{kind=link}
Diversidad genética de aislados de Ralstonia solanacearum de Cuba, mediante amplificación de las regiones repetitivas del genoma (REP-PCR)
Genetic diversity among Cuban strains of Ralstonia solanacearum assessed by repetitive sequence-based polymerase chain reaction (REP-PCR)
IDirección de Sanidad Vegetal. Centro Nacional de Sanidad Agropecuaria (CENSA). Carretera de Tapaste y Autopista Nacional, San José de Las Lajas, CP 32 700, Mayabeque, Cuba.
IICentro Nacional de Sanidad Vegetal, Ayuntamiento # 231 e/ San Pedro y Lombillo, Plaza de la Revolución. La Habana. Cuba.
]]>
RESUMEN
Se estableció la diversidad genética de 30 aislados cubanos de Ralstonia. solanacearum, obtenidos de papa y tomate y pertenecientes a los biovares 1 y 2, mediante la amplificación de las secuencias consenso repetitivas intergénicas de enterobacterias (rep-PCR) con los cebadores ERIC. El método permitió distinguir 11 genotipos entre los aislados cubanos. Las agrupaciones primarias alcanzadas, mediante el análisis de conglomerados, se correspondieron con el biovar de los aislamientos, mientras que las subdivisiones encontradas dentro de cada grupo, tuvieron relación con el hospedero del cual fueron obtenidos. Los aislados de papa mostraron un coeficiente de similitud del 37,2% y la presencia de variantes específicas para cada región geográfica dentro de cada biovar. Los aislados de tomate registraron una similitud del 71,3%, con la presencia cruzada del mismo genotipo en diferentes regiones geográficas y la incidencia de diferentes variantes genéticas en una misma región. El Análisis de Componentes Principales explicó el 81,32% de la variabilidad de los aislados, lo que corroboró las agrupaciones de acuerdo al hospedero y el biovar, y permitió establecer relaciones según la ubicación geográfica. Las distribución diferencial de la diversidad genética entre los aislados cubanos de R. solanacearum, que se obtuvieron de los cultivos de papa y tomate, puede ser una herramienta útil para el establecimiento de medidas específicas para el control de la Marchitez Bacteriana en Cuba.
Palabras clave: Ralstonia solanacearum, diversidad genética, rep-PCR, papa, tomate.
ABSTRACT
The genetic diversity among 30 Cuban strains of Ralstonia solanacearum isolated from potato and tomato and belonging to biovars 1 and 2, was assessed by the repetitive sequence based polymerase chain reaction (rep-PCR) method with ERIC primer sets. The method defined 11 genotypes from all the strains. The primary groupings obtained by cluster analysis corresponded with the strain biovar, while the subdivisions found within each group were related to the host from which the strains were obtained. Potato isolates showed a 37.2% similarity and the presence of specific variants within each biovar for each geographic region. Isolates from tomato showed a similarity of 71.3 % with the cross presence of the same genotype in different geographic regions and the incidence of different genetic variants in the same region. The Principal Component Analysis explained 81.32 % of isolate variability, which corroborated the groupings according to the host and biovar and allowed to establish relationships based on the geographic location. The differential distribution of the genetic diversity among Cuban R. solanacearum strains from potato and tomato crops may be useful information to establish specific measures to control the bacterial wilt disease in Cuba.
Key words: Ralstonia solanacearum, genetic diversity, rep-PCR, potato, tomato.
