Diferencias en la fagocitosis de Mycobacterium ´habana´ TMC 5135 en cultivos celulares de macrófagos murinos
Differences in phagocytosis of Mycobacterium ´habana´ TMC 5135 by murine macrophages cellular cultures
Iliana Valdés-Hernández,1* Jany Valdés-Martínez,1 Hermis Rodríguez-Sánchez, 2 Arianna Yglesias-Rivera,3 Alexis Díaz-García,3 Miguel Echemendía-Font1
1 Laboratorio Nacional de Referencia de Tuberculosis, Lepra y Micobacterias. Centro Colaborador OPS/OMS. Centro de Investigación, Diagnóstico y Referencia (CIDR). Instituto de Medicina Tropical ¨Pedro Kourí¨. La Habana, Cuba.
2 Laboratorio de Cultivos Celulares. Departamento de Sistemas Críticos. Centro de Investigación, Diagnóstico y Referencia (CIDR). Instituto de Medicina Tropical ¨Pedro Kourí¨. La Habana, Cuba.
]]> 3 Laboratorios de Producciones Biológico-Farmacéuticas (LABIOFAM). La Habana, Cuba.
email: iliana@ipk.sld.cu
* Licenciada en Microbiología. Máster en Bacteriología-Micología. Doctor en Ciencias de la Salud. Investigador Auxiliar. Profesor Auxiliar.
RESUMEN
La tuberculosis es uno de los mayores problemas que enfrenta la salud mundial en la actualidad. La única vacuna disponible contra esta enfermedad es la BCG, esta protege solo contra la tuberculosis grave de la infancia, lo cual plantea un reto en la búsqueda de nuevos candidatos vacunales. Teniendo en cuenta el antecedente protector de Mycobacterium ´habana´ contra la tuberculosis experimental, nos propusimos aportar elementos que avalen el empleo de M. ´habana´ TMC 5135 como candidato vacunal contra la tuberculosis, mediante estudios de infección en cultivos celulares de macrófagos murinos. Se caracterizó el proceso de fagocitosis de esta micobacteria por cultivos primarios de macrófagos peritoneales murinos y por la línea celular RAW 264.7, para lo cual se determinó el porcentaje de fagocitosis y el número fagocítico. El presente trabajo demostró que el proceso de fagocitosis de M. ´habana´ TMC 5135 está influenciado por la fuente celular empleada como célula hospedadora, así como por la carga bacteriana infectante y el tiempo de exposición a la misma. La presente investigación contribuye a la caracterización de la infección por esta micobacteria en sus principales células blanco de la inmunidad innata y traza el camino de futuras investigaciones para evaluar la activación de mecanismos efectores de la inmunidad innata frente a este candidato.
Palabras clave: Mycobacterium ´habana´, vacuna, tuberculosis, macrófagos.
ABSTRACT
Tuberculosis remains as a major problem in the global health. BCG is the available vaccine against tuberculosis but only protects against severe form of disease during childhood, so the search for new vaccine candidates is a challenge. Taking into account the protective capacity of Mycobacterium ´habana´ against experimental tuberculosis, we proposed in vitro experiments using murine macrophages (peritoneal macrophages and cell line Raw 264.7) to characterize phagocytic process of this candidate. Phagocitic index and phagocytic number were calculated. The present work demonstrated that the phagocytosis process of M. habana TMC 5135 is influenced by the cellular source used as host cell, as well as by the infecting bacterial load and the time of exposure. The present investigation contributes to the characterization of the infection by this mycobacteria in its main target cells of innate immunity and it suggest future investigations to evaluate the activation of effector mechanisms of the innate immunity against this candidate.
]]> Keywords: Mycobacterium ´habana´, vaccine, tuberculosis, macrophages.
INTRODUCCIÓN
La vacuna de Calmette-Guérin (BCG) es la única disponible para la prevención de la tuberculosis (TB). Su principales ventajas se asocian a su un bajo costo y amplio historial de seguridad, a lo que se suma su eficacia protectora de más de un 80% para las formas graves de la enfermedad en la infancia. Sin embargo, estudios demuestran que la protección que confiere es transitoria, en un rango de 10-20 años y no brinda protección contra la enfermedad pulmonar (1).
