamente con el que incluye jugo V8 (Figura 4A).Los valores de actividad xilanasa en los medios con extracto de tabaco y con jugo V8, también fueron elevados respecto a los que contenían PDA y no difirieron estadísticamente entre sí (Figura 4B). Sin embargo, la combinación de ambos fue la que alcanzó los mayores valores de actividad enzimática casi duplicando los encontrados en cada uno de los medios naturales por separado.
Varios estudios relacionados también con la búsqueda de medios de cultivo que induzcan la actividad de estas enzimas fueron realizados por Cardinale y Matta (25), quienes compararon la inducción de xilanasa en Fusarium oxysporum Schltdl. con diferentes fuentes de carbono y detectaron que los mayores valores se obtenían en presencia de paredes celulares o de xilano. Asimismo, Akinrefon (19) determinó actividad poligalacturonasa en filtrados de P. palmivora cuando se incluyó pulpa de cacao, pero no cuando se empleó el medio PDA. Nuestros resultados coinciden con los este autor, lo que corrobora la especificidad de la enzima por el sustrato que coloniza. Estas diferencias observadas en las actividades enzimáticas en los diferentes medios de cultivo pudieran explicarse por el hecho que las enzimas poligalacturonasas son codificadas por familias multigénicas cuyo número varía con la especificidad de la interacción (3).
Según diferentes autores, la actividad de las enzimas evaluadas puede ser una medida de la patogenicidad de una cepa (9, 20), por lo que cobra importancia contar con un medio de cultivo que permita su mejor expresión, y por tanto, un empleo más efectivo del microorganismo al realizar ensayos de infección o protección de plantas contra este patógeno.
Generalmente, un medio de cultivo que favorezca el crecimiento de Phytophthora, no tiene por qué ser el más adecuado para la secreción de enzimas como poligalacturonasas y pectin metil esterasas (19). Sin embargo, se comprobó que los medios de cultivo que contienen V8, favorecen el crecimiento, esporulación y secreción de enzimas de P. nicotianae. Estos resultados son de gran utilidad, ya que ofrecen variantes con medios de cultivo que acortan el tiempo de experimentación, debido al aumento de la velocidad de crecimiento de este microorganismo. Además, inducen la formación de un mayor número de zoosporas y permite que la cepa exprese en mayor medida su potencial patogénico referido a enzimas poligalacturonasas y xilanasas.
REFERENCIAS
1. Erwin DC, Ribeiro OK. Phytophthora diseases worldwide. The American Phytopathological Society, St Paul, Minnesota, USA. 1996. 562 páginas.
2. Espino E. Instituto de Investigaciones del Tabaco. Manual Práctico del supervisor agrícola del tabaco. 2da Edición. 2006. 60 páginas.
3. Esquerré-Tugayé MT, Boudart G, Dumas B. Cell wall degrading enzymes, inhibitory proteins, and oligosaccharides participate in the molecular dialogue between plants and pathogen. Plant Physiol Biochem. 2000;38:157-163.
4. Hardham A. Cell biology of plant- oomycete interactions. Cellular Microbiology. 2007;9(1):31-39.
5. Tian MY, Win J, Song J, van der Hoorn R, van der Knaap E, Kamoun S. A Phytophthora infestans cystatin-like protein targets a novel tomato papain-like apoplastic protease. Plant Physiology. 2007;143:364-377.
6. Göhre V, Robatzek S. Breaking the barriers: Microbial effector molecules subvert plant immunity. Annual Review of Phytopathology. 2008;46:189-215.
7. Haas BJ, Kamoun S, Zody MC, Jiang RHY, Handsaker RE, Cano LM, et al. Genome sequence and analysis of the Irish potato famine pathogen Phytophthora infestans. Nature. 2009;461:393-398.
8. Avrova AO, Boevink PC, Young V, Grenville-Briggs L, van West P, et al. A novel Phytophthora infestans haustorium- specific membrane protein is required for infection of potato. Cellular Microbiology. 2008;10(11):2271-2284.
9. Jian Y, Zhen B, Xiu W, Guo X. Polygalacturonase, pectate lyase and pectin methylesterase activity in pathogenic strains of Phytophthora capsici incubated under different conditions. Journal of Phytopathology. 2009;157:585-591.
10.Al-Najada AR, Al-Hindi RR, Mohamed SA. Characterization of polygalacturonases from fruit spoilage Fusarium oxysporum and Aspergillus tubingensis. Afr J of Biotechnol. 2012;11(34):8527-8536.
11.Bhosale HJ, Sukalkar SR, Uzma SMZ, Kadam TA. Production of xylanase by Streptomyces rameus grown on agricultural wastes. Biotechnol Bioinf Bioeng. 2011;1(4):505-512.
12.Ahmed S, Riaz S, Jamil A. Molecular cloning of fungal xylanases: An overview. Applied Microbiology and Biotechnology. 2009;84(1):19-35.
13.Kamble RD, Jadhav AR. Xylanase production under solid state and submerged fermentation conditions by bacterial strains. African Journal of Microbiology Research. 2012;6(20):4292-4297.
14.Buga ML, Ibrahim S, Nok AJ. Partially purified polygalacturonase from Aspergillus niger (SA6). Afr J Biotechnol. 2010;9:8944-8954.
15.Dogan N, Tari C. Characterization of three-phase partitioned exo-polygalacturonase from Aspergillus sojae with uniqu properties. J Biochem Eng. 2008;39:43-50.
16.Damasio ARL, da Silva TM, Maller A, Jorge JA, Terenzi HF, Polizeli ML. Purification and partial characterization of an exo-polygalacturonase from Paecilomyces variotii liquid cultures. Appl Biochem Biotechnol. 2010;160:1496-1507.
17.Kamoun S. Molecular genetics of Pathogenic Oomycetes. Eukariotic Cell. 2003;2(2):191-199.
18. Yan HZ, Liou RF. Cloning and analysis of pppg1, an inducible endopolygalacturonase gene from the oomycete plant pathogen Phytophthora parasitica. Fung Genet Biol. 2005;42:339-350.
19.Akinrefon OA. Production of Extracellular Enzymes by Phytophthora palmivora Butl. J gen Microbiol. 1968;51:67-74.
20.Lalaoui F, Halama P, Dumortier V, Paul B. Cell wall-degrading enzymes produced in vitro by isolates of Phaeosphaeria nodorum differing in aggressiveness. Plant Pathology. 2000;49:727-733.
21.López M, Tomás T. Estudio de medios de cultivo sobre el crecimiento lineal y la esporulación de Phytophthora infestans. Fitosanidad. 1999;3(3):23-31.
22.Cotilla L, Díaz A, Berroa G, Rodríguez R. Influencia de cuatro medios de cultivo sobre el crecimiento micelial de Phytophthora palmívora (Butler) Butl. Centro Agrícola. 2007;34(2):49-52.
23.Sun SM. Methods in plant molecular biology and agricultural biotechnology: A laboratory training manual, Asian Research and Development Center. Tainan, Taiwan, 1994, 94p.
24.Nelson N. A photometric adaptation of the Somogyi method for the determination of glucose. J Biol Chem. 1944;153:375-380.
25.Cardinale F, Matta A. Basic xylanases from the fungal tomato pathogen Fusarium oxysporum esp. lycopersici. Journal of Plant Pathology. 2001;83:27-36.
Recibido: 1-11-2013. ]]>
Aceptado: 6-1-2014. ]]>
{kind=link}
{kind=link}
{kind=link}