
Diversidad genética de aislados de Ralstonia solanacearum procedentes de tres regiones de Colombia
Genetic diversity of Ralstonia solanacearum isolates from three Colombian regions
Carolina Cardozo BurgosI, Bernardo Silva AguilarI, Mauricio Salazar YepesII, Juan Gonzalo Morales OsorioIII*
IUniversidad Nacional de Colombia sede Palmira, Facultad de Ciencias Agropecuarias, Departamento de Ciencias Agrícolas, Carrera Ingeniería Agronómica, Carrera 32 No 12 - 00 Chapinero, Vía Candelaria, Palmira - Valle del Cauca - Colombia.
IIUniversidad Nacional de Colombia sede Medellín, Facultad de Ciencias, Escuela de Biociencias. Calle 59A No. 63-20, Medellín-Colombia, Sur América. ]]>
IIIUniversidad Nacional de Colombia sede Medellín, Facultad de Ciencias Agrarias, Departamento de Ciencias Agronómicas, Bloque 11. Calle 59A No. 63-20, Medellín-Colombia, Sur América. Tel: 0057 4 4309099, Fax: 0057 4 4309118.
RESUMEN
Este trabajo se realizó con el objetivo de analizar la diversidad genética del complejo de especies Ralstonia solanacearum en tres regiones de Colombia donde se siembran cultivos hospedantes de gran importancia económica para el país. Se colectaron 69 aislamientos procedentes de suelo y diferentes hospedantes. Para determinar la diversidad genética entre los aislados se utilizaron los marcadores moleculares del tipo microsatélite amplificados al azar (RAMs). Los datos se analizaron utilizando el coeficiente de Nei-Li, el análisis de correspondencia múltiple (ACM) y el índice de Shannon. Los valores del coeficiente de Nei-Li, a un nivel de similaridad de 0,76%, permitieron agrupar los aislamientos en tres divisiones. El ACM arrojó un valor de similitud del 67% al interior de 10 grupos y del 62% entre los grupos. El Coeficiente de diferenciación genética (Gst) mostró un valor de 0,13 e indicó que existe mayor diversidad genética de los aislados dentro de cada grupo que cuando se comparan los diferentes grupos entre sí. El valor del índice de Shannon corresponde con los otros índices y muestra que los aislados en cada grupo presentan una diversidad mayor que la que existe entre los grupos. La interpretación del valor de Gst interpoblacional (0,0514) sugiere una diferenciación genética moderada de la población. Los resultados indican o señalan alto flujo de genes entre las poblaciones.
Palabras clave: Ralstonia solanacearum, diversidad genética, RAMs.
ABSTRACT
The present work was performed to analyze the genetic diversity of the R. solanacearum species complex in three Colombian regions where economically important host crops are grown. Sixty nine isolates were collected from different hosts and soils. The genetic diversity was determined using Random Amplified Microsatellites (RAMs) as molecular markers . The data were analyzed by the Nei and Li coefficient, Multiple Correspondence Analysis (MCA), and the Shannon index. The Nei and LI coefficient values found at a similarity level of 0.76% allowed grouping the isolates into three major divisions.. The MCA showed a similarity coefficient of 67% within 10 groups and 62% between groups. The Genetic differentiation coefficient (Gst) showed a value of 0.13 indicating that the genetic diversity was higher within each group of isolates than when the different groups were compared one another. The Shannon index value corresponded with the other calculated indices, indicating that the isolates within each group showed a higher genetic diversity than that observed between the different groups. The Inter-population Gst value (0.0514) suggested a moderate genetic differentiation in the population. The results suggested a high gene flow between populations.
Key words: Ralstonia solanacearum, genetic diversity, RAMs.
]]>
INTRODUCCIÓN
La marchitez bacteriana causada por Ralstonia solanacearum (Smith 1896, Yabuuchi et al., 1996) es una enfermedad que provoca grandes pérdidas económicas a nivel mundial (1). Esta enfermedad está ampliamente distribuida en las regiones tropicales, subtropicales y templadas del mundo. Se conoce que esta bacteria afecta a más de 250 especies de plantas, clasificadas dentro de 54 familias botánicas diferentes (2). Se informó la presencia de 28 nuevos hospedantes para este patógeno en trabajos realizados durante los últimos 10 años en Colombia, lo que sugiere una elevada adaptabilidad del microorganismo (2). Esta especie bacteriana se clasifica en 5 razas, según la patogenicidad y el rango de plantas hospedantes que afecta (2). La clasificación en Biovares se basa en la capacidad metabólica de utilizar tres disacáridos (cellobiosa, lactosa y maltosa) y tres alcoholes hexosas (dulcitol, manitol y sorbitol) (2,3). La raza 1 está principalmente relacionada con plantas pertenecientes a la familia Solanaceae; la raza 2 afecta las familias Musaceae y Heliconiaceae en las que causa marchitez bacteriana en plátanos triploides y heliconias en áreas tropicales, la enfermedad es denominada Moko bacteriano; la raza 3 afecta algunas Solanaceas y geranios, principalmente en zonas templadas o de clima frío moderado tropical ; la raza 4 afecta preferentemente el cultivo del jengibre (1,2,3); la raza 5 se especializa en plantas del género Morus sp. (4). Numerosos hospedantes de este patógeno se cultivan en Colombia, entre estos se encuentran la papa, el tomate, las hortalizas, las plantas ornamentales como las heliconias y los cultivos como el banano y el plátano: estos dos últimos constituyen renglones importantes en la economía y en la alimentación del país. Adicionalmente, en Colombia se encuentra la mayor diversidad de heliconias en el mundo (5), por lo que la bacteria es también una amenaza para la biodiversidad y su conservación en bancos de germoplasma.