Estos inconvenientes han sido la causa del aumento en la búsqueda de nuevas alternativas de vacunación contra la TB. Estas van desde vacunas de subunidades, de ácido desoxirribonucleico (ADN), vacunas vivas atenuadas obtenidas a partir de la inducción de mutaciones en genes que codifican para importantes factores de virulencia de M. tuberculosis hasta micobacterias no tuberculosas (MNT). Dentro de todas estas alternativas, las vacunas vivas constituyen el principal arsenal en la lucha contra la TB ya que permiten la expresión de toda una gama de antígenos micobacterianos y con ello la estimulación de diferentes poblaciones de células T (2).
Estudios experimentales in vivo demuestran la protección que ofrece la vacunación con Mycobacterium ´habana´ en modelos murinos luego del reto con M. tuberculosis H37Rv, informando valores de supervivencia 20% superiores al compararlos con los que ofrece la vacunación con BCG, bajo las mismas condiciones experimentales (3, 4). Sin embargo, se debe tener en cuenta que para el desarrollo y aprobación de cualquier candidato vacunal vivo es necesario conocer su comportamiento durante la infección in vitro de sus principales dianas celulares. El presente trabajo evalúa el proceso de fagocitosis de M. habana al infectar cultivos celulares de macrófagos (MΦ) murinos.
MATERIALES Y MÉTODOS
Bacterias: M. ´habana´ TMC 5135 se sembró en 250 mL de medio líquido Midlebrook 7H9 (DifcoLabs, Detroit, EUA) suplementado con ácido oleico-albúmina-dextrosa-catalasa (OADC, Becton Dickinson, EUA). El cultivo se incubó a 37°C, durante 10 días en un agitador orbital (IKA labortechnik) a 300 g (4). La pureza del cultivo se comprobó a través de la tinción de Ziehl-Neelsen (5). Paralelamente se sembró una alícuota en medio Agar Sangre de Carnero al 5%, con el fin de descartar contaminaciones por bacterias de crecimiento rápido.
]]> Preparación del inóculo bacteriano: Los cultivos se centrifugaron (Sigma 3-18) a 1731 RCF por 15 min y la biomasa resultante se resuspendió en tampón salina-fosfato (PBS, de sus siglas en inglés) con Tween-80 0,05%. Los agregados celulares se dispersaron mediante 10 ciclos de vórtex (GENIE 2) de 1 min. Las suspensiones bacterianas se centrifugaron a 441 RCF (Sigma 3-18) durante 15 min y el pellet se restituyó en PBS (4). El inóculo se ajustó según el tubo 0,5 de la escala de McFarland equivalente a 1,5 x 108 células/mL. A partir del mismo se realizaron las diluciones pertinentes para obtener las concentraciones celulares que se emplearon en el proceso de infección: multiplicidades de infección (MOI) de 1 y 5 bacilos por MF (MOI 1:1 y MOI 5:1). El inóculo se verificó a través del conteo del número de unidades formadoras de colonias (UFC) en medio Midleebrook 7H10 (DifcoLabs, Detroit, EUA) enriquecido con OADC.Cultivos primarios de macrófagos peritoneales murinos: Se sacrificaron ratones Balb/c, de 8 semanas de vida y un peso aproximado de 16-20 g, empleando el método de dislocación cervical (6). Posteriormente se retiró la piel de la zona abdominal. Se inyectaron 5 mL de medio RPMI (Sigma) frío y estéril en la cavidad peritoneal y se dio un ligero masaje. Luego se extrajo el líquido depositado en la cavidad peritoneal y se pasó a tubos estériles fríos. La concentración celular se ajustó a 2 x 105 células/mL mediante conteo en cámara de Neubauer (Western Germany), con azul tripano (Sigma). Las placas de 24 pozos (Greiner bio-one), previa colocación de cubreobjetos para cultivos celulares (THERMANOXTM) en el fondo de los pocillos, se sembraron con 1 mL de la suspensión celular (7) y se incubaron durante toda la noche a 37ºC en atmósfera de CO2 al 5%.
Cultivo de la línea celular de macrófagos murinos RAW 264.7: Los cultivos celulares de la línea de MΦ murinos RAW 264.7 se obtuvieron del Laboratorio de Cultivos Celulares y Anticuerpos Monoclonales del Departamento de Sistemas Críticos del Centro de Investigación, Diagnóstico y Referencia (CIDR) del Instituto de Medicina Tropical Pedro Kourí de La Habana (IPK). Las células RAW 264.7 se mantuvieron en medio Dulbecco Low Glucosa (Sigma) suplementado con suero fetal bovino (SFB, Labome) inactivado, al 10% sin antibióticos. Se sembraron 2 x 105 células/pozo en placas de 24 pocillos, previamente dotadas de cubreobjetos para cultivos celulares. Estas se incubaron durante toda la noche a 37ºC en atmósfera de CO2 al 5% (8).