INTRODUCCIÓN ]]>
La marchitez bacteriana causada por Ralstonia. solanacearum (Smith) Yabuuchi, es una enfermedad que se encuentra distribuida a nivel mundial y afecta a cientos de especies en 54 familias de plantas, entre las que se encuentran cultivos de importancia económica como tomate (Solanum lycopersicum L.), papa (Solanum tuberosum L.), plátano (Musa sp.), tabaco (Nicotiana tabacum L. , entre otros (1,2). Las afectaciones provocadas por este patógeno radican en el hecho de que destruye los cultivos, y posee una excepcional habilidad para sobrevivir en el agua, el suelo y la rizosfera de las plantas no hospederas (3,4).Debido a su elevado número de hospedantes, R. solanacearum se considera un complejo de especies, formado por un grupo heterogéneo de aislados con características comunes (5). La tipificación en razas y biovares fue la primera clasificación informal empleada a nivel subespecífico dentro de R. solanacearum. La denominación en razas se basa en la gama de hospedantes, mientras que las diferencias en la actividad metabólica, se emplean para definir seis biovares (6). Posteriormente, el complejo de especies quedó subdividido en cuatro grupos genéticos (Filotipos), de acuerdo a los análisis de las secuencias de los genes hrp, de la endoglucanasa y del ARNr16S, los cuales se corresponden con el origen geográfico primario de los aislados analizados (7).
Con el desarrollo de las técnicas moleculares, numerosas metodologías fueron utilizadas en el estudio de las relaciones intraespecíficas dentro de R. solanacearum. Entre estas se encuentran los análisis del polimorfismo de la longitud de los fragmentos de restricción y amplificación (RFLP y AFLP, respectivamente) (8, 9) y el análisis de los perfiles obtenidos con la amplificación de las regiones repetitivas del genoma (rep-PCR) (10, 11), entre otras. Los patrones logrados con estos marcadores permiten establecer agrupaciones en correspondencia con características como la raza, el biovar o el origen geográfico de los aislados (10, 11).
La marchitez bacteriana se encuentra ampliamente distribuida en todas las islas del Caribe y países del continente americano (12). Las condiciones climáticas presentes en Cuba son propicias para el desarrollo y supervivencia de R. solanacearum, a lo que se suma el riesgo de introducción de cepas foráneas por la importación de semillas procedentes de diferentes países (13).
Cuba cuenta con métodos de detección e identificación, basados en el aislamiento de la bacteria en medios de cultivos y la caracterización bioquímica. Además, se emplean, como técnicas rápidas de detección, la inmunofluorescencia indirecta (IFI) y la Reacción en Cadena de la Polimerasa (PCR) (13); sin embargo, no se han desarrollado métodos para la caracterizac ión molecular de los aislados presentes en el país.
El objetivo de este trabajo fue evaluar la diversidad genética de 31 aislados cubanos de R. solanacearum, basado en la amplificación de las regiones repetitivas del genoma rep-PCR, específicamente de las secuencias consenso repetitivas intergénicas de enterobacterias (ERIC-PCR).
MATERIALES Y MÉTODOS
Aislados empleados y obtención de muestras para la PCR
Para este estudio se utilizaron 31 aislados cubanos pertenecientes al cepario del Laboratorio Central de Cuarentena Vegetal (LCCV) y ADN de una cepa de referencia de Taiwán (Tabla 1). La caracterización en biovares fue previamente realizada, según lo descrito por Denny y Hayward (14); las preparaciones de ADN se realizaron de acuerdo a lo establecido por Seal et al. (15). ]]>
La amplificación de las secuencias repetitivas del genoma (rep-PCR) se realizó con los iniciadores ERIC1R (5´-ATGTAAGCTCCTGGGGATTCAC-3´) y ERIC2 (5´- AAGTAAGTGACTGGGGTGAGCG-3´). La mezcla de reacción se conformó por tampón de PCR 5x, 25mM de MgCl2, 25mM de cada dNTP, 30pmol de cada iniciador y 2 unidades de Taq polimerasa, para un volumen final de 25µl. El programa de amplificación constó de un ciclo a 95 °C durante 7 min, seguido de 30 ciclos a 94 °C durante 1 min, 52 °C durante 1 min, 65 °C durante 8 min y un ciclo final de extensión a 65 °C durante 15 min. Las amplificaciones se repitieron al menos dos veces por aislado para garantizar la reproducibilidad de los patrones de bandas obtenidos.Se tomaron 12 ul de cada producto de amplificación, se aplicaron en un gel de agarosa al 1,5 % en tampón TBE al 0,5 % y se empleó un patrón de 250 pares de bases Invitrogen®. La corrida se realizó, aproximadamente, durante dos horas, al cabo de este tiempo se tiñó con bromuro de etidio y el resultado se visualizó en un transluminador de luz UV (Macrovue, LKB).