El complejo de especies R. solanacearum presenta gran diversidad a nivel mundial (6,7). Las clasificaciones basadas en razas y biovares no son completamente exactas, ya que algunas cepas que ocasionan la enfermedad del Moko bacteriano en Musáceas pueden afectar también el tomate y la papa (2,3,8,9). Por tanto, los problemas presentes en el manejo de la enfermedad y en los programas de mejoramiento para obtener cultivos resistentes se debe a la amplia diversidad y a la distribución geográfica del agente etiológico, así como a la alta capacidad de adaptación a diferentes ambientes y hospedantes (2,3,6,10), lo que implica que se deben conocer estos aspectos básicos para diseñar estrategias apropiadas para el manejo de la enfermedad.
Las técnicas moleculares permiten diferenciar los grupos de acuerdo a las características como son el origen geográfico o el hospedante (11,12,13). Los aislados de esta bacteria se clasificaron con la utilización de diversos tipos de marcadores moleculares: RFLP's, secuencias de regiones intergénicas ITS y de los genes hrpB, AFLP's, análisis de secuencia del RNAr 16S y Endoglucanasa (egl) (12,14,15). Sobre la base de estos marcadores moleculares se definieron los términos «filotipo» para referirse a los grupos subespecíficos y «sequevar» para definir los grupos infrasubespecíficos. Los estudios que se basaron en análisis de secuencias de varios loci (MLSA) permitieron identificar ocho grupos de cepas que presentaron diferentes patrones evolutivos, distribuidos dentro de los cinco filotipos previamente clasificados (16). También se logró determinar la diversificación,la estructura poblacional recientemente expandida, el alto nivel de recombinación homóloga y la evidencia de migraciones a larga distancia (16). Fegan (17) informó que los aislamientos de la raza 2 de R. solanacearum están asociados al filotipo II y que las cepas evaluadas MLG 24, 25 y 28 correspondían a los secuevares 3, 4 y 6, respectivamente.
En Colombia se realizaron estudios de las poblacionales de R. solanacearum utilizando marcadores moleculares. Para identificar los aislamientos de R. solanacearum de musáceas, colectados en diferentes regiones del país, Gómez et al. (18) usaron la secuenciación del ADN ribosomal 16S. En las regiones productoras de banano de Urabá-Colombia y Magdalena se caracterizaron molecularmente los aislados de R. solanacearum procedentes de suelo, plantas de bananos y de arvenses; para ello se utilizó PCR múltiple y secuenciación de los genes Endoglucanasa (egl), la subunidad del flagelo (fliC) y la región 16S del ADNr; se encontró que los aislados provenientes de Urabá pertenecían al filotipo II, secuevar 4 y los del Magdalena al secuevar 6 (19). Sin embargo, algunos asilamientos no se pudieron clasificar en ningún secuevar conocido para la raza 2, lo que indica la necesidad de continuar con estudios de caracterización molecular usando otros marcadores (19). Los estudios previos de caracterización molecular de R. solanacearum raza 2, en Colombia, se han realizado principalmente con aislados de la región de Urabá (19,20), por lo que se hace necesario extender los estudios a otras regiones colombianas productoras de cultivos hospedantes de la bacteria.
El objetivo del presente trabajo fue determinar la variabilidad genética de aislamientos de R. solanacearum colectados en diferentes regiones de Colombia, utilizando marcadores moleculares del tipo Amplificación aleatoria de microsatélites (RAMs).
MATERIALES Y MÉTODOS
Aislamientos de R. solanacearum ]]>
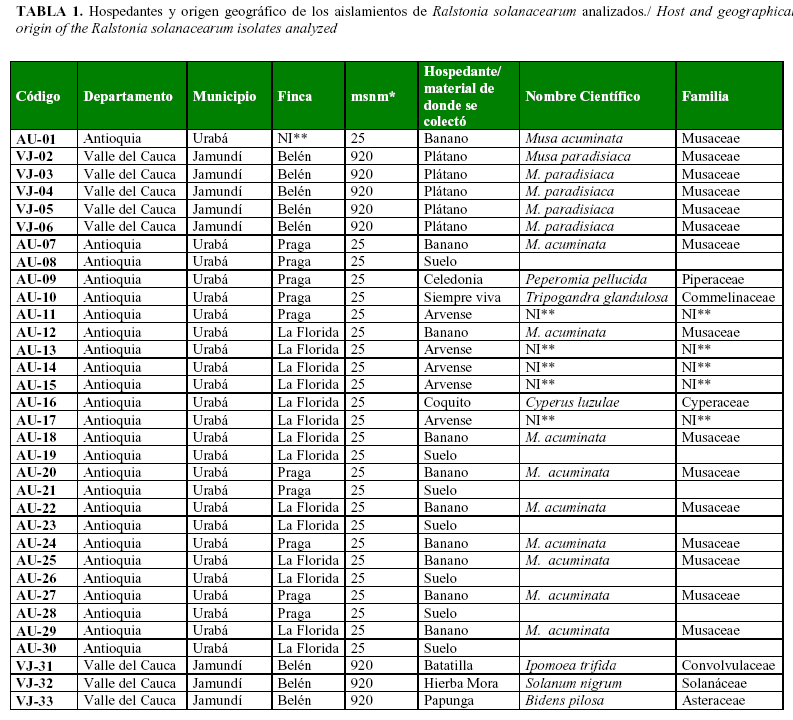
Se colectaron muestras vegetales en plantaciones comerciales, plantas arvenses y suelo asociado a estas, en los departamentos colombianos de Antioquia (región Urabá), Quindío y Valle del Cauca (Tabla 1a y 1b), regiones todas productoras de banano, plátano, tomate, heliconias y otras plantas hospedantes de este patógeno. Se colectaron plantas clasificadas en 13 familias botánicas, 23 géneros y 32 especies. Las muestras se envolvieron en servilletas de papel y se guardaron en neveras de icopor a temperatura ambiente, para el envío al Herbario MEDEL de la Universidad Nacional de Colombia sede Medellín donde se identificaron.Para obtener el aislamiento de la bacteria se utilizó el medio semiselectivo Kelman (TZC), según el método descrito por Prieto et al. (2). Para ello se tomó parte del tallo y la raíz de las plantas colectadas, se desinfestaron con etanol (70%) por 30 segundos; luego se lavaron con agua destilada, se sumergieron por 30 segundos en una solución de hipoclorito de sodio (0,5%), se lavaron con agua destilada estéril, se secaron con una toalla estéril; en cámara de flujo laminar se cortaron pequeñas porciones de 1mm2 aproximadamente y se sembraron secciones que contenían los haces vasculares en placas Petri con el medio semiselectivo.