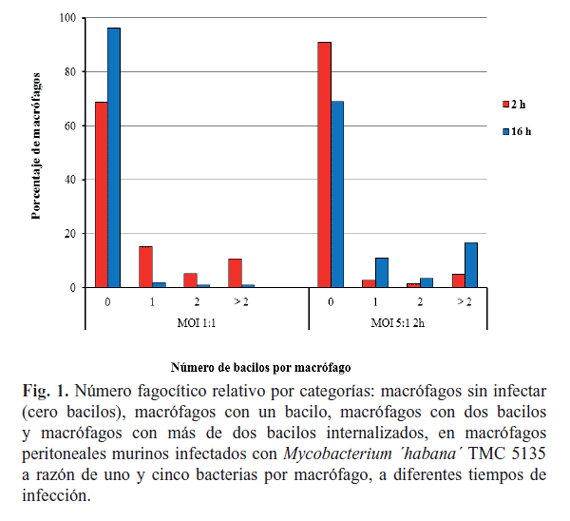
Caracterización del proceso de fagocitosis de M. ´habana´ TMC 5135 por cultivos primarios y una línea celular de macrófagos murinos: Los cultivos celulares de MΦ murinos (Línea celular RAW 264.7 y MΦ peritoneales) se inocularon, en paralelo, empleando las MOI 1:1 y 5:1. Las placas se incubaron a 37°C en 5% de CO2. Se emplearon dos tiempos de ensayo posinfección para cada MOI (2 h y 16 h). Al cabo de los tiempos seleccionados, las bacterias no fagocitadas se eliminaron a través de dos lavados con medio Dulbecco (para la línea RAW 264.7) o RPMI suplementado con SFB 10% (para los MΦ peritoneales). Las células infectadas se incubaron por 24 h más, en iguales condiciones (7). Transcurrido este tiempo las células se lavaron tres veces con el medio de cultivo correspondiente para remover las bacterias extracelulares. Los cubreobjetos con las células adheridas se fijaron con formalina al 10% durante 1 h y se tiñeron empleando la tinción de Ziehl Neelsen modificada para cultivos celulares (9). Se determinó el porcentaje de fagocitosis (PF) como la media del número de MΦ con bacterias ingeridas entre el total de MΦ por 100% (7). Paralelamente se calculó el número fagocítico (NF) para lo cual se dividieron las células observadas en cuatro categorías atendiendo a la presencia y número de bacilos en su interior: MΦ sin bacilos fagocitados, MΦ con un bacilo, MΦ con dos bacilos y MΦ con más de dos bacilos internalizados. El NF se representó como el porcentaje de MΦ en cada categoría.
Análisis estadístico: Se empleó el programa NCSS, versión 10.0.9 (Number Cruncher Statistical Systems) para el análisis estadístico de los resultados. Los experimentos realizados contaron con tres réplicas por lo que se determinó la media de los valores obtenidos y la desviación estándar correspondiente. La significación estadística de los resultados se calculó a través de la prueba t de Student de comparación de medias para muestras independientes. En todos los casos se consideraron diferencias significativas para valores de p<0,05.
Consideraciones éticas de la investigación: En el trabajo con animales de laboratorio se siguió el Principio de las 3 Rs (Reemplazo, Reducción y Refinamiento) (10). El protocolo experimental estuvo en concordancia con las regulaciones internacionales para el manejo y cuidado de los animales de experimentación (11) y las buenas prácticas de Laboratorio para estudios preclínicos (12). El protocolo contó con la aprobación del Comité de Ética de la Investigación del IPK y el Comité Institucional para el Cuidado y Uso de los Animales de Laboratorio (código CEI-IPK 46-17).
RESULTADOS Y DISCUSIÓN
Los términos índice, porcentaje y número fagocítico se emplean en la literatura internacional indistintamente: (i) para designar la relación entre las células con evidencias de fagocitosis con respecto al total o a un número previamente establecido (ej. 300 células contadas); (ii) para agrupar las células fagocíticas según el número de bacilos internalizados y (iii) para determinar la media de micobacterias por célula fagocítica (7). En el presente estudio se calculó el PF, así como el NF para determinar la eficiencia de la cepa M. ´habana´ TMC 5135 al infectar MΦ murinos. La Tabla 1 muestra los PF tras infectar MΦ peritoneales, empleando diferentes MOI y tiempos de fagocitosis. En el caso de los MΦ peritoneales, para la MOI 1:1 se observó una disminución del PF (de 29,5 a 4,0%) al aumentar el tiempo de contacto con la micobacteria. Resultados opuestos se observaron al aplicar una MOI mayor (5:1).