Con el objetivo de lograr un agrupamiento genético de los aislados, a partir de los patrones de bandas, se construyó una matriz binaria (0 ausencia - 1 presencia de banda). Los patrones de bandas repetidos se tomaron como único genotipo para la construcción de una nueva matriz, con la que se realizó un análisis de conglomerados. En el análisis se empleó la matriz de similitud de DICE (16) y el método UPGMA. Se utilizaron los software DendroUPGMA (17) y MEGA 5.0 (18).
La distribución de los aislados, según biovar, hospedante de origen y zona geográfica de procedencia, se obtuvo mediante el gráfico biplot, proveniente del Análisis de Componentes Principales. Las variables (patrón de bandas en código binario) fueron previamente estandarizadas y se empleó la matriz de correlaciones. Este análisis se realizó con el sistema STATISTICA, versión 7.0 (19).
RESULTADOS
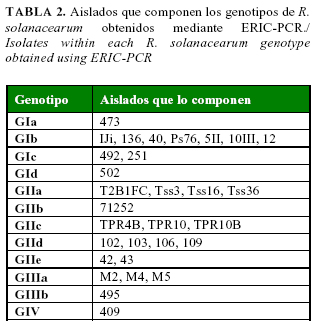
La amplificación de las secuencias repetitivas del genoma, con el empleo de los cebadores ERIC, permitió la amplificación de 12 bandas polimórficas y dio lugar a la formación de 12 genotipos (Tabla 2). Se obtuvieron resultados reproducibles con todos los aislados, excepto con Col 7, para el cual no se logró un patrón repetitivo. Seis de los genotipos fueron aislados de tomate, cinco aislados de papa, y el genotipo restante representó la cepa de referencia 409, procedente de Taiwán.
En el dendrograma se obtuvieron 4 grupos principales (Fig. 1): en los Grupos I y II se agruparon aislados procedentes de papa y tomate, pertenecientes al biovar 1; el Grupo III estuvo conformado por aislados obtenidos de papa pertenecientes al biovar 2, y en el Grupo IV quedó ubicada la cepa de referencia de Taiwán (biovar 4). Las divisiones primarias observadas en los agrupamientos se correspondieron con el biovar de los aislados analizados, mientras los subgrupos obtenidos dentro de cada uno de estos conjuntos principales, se correspondieron con los hospederos de origen.
Los aislados cubanos pertenecientes al biovar1 manifestaron la mayor diversidad, pues se agrupan a 9 de los 11 genotipos cubanos, por lo que muestra el 60,4% de coeficiente de similitud; sin embargo, los dos aislados del biovar 2, obtenidos de papa, formaron dos genotipos con una similitud entre ellos del 90,9%. El coeficiente medio de similitud entre los aislados de tomate fue del 71,3 %, mientras que el registrado entre los aislados obtenidos de papa fue del 37,2 %. ]]>
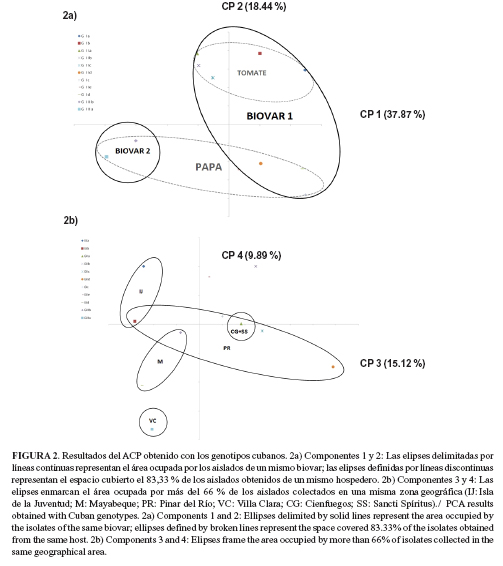
Los aislados de papa fueron divididos de forma primaria de acuerdo al biovar, mientras que las subdivisiones en genotipos estuvieron en correspondencia con la provincia en la cual se obtuvieron. Los aislados de tomate mostraron una distribución más compleja con la presencia del mismo genotipo en diferentes ubicaciones geográficas (genotipo GIb en la Isla de la Juventud y Pinar del Río; Genotipo GIIa en Cienfuegos y Sancti Spírutis) y la incidencia de diferentes variantes genéticas en una misma región (genotipos GIb, GIc, GIIb, GIIc y GIId en Pinar del Río) (Figura 1).Los gráficos construidos mediante el ACP se manifiestan en la figura 2. Con el uso de los cuatro primeros componentes (ejes 1 y 2; 3 y 4) fue posible explicar la diversidad encontrada entre los perfiles obtenidos por ERIC-PCR en el 81,32 % del total.