Las muestras de suelo se procesaron siguiendo el método informado por Pradhanang et al. (21). Las placas se incubaron a 28ºC por dos días y se seleccionaron las colonias características de color vino tinto con presencia de un halo dorado alrededor, bien definido en forma de «ojo de pescado», tal y como se describió previamente para R. solanacearum (2).
Los aislamientos obtenidos se confirmaron como la especie R. solanacearum, para ello se usó el kit comercial ImmunoStrip® test Kit de Agdia® y se siguieron las instrucciones del fabricante.
Las cepas de R. solanacearum seleccionadas se cultivaron en 1,5 ml de medio líquido (CPG) estéril compuesto por Casaminoácido 1 g, Peptona 10 g, Glucosa 4 g por litro de medio y se mantuvieron a 22ºC y 250 rpm, por 18 horas. Se tomaron 700 µl de cultivo bacteriano, se mezclaron suavemente con 300 µl de glicerol estéril y se almacenaron a -80°C para su uso posterior.
Extracción de ADN
Los aislamientos se cultivaron en 1,5ml del medio líquido descrito previamente y se mantuvieron a 22ºC y 250 rpm por 15 horas (2). Para extraer el ADN, las células bacterianas se colectaron por centrifugación a 11000 rpm por 4 minutos. El pellet se resuspendió en 500µL de Buffer TE 1X (10mM de Tris, 1mM de EDTA pH 8.0); se adicionaron 15 µl de SDS al 20% y tres microlitros de proteinasa K. La mezcla obtenida se agitó en vortex y se incubó a 56ºC por una hora, agitando en vortex cada 5 minutos. Luego, se adicionaron 100 µl de NaCl 5 M, se mezcló suavemente por inversión y se añadieron 80 µl de CTAB/NaCl (10% CTAB / 0.7M NaCl) mezclando suavemente. Se incubaron nuevamente a 65ºC por diez minutos. Se colocaron en hielo por 15 minutos, se adicionaron 200 µl de la mezcla cloroformo-alcohol isoamílico (24:1), se incubaron a -20ºC por diez minutos y se centrifugaron a 13000 rpm por diez minutos.
El sobrenadante se transfirió a un nuevo tubo eppendorf y se adicionaron 800 µl de etanol al 96%, se mezcló suavemente y se incubaron a -20ºC por una hora. Luego se centrifugó a 13000 rpm por diez minutos. El precipitado se dejó secar por 15 minutos a temperatura ambiente y se añadió 50 µl de Buffer TE 1X. Luego se adicionaron tres microlitros de RNAsa (10 mg.ml-1) (Fermentas) a cada muestra y se guardaron a 4ºC.
Para determinar la calidad y concentración del ADN se tomaron dos microlitros de ADN y se visualizaron por irradiación de luz ultravioleta en un transiluminador (KODAK EDAS 290) sobre un gel de agarosa al 0,8% en TBE al 0,5 X teñido con bromuro de etidio (0,5 ng.ml-1). La concentración de ADN presente en cada muestra se estimó por comparación con patrones de peso molecular de ADN del bacteriófago lambda, con concentraciones finales de 20, 40, 60, 80 y 100 ng. El ADN cuantificado se diluyó en agua para HPLC hasta una concentración final de 5 ng.µl-1 y se almacenó a -20ºC para uso posterior.
Amplificación de RAMs ]]>
La amplificación de los fragmentos de ADN con cebadores diseñados para los marcadores RAM, se realizó en un termociclador PTC-100 (MJ Research). La reacción en cadena de la polimerasa (PCR) se realizó en un volumen final de 25 µl, las concentración finales fueron: 1X buffer Taq (Fermentas); 0,2 mM de cada dNTP; 2 µM de cada cebador; 1,5 mM de MgCl2; 20 ng de ADN para cada aislamiento; 1 U de Taq polimerasa y H2O hasta completar 25 µl.Para la amplificación de los fragmentos se utilizaron las siguientes condiciones: desnaturalización inicial a 95ºC por 5 min, posteriormente 37 ciclos de una desnaturalización a 95 ºC por 30 s, hibridación de cebadores a 50ºC por 45 s y extensión a 72ºC por 2 min; finalmente se hizo una extensión a 72ºC por 7 min. En el diseño de los cebadores se agregó una región degenerada adicional (Tabla 2).