Los resultados obtenidos en el presente experimento pudieran aparentar ser contradictorios si tenemos en cuenta que para la MOI 1:1 obtuvimos un mayor PF a las 2 h de exposición que a las 16 h. Sin embargo, atendiendo a que la carga bacilar infectante fue baja y el tiempo de exposición el mínimo indispensable para que se produzca el proceso de fagocitosis, podríamos estar observando a las 2 h prácticamente la totalidad de bacilos que fueron internalizados. En cambio, cuando el tiempo de fagocitosis se hace mayor, una parte de los bacilos que fueron internalizados se encuentran sometidos a los mecanismos efectores de los MΦ para lograr su eliminación, lo que puede repercutir en el bajo número de bacilos observados. Estos hallazgos se corresponden con la visualización, en las láminas correspondientes a este tiempo, de bacilos fragmentados como prueba de la acción bactericida de los MΦ.
]]> El aumento del porcentaje de MΦ sin bacilos internalizados para las 16 h, acompañado por una reducción en los pertenecientes a las restantes categorías (con uno, dos y más de dos bacilos) corroboran el planteamiento anterior (Fig. 1). El análisis de la infección con una MOI 5:1 reveló el efecto contrario al descrito anteriormente (Tabla 1). En nuestra opinión, esta observación se debe a que la carga bacilar infectante fue cinco veces mayor con respecto a la MOI 1:1. Los MΦ reconocen patrones moleculares presentes en los microorganismos a través de sus receptores de membrana. La presencia de un mayor número de bacterias a ser reconocidas por estos receptores pudiera ser la causa de impedimentos estéricos que retarden el proceso de ingestión de las bacterias por los MΦ, observándose a las 2 h solo un 9% de células con bacilos internalizados. Este valor aumenta con el aumento del tiempo de exposición, indicando la ingestión de un mayor número de bacilos, independientemente de que para las 16 h ya deben estar activados los mecanismos microbicidas de los fagocitos infectados. Estos resultados se confirman al observar un menor porcentaje de MΦ sin infectar a las 16 h (Fig. 1), aparejado de un aumento en el NF de aquellas células con uno, dos y más de dos micobacterias internalizadas.Para esta última categoría se observó una duplicación del NF lo cual estuvo influido por el enfrentamiento de los MΦ a una carga bacteriana mayor. Como se muestra en la Tabla 2, la línea celular de MΦ murinos RAW 264.7 infectada con una MOI 1:1 mostró similar PF al compararla con MΦ peritoneales, en iguales condiciones de infección (29% vs. 29,5%); sin embargo, en la primera se observó un mayor NF para aquellos MΦ con más de dos bacilos internalizados, lo que pudiera constituir un marcador de una mayor eficiencia en el proceso de reconocimiento e ingestión (Fig. 2). La ingestión es una fase que distingue al proceso de digestión fagocítica. La unión de la partícula a la membrana plasmática de los MΦ es necesaria pero no suficiente para su englobamiento, requiriéndose cambios importantes en el citoesqueleto celular para que se lleve a cabo la internalización de los microorganismos (13).
Estos cambios dependen de la naturaleza de la célula infectada y a pesar de que no existen referencias que avalen este planteamiento, en este mecanismo pudiera radicar el comportamiento diferente entre los MΦ peritoneales y los pertenecientes a la línea RAW 264.7. Al aumentar la carga bacilar (MOI 5:1) se observó un 68% de células infectadas, luego de 2 h de exposición al inóculo bacteriano, lo cual difirió significativamente de lo observado para este mismo tiempo en los MΦ peritoneales. De igual forma, al aumentar el tiempo de exposición se observaron resultados significativamente diferentes. Welin y Lerm atribuyen la obtención de resultados contradictorios en este sentido, al efecto de múltiples factores (14). La fuente de los cultivos primarios o de las líneas celulares, la MOI empleada y el tiempo de infección pueden influenciar decisivamente en el resultado de la infección experimental (14). En opinión de varios expertos, el estado de activación de los MΦ constituye un factor trascendental en la determinación de cómo el hospedero puede controlar el crecimiento micobacteriano (15).