Con el trazo de los ejes (componentes 1 y 2), las agrupaciones más claras correspondieron, al igual que para el análisis de conglomerados, a las divisiones en biovares y hospederos de procedencia. Con el trazo de los ejes 3 y 4, los genotipos fueron agrupados en función de la ubicación geográfica. La mayor dispersión en los puntos se observó para la provincia de Pinar del Río, con solapamientos en las áreas descritas por los genotipos de provincias como Mayabeque e Isla de la Juventud, y con regiones del centro del país, como Cienfuegos y Sancti Spíritus. El genotipo formado por los aislados de Villa Clara no mostró intersecciones con el resto de las variantes.
DISCUSIÓN
La técnica de rep-PCR ha sido empleada por varios autores para el estudio de la diversidad genética de R. solanacearum con resultados variables. Horita y Tsuchiya (10) hallaron que los principales grupos obtenidos con aislados en Japón, mediante el empleo de esta técnica, se correspondieron con las divisiones en razas, mientras que Xue et al. (11) encontraron cierta correlación con la zona de origen y el hospedero de aislados de R. solanacearum en China. Norman et al. (20) pudieron subdividir un grupo de aislados de R. solanacearum, logrados a partir de material vegetal de importación a los Estados Unidos, de acuerdo al biovar y con una menor correlación de acuerdo al hospedero y al origen geográfico. Lemessa et al. (21), por su parte, encontraron dos grupos principales con esta técnica entre los aislados de R. solanacearum en Etiopía, los cuales correspondían a los biovares que circulaban en el país.
En este estudio, la técnica de ERIC-PCR permitió una división primaria de acuerdo al biovar de los aislados y de subdividir los principales grupos obtenidos de acuerdo al hospedero. Por otra parte, el ACP reveló relaciones desde el punto de vista espacial entre los genotipos, aunque al igual que los resultados obtenidos por otros autores, las divisiones de acuerdo al origen geográfico no fueron claras para la totalidad de las muestras (11, 20). No obstante, el solapamiento observado en el ACP de los genotipos en Pinar del Río con los genotipos de otras provincias, sugiere que esta región representa la zona de mayor prevalencia y diversidad de la bacteria y como posible punto de diseminación del patógeno hacia otras regiones del país, aspecto que requiere de estudios posteriores.
Una mayor diversidad genética se registró entre los aislados cubanos pertenecientes al biovar 1, mientras que los aislamientos del biovar 2, obtenidos únicamente de papa, mostraron patrones muy similares. Las cepas del biovar 1 se agrupan en la raza 1 de R. solanacearum que afecta el mayor grupo de hospedantes de la especie, mientras que las cepas del biovar 2 se concentran en la raza 3 y comprenden una gama de hospedantes limitada, principalmente a solanáceas (1, 20). Esta pudiera ser la razón por la cual el biovar 2 de R. solanacearum está expuesto a una mayor presión selectiva (20) y por lo que otros autores describieron, a una menor diversidad genética entre los aislados que lo componen en comparación con el biovar 1 (10, 20).