Los fragmentos amplificados de ADN se separaron por electroforesis en geles de agarosa (2%) preparados en solución tampón TBE 0.5 X, se visualizaron bajo luz ultravioleta, tal y como se describió. Se utilizó un marcador de peso molecular de 100pb (Fermentas).
Designaciones para los sitios degenerados: D (G, A, T); B (G,T,C); H (A,T,C); V (G,A,C); Y (C,T).
Análisis de los datos
Para cada perfil electroforético se registró la presencia o ausencia de bandas y las bandas en común, en una matriz binaria de datos, en la cual los aislamientos se organizaron en filas y, las bandas identificadas, en columnas (22).
Los análisis se realizaron utilizando el programa de computación NTSYS-pc versión 2.02 (Numerical Taxonomy System for Personal Computer). Se realizaron el Análisis de similitud según el Coeficiente de Nei-Li (22), el Análisis de Correspondencia Múltiple y el Índice de Shannon. La matriz de similitud se construyó con el programa SIMQUAL del paquete NTSYS-pc versión 2.02 (22). Con la matriz se obtuvo una de agrupamiento con el programa SAHN de NTSYS-pc y se usó el método UPGMA (Unweighted pair-group arithmetic mean); a partir de la misma se construyó un dendrograma con el programa TREE del paquete NTSYS-pc versión 2.02 (22). Se emplearon los valores de Bootstrap (basados en 500 re-muestreos) para estimar la confiabilidad del patrón de agrupamiento. El análisis de correspondencia múltiple se realizó con el método CMA, del programa NTSYS-pc versión 2.02 (22). Como haplotipos diferentes se definieron aquellos patrones que se diferenciaron, al menos, en una banda.
Análisis de Similaridad de Nei-Li
Sij =2a/(2a+b+c) ]]>
Sij es la similaridad entre dos individuos i y j; a es el Número de bandas presentes simultáneamente en ambos i y j; b es el Número de bandas presentes en i y ausentes en j; c es el Número de bandas presentes en j y ausentes en i. Las bandas que coincidieron se promediaron por un factor de 2 para alcanzar una mejor diferenciación de los individuos con bajo nivel de similitud (22).El cálculo de la diversidad, de acuerdo con los haplotipos encontrados en cada grupo y en toda la población, se realizó a través de la siguiente fórmula:
H= 1 - + f (i)2
Donde f(i) es la frecuencia del patrón de la población; + f (i) 2 es la probabilidad de que los individuos tomados al azar tengan el patrón i; H es la probabilidad de que los individuos tomados al azar tengan diferente patrón. H es el valor con el que se presenta la diversidad de la población.
La diversidad para subgrupos de cualquier naturaleza se calculó mediante la fórmula:
Donde n es el número de individuos totales; Ht es la diversidad total; HS es la diversidad dentro de cada subgrupo o subpoblación; HSt es la diversidad entre subgrupos o subpoblaciones.
La mayor diversidad genética posible en cada subgrupo se estimó mediante el coeficiente de diferenciación genética GST.
Índice de diversidad de Shannon (intrapoblacional) ]]>
El índice de diversidad de Shannon se estimó con el programa Popgene32 versión 1.31 (23) y se calculó con la fórmula:Donde Pij es la frecuencia del carácter i (presencia, i=1 o la ausencia de la banda i=0) en la población j y m es el número de caracteres (bandas o loci) analizado. Su valor mínimo es cero y se hace mayor al aumentar la diversidad (23).
El nivel de distribución de la diversidad genética total se calculó mediante el índice GST, equivalente multialélico de FST (22). Los valores que se obtuvieron para GST (FST) se interpretaron siguiendo las pautas recomendadas por Wright (24) para el equivalente FST: un rango de 0,0 a 0,05 indica poca diferencia entre poblaciones; de 0,05 a 0,15, diferenciación genética moderada; de 0,15 a 0,25, diferenciación genética alta; valores superiores a 0,25, diferenciación genética muy alta.
RESULTADOS
Aislamientos
Se obtuvo 69 aislamientos positivos para R. solanacearum, a partir de muestras de plantas de 13 familias botánicas diferentes y de suelo (Tabla 1a y 1b). Posteriormente, se procedió a realizar el análisis de diversidad genética de los aislados.