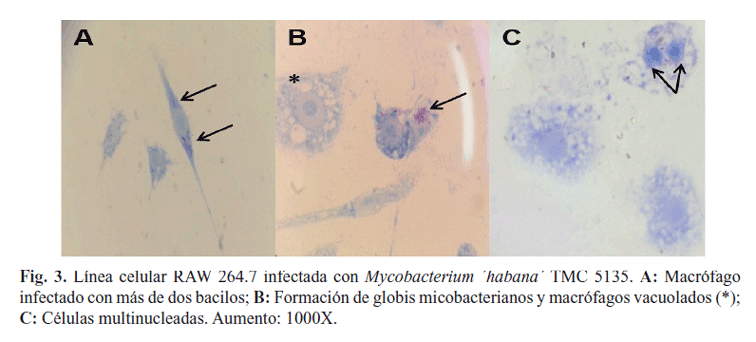
Teniendo en cuenta estas evidencias, las diferencias entre los protocolos de aislamiento/diferenciación y la variación en la robustez de las líneas celulares, resulta en una permisibilidad diferencial a la infección. En este caso pudimos observar NF que no variaron de manera significativa en las diferentes categorías en correspondencia con el tiempo de infección (Fig. 2). Adicionalmente, los MΦ pertenecientes a la línea celular RAW 264.7 mostraron una morfología diferente a la observada en los MΦ peritoneales. Se pudo constatar la presencia de MΦ vacuolados (Fig. 3B*) y células multinucleadas (Fig. 3C). Llama la atención que la mayoría de las células infectadas mostraron más de dos bacilos (Fig. 3A-C) y en algunas de ellas estos bacilos se encontraban formando globis (Fig. 3B). La observación de MΦ vacuolados en el presente experimento constituye una muestra de la activación de los mecanismos microbicidas innatos de este tipo celular ante la presencia de M. ´habana´. Estos juegan un papel crucial en la eliminación de las micobacterias. Estas células también se describen en el curso de la infección tuberculosa y se ha demostrado que en el proceso de formación de los granulomas pulmonares en individuos infectados, los MΦ se diferencian a células epitelioides, mostrándose altamente vacuolados o incluso fusionándose para formar células gigantes multinucleadas (16).
De igual forma, se pudo demostrar la presencia de células multinucleadas en el curso del experimento al emplear la línea RAW 264.7. Estudios previos demuestran que estas no solo desempeñan una función inmunorregulatoria sino que también depolimerizan las estructuras protéicas xenogénicas ingeridas. Algunos estudios han demostrado que la infección de MΦ multinucleados con micobacterias va seguida por la inducción de apoptosis, a través de la activación de las cascadas de caspasas, catepsinas y serina proteasas con propiedades antimicrobianas (16).
Algunos autores postulan que la apoptosis de las células multinucleadas infectadas es un componente de los mecanismos de defensa del sistema inmune para delimitar el crecimiento bacteriano, pero la misma se asocia también con un alto riesgo de cambios destructores en los órganos dañados (17). Orme en 2014 postuló la hipótesis del impacto de la agregación celular de M. tuberculosis en la interacción patógeno-hospedero (18). Por su parte Mahamed y colaboradores demostraron que los agregados intracelulares de M. tuberculosis no se eliminan tras la muerte de los macrófagos infectados y crecen rápidamente en el interior de la célula muerta (19). En nuestro estudio, la observación de agregados micobacterianos (globis) en el interior de los MΦ infectados con M. ´habana´ TMC 5135 permite inferir el destino de estos y la posible repercusión en la dinámica de la respuesta inmune a la infección. Entre los criterios generales que permiten establecer la calidad de un nuevo producto vacunal se encuentran los datos que se obtienen a partir del uso de sistemas in vitro de cultivos celulares (20). En el largo camino que implica la demostración del potencial vacunal de M. ´habana´, la presente investigación demostró que el proceso de fagocitosis de Mycobacterium ´habana´ TMC 5135 está influenciado por la fuente celular empleada como célula hospedadora, así como por la carga bacteriana infectante y el tiempo de exposición a la misma. Dichos aspectos contribuyen a la caracterización de la infección por esta micobacteria en sus principales células blanco de la inmunidad innata y traza el camino de futuras investigaciones preclínicas para evaluar la activación de mecanismos efectores de la inmunidad innata frente a este candidato.