La distribución geográfica cruzada, mostrada por los genotipos obtenidos de tomate, sugiere que el patógeno puede diversificarse localmente en este cultivo en diferentes genotipos y que también es capaz de diseminarse de forma clonal entre provincias colindantes. Este fenómeno puede estar asociado con las prácticas agrícolas de baja intensidad empleadas en este cultivo, las cuales, junto al comercio y movimiento de las cosechas, contribuyen a una rápida diversificación y diseminación de los patógenos de plantas (22). Por otra parte, las limitadas diversificación y diseminación geográfica del patógeno a partir de los linajes aislados de papa, pueden ser consecuencias del manejo intensivo de este cultivo en el país y de una mayor eficacia en las medidas de control, en comparación con el cultivo del tomate.
Estudios más profundos con el empleo de otros marcadores, o mediante el análisis de la secuencia de genes conservados como el ARNr16S o el gen de la endoglucanasa, deberán ser asumidos en el futuro, a fin de esclarecer el origen geográfico primario de los aislados en estudio (7). ]]>
La técnica de rep-PCR continúa siendo utilizada con éxito para el estudio de la diversidad genética de varias especies de bacterias fitopatógenas (23, 24, 25). Es fácil de usar y puede ser aplicada a un gran número de muestras (26). Aunque emplea cierto tiempo para la obtención de los resultados, no requiere de la secuenciación, por lo que constituye una metodología útil para los estudios de diversidad genética en los países en vías de desarrollo (25).El número de aislados estudiados no es suficiente para arribar a conclusiones definitivas sobre la diversidad de las poblaciones de R. solanacearum en Cuba. No obstante, sienta las bases para los futuros estudios de la diversidad genética del patógeno en la isla, al mismo tiempo que provee una valiosa información sobre el establecimiento de las medidas de control de la Marchitez Bacteriana en el país.
REFERENCIAS
1. Álvarez MB. Biology of Ralstonia solanacearum Phylotype II in host and non-host environments. (PhD Thesis), Instituto Valenciano de Investigaciones Agrarias, Valencia, España. 2009.
2. Cellier G, Prior P. Deciphering Phenotypic Diversity of Ralstonia solanacearum strains pathogenic to potato. The American Phytopathological Society. 2010;100(11):1250-1261.
3. Carmeille A, Prior P, Kodja H, Chiroleu F, Luisetti P, Besse P. Evaluation of Resistance to Race 3, Biovar 2 of Ralstonia solanacearum in Tomato Germplasm. J Phytopathology. 2003;154(3):398-402. ]]>
4. González I, Arias Y, Peteira B. Plant-phytopathogen bacteria interaction: case study Ralstonia solanacearum-host plants. Rev Protección Veg. 2009;24(2):61-80.
5. Ji P, Allen C, Sanchez-Perez A, Yao J, Elphinstone JG, et al. New diversity of Ralstonia solanacearum strains associated with vegetable and ornamental crops in Florida. Plant Dis. 2007;91:195-203.
6. Schaad NW, Jones JB, Chun W. Ralstonia solanacearum. In: Plant Pathogenic Bacteria. 3rd Edition. Editorial APS Press. 2001; 151-162.
7. Fegan M, Prior P. How complex is the Ralstonia solanacearum species complex? En: Allen C, Prior P and Hayward AC editors. Bacterial wilt disease and the Ralstonia solanacearum species complex. American Phytopathological Society Press, St. Paul, MN. 2007; 449-461.
8. Cook D, Sequeira J. Strain differenciation of Pseudomonas solanacearum by molecular genetics methods. En: Hayward AC and Hartman GL. Bacterial wilt: The disease and the causative agent. Pseudomonas solanacearum. Editorial Willingford CAB international. 2004; 77-93. ]]>
9. Dookun A, Saumtally Seal S. Genetic Diversity in Ralstonia solanacearum Strains from Mauritius using Restriction Fragment Length Polymorphisms. J Phytopathology. 2001;149:51-55.
10.Horita M, Tsuchiya K. Genetic diversity of Japanese strains of Ralstonia solanacearum. Phytopathology. 2001;91:399-407.