Selección de cebadores
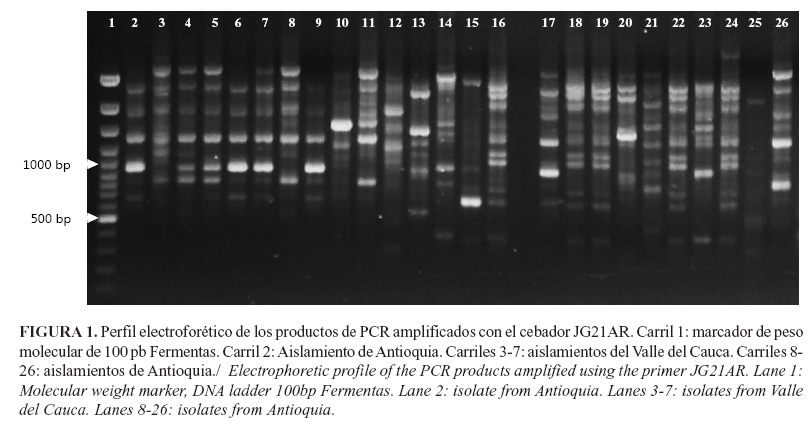
De seis cebadores evaluados inicialmente, se seleccionaron cinco, los que proporcionaron un patrón de banda polimórfico y reproducible (Tabla 2, Fig. 1). Con estos cebadores se identificaron 43 bandas polimórficas con tamaños de 250 a 1200pb, en las diferentes cepas bacterianas. La comparación de los patrones de bandas identificadas se usó para la construcción de la matriz binaria de datos que se utilizó para los cálculos de diversidad genética, el Análisis de Correspondencia Múltiple y el Índice de Shannon. ]]>
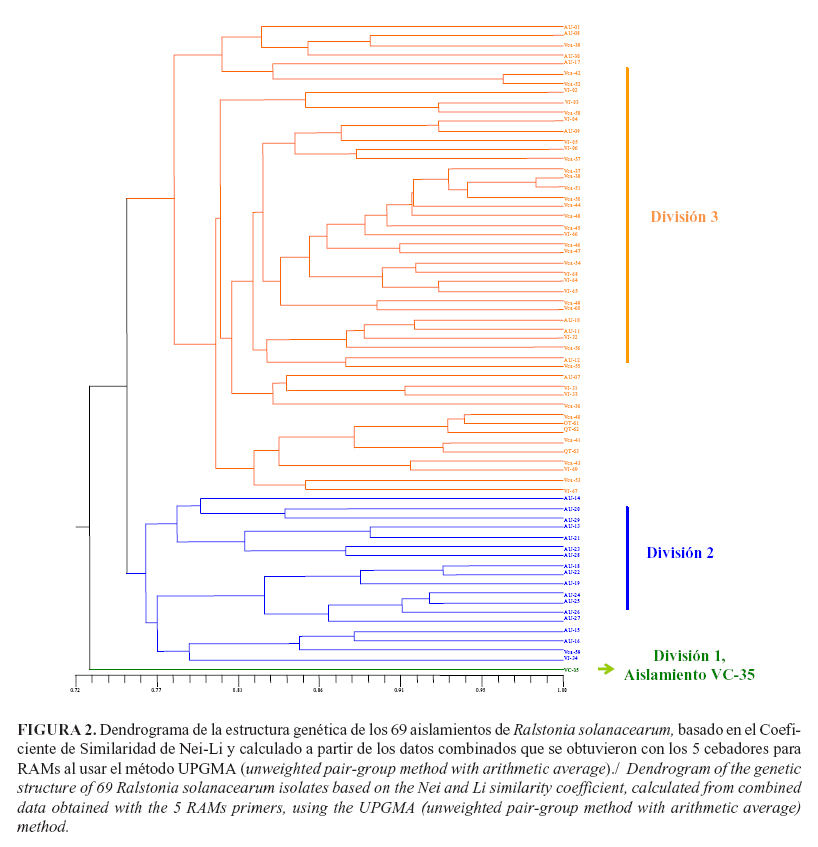
Análisis de la diversidad genéticaLos análisis realizados, a partir del el coeficiente de Nei-Li (22) con un nivel de similitud de 0,76% entre los aislados, mostró tres divisiones principales (Fig. 2). En la primera división se identificó solo un aislamiento (VC-35) proveniente de S. lycopersicum (tomate), usualmente clasificado como raza 1, colectado a 920 metros sobre el nivel del mar (msnm) en el municipio de Pradera en el departamento del Valle del Cauca (Tabla 1a y 1b). En la segunda división se encontraron 18 aislamientos, 16 de ellos colectados en la región de Urabá, departamento Antioquia, donde se cultiva banano y plátano de exportación, así como dos aislamientos provenientes del Departamento del Valle del Cauca: el aislamiento VCa-59 proveniente de Heliconia bihai, colectado a 1600 msnm. y el aislamientoVJ-34, aislado de plátano, cultivar Dominico Hartón y cultivado a 920 msnm (Tabla 1a y 1b). En la tercera división se localizaron 50 aislamientos de diferentes hospedantes y de suelo, 9 provenientes de la región Urabá-Antioquia, 38 colectados en el departamento del Valle del Cauca y 3 del sur del Departamento del Quindío. Los municipios de Caicedonia que se localizan en el norte del Valle del Cauca y La Tebaida en el sur del Quindío, presentan límites geográficos comunes. Por esta razón, algunos sitios de colecta podrían estar localizados a muy corta distancia relativa entre sí, pero ubicados en diferentes departamentos. La proximidad de los sitios de colecta es un factor que se debe considerar al realizar el análisis de diversidad por zonas geográficas.
Análisis de Correspondencia Múltiple
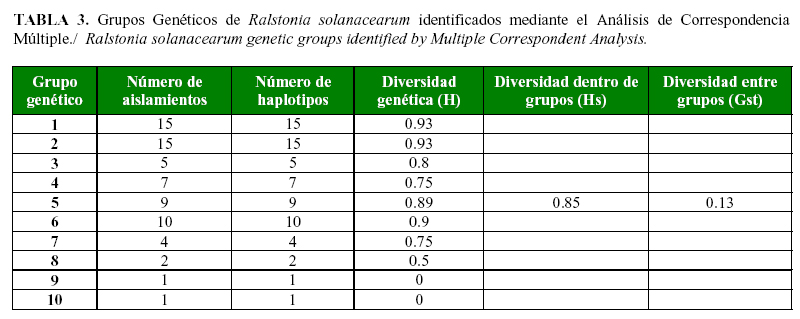
El Análisis de Correspondencia Múltiple (ACM) generó diez grupos diferentes con un coeficiente de similitud del 67% dentro de grupos y del 62% entre los grupos (Tabla 3). Cuatro de los diez grupos se conformaron por 11 aislamientos provenientes de la región de Urabá, ubicada en el departamento Antioquia. Cuarenta y tres aislamientos colectados en los departamentos Antioquia y Valle del Cauca conformaron cinco grupos diferentes. En el último grupo se ubicaron los tres aislamientos obtenidos en el departamento del Quindío, junto con 11 muestras provenientes del Valle del Cauca y una de Antioquia. El coeficiente de diferenciación genética (GST) arrojó un valor de 0,13, lo que indica que los individuos son más diversos dentro de un mismo grupo que entre los grupos identificados en este análisis (Tabla 3).