]]> REFERENCIAS
1. Liu J, Tran V, Leung AS, Zhu B. BCG vaccines: Their mechanisms of attenuation and impact on safety and protective efficacy. Human Vaccines 2009;5(2):70-8.
2. van Helden PD, Hoal EG. A new TB vaccine: Fact or fiction?. Comp Immunol Microbiol Infect Dis. 2013;36(3):287-94.
3. Valdés I, Echemendía M, Mederos L, Valdivia JA, Montoro E. Aspectos relevantes del uso de Mycobacterium ´habana´ como candidato vacunal contra la tuberculosis. VacciMonitor 2011;20(3):34-9.
4. Valdés I, Montoro E, Mata-Espinoza D, Asín O, Barrios-Payán J, Francisco A, et al. Virulence, immunogenicity and protection conferred by Mycobacterium ´habana´ in a murine model of pulmonary tuberculosis. Tuberculosis 2014;94(1):65-72.
5. Organización Panamericana de la Salud. Manual para el diagnóstico bacteriológico de la tuberculosis. Normas y guía técnica. Parte I. Baciloscopía. Washington D.C: OPS; 2008.
6. American Veterinary Medical Association. Guidelines for the Euthanasia of Animals. Meacham Road Schaumburg, IL, USA: AVMA; 2013.
7. Rajavelu P, Sulochana D. A Correlation between Phagocytosis and Apoptosis in THP-1 Cells Infected with Prevalent Strains of Mycobacterium tuberculosis. Microbiol Immunol 2007;51(2):201-10.
8. Pan F, Zhao Y, Zhu S, Sun C, Lei L, Feng X, et al. Different Transcriptional Profiles of RAW264.7 Infected with Mycobacterium tuberculosis H37Rv and BCG Identified via Deep Sequencing. PLoS ONE 2012;7(12): e51988. https://doi.org/10.1371/journal.pone.0051988.
9. Diapath S.p.A. Acid Fast Bacteria acc. Ziehl-Nelseen. Savoldini, 71-24057 Martinengo (BG), Italy: Diapath; 2013.
10. Russell WMS, Burch R. The principles of humane experimental technique. London: Methuen & Co. Ltd.; 1959.
]]>11. National Research Council. Guide for the care and use of laboratory animals. Washington DC: National Academies Press; 2011.
12. World Health Organization. Good practices for pharmaceutical microbiology laboratories, Geneva: WHO; 2013.
13. Gordon S. Phagocytosis: An Immunobiologic Process. Immunity 2016; 44:463-75.
14. Welin A, Lerm M. Inside or outside the phagosome? The controversy of the intracellular localization of Mycobacterium tuberculosis. Tuberculosis 2012;92(2):113-20.
15. Duque MA, Rojas M. Activación alternativa del macrófago: La diversidad en las respuestas de una célula de la inmunidad innata ante la complejidad de los eventos de su ambiente. Inmunología 2007;26(2):73-86.
]]>16. Santucci P, Bouzid F, Smichi N, Poncin I, Kremer L, De Chastellier C, et al. Experimental Models of Foamy Macrophages and Approaches for Dissecting the Mechanisms of Lipid Accumulation and Consumption during Dormancy and Reactivation of Tuberculosis. Front Cell Infect Microbiol 2016;6:122. doi: 10.3389/fcimb.2016.00122.
17. Ilin DA, Arkhipov S A, Shkurupy V. In Vitro Study of Cytophysiological Characteristics of Multinuclear Macrophages from Intact and BCG-Infected Mice. Bulletin of Experimental Biology and Medicine. Microbiology and Immunology 2016;160(5):668-71.
18. Orme IM. A new unifying theory of the pathogenesis of tuberculosis. Tuberculosis 2014;94:8-14.
19. Mahamed D, Boulle M, Ganga Y, Mc Arthur CH, Skroch S, Oom S, et al. Intracellular growth of Mycobacterium tuberculosis after macrophage cell death leads to serial killing of host cells. eLife 2017;6:e22028. doi: 10.7554/eLife.22028.
20. Marinova DJ, Gonzalo-Asensio N, Aguilo, Martín C. Recent developments in tuberculosis vaccines. Expert Rev Vaccines 2013;12(12):1431-8.
]]>Recibido: Diciembre de 2017 Aceptado: Febrero de 2018
]]>
{kind=link}
{kind=link}
{kind=link}