11.Xue QY, Yin YN, Yang W, Heuer H, Prior P, Guo JH, et al. Genetic diversity of Ralstonia solanacearum strains from China assessed by PCR-based fingerprints to unravel host plant- and site-dependent distribution patterns. FEMS Microbiol Ecol. 2011;75(3):507-519.
12.Wicker E, Grassart L, Coranson-Beaudu R, Mian D, Guilbaud C, et al. Ralstonia solanacearum Strains from Martinique (French West Indies) exhibiting a new pathogenic potential. Appl Environ Microbiol. 2007;73(21):6790-6801.
13.Iglesia A, Alvarez E, Martínez Y, García A. Diagnóstico molecular de la Marchitez Bacteriana. Rev Protección Veg. 2008;23(2):75-79. ]]>
14.Denny TP, Hayward AC. Gram Negative Bacteria. In: Schaad NW, Jones JB and Chun W. Plant Pathogenic Bacteria. 3rd Edition. Editorial APS Press. 2001; p.564.
15.Seal SE, Jackson LA, Young JPW, Daniels MJ. Differentiation of Pseudomonas solanacearum, Pseudomonas syzygii, Pseudomonas pickettii and the blood disease bacterium by partial 16S rRNA sequencing: Construction of oligonucleotide primers for sensitive detection by polymerase chain reaction. Journal of General Microbiology. 1993;139:1587-1594.
16.Dice LR. Measures of the amount of ecologic association between Species. Ecology. 1945;26(3):297-302.
17.Garcia-Vallve S, Palau J, Romeu A. Horizontal gene transfer in glycosyl hydrolases inferred from codon usage in Escherichia coli and Bacillus subtilis. Molecular Biology and Evolution. 1999;9:1125-1134.
18.Tamura K, Peterson D, Peterson N, Stecher G, et al. MEGA5: Molecular Evolutionary Genetics Analysis using Likelihood, Distance, and Parsimony methods. Molecular Biology and Evolution. 2011;0:0. ]]>
19.StatSoft, Inc. STATISTICA (data analysis software system), version 7. http://www.statsoft.com. 2004.
20.Norman DJ, Zapata M, Gabriel DW, Duan YP, Yuen JMF, Mangravita-Novo A, et al. Genetic diversity and host range variation of Ralstonia solanacearum strains entering North America. Phytopathology. 2009;99:1070-1077.
21.Lemessa F, Zeller W, Negeri D. Genetic diversity among strains of Ralstonia solanacearum from Ethiopia assessed by repetitive sequence-based polymerase chain reaction (rep-PCR). EJAST. 2012; 1(1): 17-26.
22.Parkinson N, Cowie C, Heeney J, Stead D. Phylogenetic structure of Xanthomonas determined by comparison of gyrB sequences. International Journal of Systematic and Evolutionary Microbiology. 2009;59:264-274.
23.Louws FJ, Bell J, Medina-Mora CM, Smart CD, Opgenorth D, Ishimaru CA, et al. rep-PCR-Mediated Genomic Fingerprinting: A Rapid and Effective Method to Identify Clavibacter michiganensis. Phytopathology. 1998;8:862-864. ]]>
24.Lewis I, McSpadden ML, Gardner BB, Opina N, Miller SA. Diversity and geographic distribution of Ralstonia solanacearum from eggplant in the Philippines. Phytopathology. 2007;97:1467-1475.
25.Singh D, Dhar S, Yadava DK. Genetic and pathogenic variability of Indian strains of Xanthomonas campestris pv. campestris causing black rot disease in crucifers. Curr Microbiol. 2011;63(6):551-560.
26.Olive DM, Dean P. Principles and Applications of Methods for DNA-Based Typing of Microbial Organisms. J Clin Microbiol. 1999;37(16):1661.
Recibido: 16-11-2014.
Aceptado: 6-2-2015. ]]>
*Autor de contacto: Yamila Martínez Zubiaur. E-mail: yamila@censa.edu.cu. ]]>
{kind=link}
{kind=link}