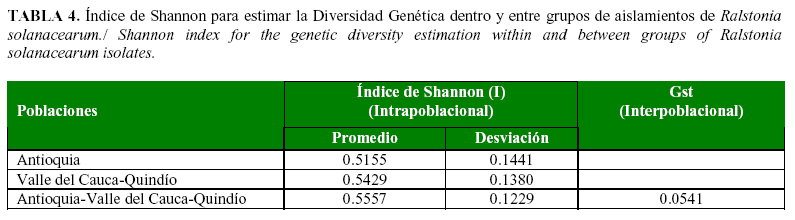
Índice de Shannon
Los valores obtenidos con el Índice de Shannon (Tabla 4) sugieren que la población estudiada es más diversa dentro de los grupos que entre ellos. El grupo de aislamientos colectados en el Valle del Cauca-Quindío es más diverso (0,5429) que el grupo de Antioquia (0,5155). La interpretación del índice GST inter-poblacional (0,0514) sugiere que la población tiene una diferenciación genética moderada (24). Este corresponde con los valores obtenidos con el coeficiente de Nei-Li en el análisis de la diversidad genética (22), los cuales indican que la identidad de la población estudiada es del 0,9460 y que la distancia entre la misma es de 0,0555. La diversidad encontrada para la población total es más alta que las que presentaron las localidades en forma individual.
DISCUSIÓN
El índice de diversidad genética de Nei-Li permitió identificar tres grupos genéticos (Fig. 2). El primer grupo constituido por un aislamiento de tomate, planta no reportada como hospedante frecuente de R. solanacearum raza 2, sino de las razas 1 y 3. Este aislamiento es diferente genéticamente a los otros dos grupos. Además, este posee los determinantes de patogenicidad para inducir síntomas de Moko en plátano.
Los estudios realizados por Fegan y Prior (25), basados en la secuencia del gen que codifica para la enzima endoglucanasa (egl), mostraron que los aislados que causaban la enfermedad del Moko bacteriano en plátano estaban muy relacionados con aislados patogénicos en tomate y papa, por lo que se demuestra que los mismos presentan un origen polifilético. Asimismo, Albuquerque et al. (8) informaron un comportamiento similar entre aislamientos de R. solanacearum provenientes de Brasil, entre los cuales uno fue patogénico a banano y tomate, otro a banano y papa. Prior y Fegan (26) comunicaron sobre los ecotipos de R. solanacearum raza 2, los cuales produjeron síntomas característicos de Moko en Musa spp. en condiciones de campo y tuvieron la habilidad de ser patogénicos en tomate y en otras solanáceas, si se inoculaban en los tallos. Estos resultados indican que el rango de hospedantes de la bacteria podría ser más amplio de lo que se conoce tradicionalmente y que no se debe subestimar la importancia de estos hallazgos, ya que las prácticas agronómicas inadecuadas pueden causar heridas en raíces o pseudotallos de banano y plátano que simulan las condiciones de inoculación artificial; de este modo se contribuye a diseminar e inducir la enfermedad entre los cultivos. ]]>
En el segundo agrupamiento se encontraron aislamientos principalmente del departamento del Valle del Cauca y del sur del departamento del Quindío. El norte del Valle del Cauca limita con el sur del Quindío. Aunque presentan límites de ordenamiento político, desde el punto de vista biológico y epidemiológico es más relevante el hecho de que comparten la misma zona de vida, lo que explicaría el nivel de similitud genética observado. Este aspecto es muy importante en los estudios epidemiológicos y de variabilidad, ya que con frecuencia se determinan los agrupamientos genéticos basados en los límites geopolíticos. Sin embargo, los organismos no dependen exclusivamente de estos, sino de las variables que influyen en la generación de variabilidad, como son el tipo de suelo y el material parental, los regímenes de precipitación, la humedad relativa, la luminosidad, la temperatura, la altura sobre el nivel del mar (sobre todo en los trópicos), los hospedantes presentes en la zona, los cultivos y las prácticas que se realizan, entre otras.Los resultados obtenidos con el análisis de correspondencia múltiple sugieren que existe un alto flujo de genes entre las diferentes áreas evaluadas. Varios factores podrían determinar el grado de flujo de genes identificado, entre ellos se encuentra el movimiento de material vegetal infectado, el cual contribuye a la diseminación de la enfermedad, la coexistencia de hospedantes prácticamente en toda la región Andina, las corrientes de agua abundantes en la zona y la rotación de cultivos con otros hospedantes de la bacteria, entre otros. La comunicación de toda la región y el movimiento de inóculo generan una baja posibilidad de que se presente una zona donde pueda existir aislamiento geográfico. La clasificación por razas, por lo general, relaciona la cepa con un tipo de hospedante; sin embargo, se ha informado infección de plátano por aislamientos de tomate de la raza 1, lo que explica que los aislamientos provenientes de diferentes hospedantes puedan agruparse en los análisis realizados.
Los estudios previos de variabilidad para aislamientos de R. solanacearum obtenidos de suelo, arvenses y plantas de banano provenientes de la región de Urabá-Colombia, usando marcadores AFLP y el índice de Shannon, mostraron resultados similares a los alcanzados en este estudio (I=0,48) (20). El mayor valor encontrado para el índice de Shannon en el grupo de aislamientos del Valle del Cauca-Quindío, respecto al grupo de Antioquia, probablemente es debido a que en los sitios de colecta localizados en el Valle del Cauca-Quindío se encuentra una mayor variedad de pisos térmicos y zonas de vida, suelos y hospedantes (Tabla 4). En este grupo están incluidas las muestras del departamento del Quindío y el aislamiento VC-35 aislado de tomate, el cual constituyó una división independiente según el coeficiente de Nei-Li (22) (Fig. 2). Este valle interandino está delimitado por las cordilleras Central y Occidental, con una zona plana ubicada alrededor de los 1000 msnm, a partir de la cual se elevan las dos cordilleras hacia el occidente y hacia el oriente, alcanzando alturas hasta de 4000 msnm, lo cual determina diversas zonas de vida. Los suelos varían enormemente, se encuentran suelos aluviales y suelos propios de laderas en zonas con cenizas volcánicas. En esta región se cultiva una gran diversidad de plantas hospedantes de R. solanacearum y se identificaron varias especies de malezas hospedantes de la bacteria (2). Estos factores, junto con el cultivo simultáneo de varios hospedantes diferentes en los mismos lotes, podría contribuir a generar y mantener una mayor diversidad genética de las poblaciones del patógeno, comparado con la región muestreada en Urabá, ubicada en el norte de Antioquia, Colombia, donde predomina el monocultivo del banano en alrededor de 30 000 h, ubicadas predominantemente en una planicie aluvial a 50 msnm, en la zona de vida bosque húmedo tropical (bh-T) sensu Holdridge. El norte del Valle del Cauca limita con el sur del Quindío, lo cual explica que ambos grupos de aislamientos tengan una diversidad genética similar. Esto se debe a que se colectaron en la misma zona de vida sensu Holdridge.
Al analizar los diferentes valores de diversidad genética que presentaron las localidades se obtiene como resultado una mayor diversidad total. A ello contribuyen varios factores, como son las diferentes zonas de cultivo y el distanciamiento de las regiones geográficas de colecta, principalmente Urabá, Antioquia y Valle del Cauca Quindío, además de las diferentes familias, géneros y especies de hospedantes muestreados; así como otros factores edafo-climáticos, los cuales, de forma combinada, generan presiones de selección positiva para la adaptación de cada aislamiento a su entorno.
Los marcadores RAMs se han utilizado ampliamente para cuantificar la variabilidad genética en diversos organismos, incluso los microorganismos fitopatógenos como R. solanacearum (22,27). Estos marcadores moleculares son reproducibles, económicos y de baja complejidad técnica, por lo que constituyen una herramienta apropiada para la caracterización genética de poblaciones de R. solanacearum.
AGRADECIMIENTOS
Los autores agradecen a Andrés Mauricio Posso, por su ayuda para el análisis informático, al profesor Luis Fernando Patiño Hoyos por el suministro y la identificación de aislamientos. Este trabajo fue financiado por SENA-COLCIENCIAS proyecto: Development of a method of asymptomatic detection of the banana and plantain Moko disease PCR-based and applications to disease management, código: 8242-07-16025; Universidad Nacional de Colombia sede Medellín y sede Palmira (DIPAL) y al herbario MEDEL por la identificación de las plantas hospedantes.
REFERENCIAS ]]>
1. Peeters N, Guidot A, Vailleau F, Valls M. Ralstonia solanacearum, a widespread bacterial plant pathogen in the post-genomic era. Mol Plant Pathol. 2013;14(7):651-662.
2. Prieto J, Morales J, Salazar M. Identification of new hosts for Ralstonia solanacearum (Smith) race 2 from Colombia. Rev Protección Veg. 2012;27(3):151-161.
3. Genin S. Molecular traits controlling host range and adaptation to plants in Ralstonia solanacearum. New Phytol. 2010;187:920-928.
4. EPPO. Ralstonia solanacearum. OEPP/EPPO Bull. 2004;34:173-178.
5. Díaz J, Ávila L, Oyola J. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Sondeo del mercado internacional de Heliconias y Follajes Tropicales. Bogotá, Colombia; 2002. p. 32-33. ]]>
6. Huet G. Breeding for resistances to Ralstonia solanacearum. Front Plant Sci. 2014;5(715):1-5.
7. Safni I, Cleenwerck I, De Vos P, Fegan M, Sly L, Kappler U. Polyphasic taxonomic revision of the Ralstonia solanacearum species complex: proposal to emend the descriptions of Ralstonia solanacearum and Ralstonia syzygii and reclassify current R. syzygii strains as Ralstonia syzygii subsp. syzygii subsp. nov., R. solanacearum phylotype IV strains as Ralstonia syzygii subsp. indonesiensis subsp. nov., banana blood disease bacterium strains as Ralstonia syzygii subsp. celebesensis subsp. nov. and R. solanacearum phylotype I and III strains as Ralstonia pseudosolanacearum sp. nov. Int J Syst Evol Microbiol. 2014;64:3087-3103.
8. Albuquerque G, Santos L, Felix K, Rollemberg C, Silva A, Souza E, et al. Moko Disease-Causing Strains of Ralstonia solanacearum from Brazil Extend Known Diversity in Paraphyletic Phylotype II. Phytopathol. 2014;104(11):1175-82.
9. Deberdt P, Guyot J, Coranson-Beaudu R, Launay J, Noreskal M, Rivière P, et al. Diversity of Ralstonia solanacearum in French Guiana Expands Knowledge of the Emerging Ecotype. Phytopathol. 2014;104(6):586-596.
10.Asi Nion Y, Toyota K. Recent Trends in Control Methods for Bacterial Wilt Diseases Caused by Ralstonia solanacearum. Microb Environ. 2015;30(1):1-11. ]]>
11.Xue Q, Yin Y, Yang W, Heuer H, Prior P, et al. Genetic diversity of Ralstonia solanacearum strains from China assessed by PCR-based fingerprints to unravel host plant- and site-dependent distribution patterns. FEMS Microb Ecol. 2011;75:507-519.
12.Cellier G, Remenant B, Chiroleu F, Lefeuvre P, Prior P. Phylogeny and population structure of brown rot- and Moko disease-causing strains of Ralstonia solanacearum phylotype II. Appl Environ Microb. 2012;78(7):2367-2375.
13.N'guessan C, Brisse S, Le Roux-Nio A, Poussier S, Koné D, Wicker E. Development of variable number of tandem repeats typing schemes for Ralstonia solanacearum, the agent of bacterial wilt, banana Moko disease and potato brown rot. J Microbiol Methods. 2013;92(3):366-74.
14.Poussier S, Trigalet-Demery D, Vandewalle P, Goffinet B, Luisetti J, Trigalet A. Genetic diversity of Ralstonia solanacearum as assessed by PCR-RFLP of the hrp gene region, AFLP and 16S rRNA sequence analysis, and identification of an African subdivision. Microb. 2000;146:1679-1692.
15.Cellier G, Prior P. Deciphering phenotypic diversity of Ralstonia solanacearum strains pathogenic to potato. Phytopathol. 2010;100:1250-1261. ]]>
16.Wicker E, Lefeuvre P, Cambiaire J, Lemaire C, Poussier S, Prior P. Contrasting recombination patterns and demographic histories of the plant pathogen Ralstonia solanacearum inferred from MLSA. The ISME J. 2012;6:961-974.
17.Fegan M. Bacterial wilt diseases of banana: evolution and ecology. En: Allen C, Prior P, Hayward A (Eds.), Bacterial Wilt Disease and the Ralstonia solanacearum Species Complex. APS Press, St. Paul. Minnesota, EEUU; 2005. p. 379-386.
18.Gómez E, Álvarez E, Llano G. Identificación y caracterización de cepas de R. solanacearum raza 2, agente causante del Moko de plátano en Colombia. Fitopatol Col. 2004;28(2):71-75.
19.Cardozo C, Rodríguez P, Marín M. Caracterización molecular del complejo de especies Ralstonia solanacearum en la zona bananera de Urabá. Agron Col. 2009;27(2):203-210.
20.Cardozo C, Rodríguez P, Cotes J, Marín M. Variabilidad genética de la bacteria Ralstonia solanacearum (Burkholderiales: Burholderiaceae) en la zona bananera de Urabá (Colombia). Int J Trop Biol. 2010;58(1):31-44. ]]>
21.Pradhanang P, Elphinstone J, Fox R. Sensitive detection of Ralstonia solanacearum in soil: a comparison of different detection techniques. Plant Pathol. 2000;49:414-422.
22.Morillo A, Morillo Y, Hernando E, Ávila I. Molecular characterization of the plum collection (Prunus domestica (L.) Borkh) of the Pedagogical and Technological University of Colombia. Afr J Biotechnol. 2015;14(3):257-263.
23.Yeh C, Yang R, Boyle T. University of Alberta, POPGENE32 version 1.31, Windows-based software for population genetic analyses. Canada; 1997.
24.Felsenstein J. University of Washington, Theoretical Evolutionary Genetics, Genome 562. Seattle, Washington; 2015.
25.Fegan M, Prior P. Diverse members of the Ralstonia solanacearum species complex cause bacterial wilts of banana. Australas Plant Pathol. 2006;35:93-101. ]]>
26.Prior P, Fegan M. Diversity and Molecular detection of Ralstonia solanacearum Race 2 Strains by Multiplex PCR. En: Allen C, Prior P, Hayward A (Eds.), Bacterial Wilt Disease and the Ralstonia solanacearum Species Complex. APS Press, St. Paul. Minnesota, EEUU; 2005. p. 405-414.
27.CIAT. Cassava and tropical fruit pathology. Activity 6. Identification of RAMS markers to evaluate genetic diversity of selected isolates of Ralstonia solanacearum obtained from plantain. Integrated pest and disease management in major agroecosystems. Project- PE1. Annual report, CIAT, Cali, Colombia; 2003. p. 127-129.
Recibido: 2-4-2015.
Aceptado: 20-6-2015.
]]>
* Autor para correspondencia: Juan Gonzalo Morales Osorio. Correo electrónico: jgmoraleso@unal.edu.co. ]]>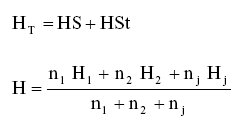
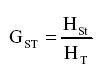
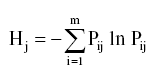
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}